История изучения
Биологическая роль некоторых реакций цикла Кребса (ЦК) была изучена американским биохимиком венгерского происхождения Альбертом Сент-Дьердьи. В частности, он выделил ключевой компонент ЦТК — фумарат. Исследования в этом направлении продолжил Ганс Кребс. В итоге он установил всю последовательность реакций и соединений, образующиеся на всех этапах процесса. Ученый не смог определить, с преобразования какой кислоты начинается цикл — лимонной или изолимонной. Сейчас известно, что это лимонная кислота. Поэтому ЦК называют также цитратным или циклом лимонной кислоты.
Позднее американец Альберт Ленинджер, занимающийся биоэнергетикой, определил, что все реакции ЦК протекают в митохондриях клеток. С получением доступа к изотопам углерода появилась возможность более досконального изучения и уточнения данных о промежуточных соединениях на разных этапах цикла.
С пищей в организм поступают три основные группы сложных биохимических соединений — белки, жиры и углеводы. Они являются первичными метаболитами, потому что участвуют в обмене веществ или в метаболизме. Этот процесс происходит между любыми живыми клетками и окружающей средой непрерывно. Суть цикла Кребса заключается в том, что он является областью схождения двух путей метаболизма. Это следующие процессы:
- катаболизм, при котором происходит распад более сложных веществ на простые, в частности, глюкозы на моносахариды;
- анаболизм — синтез сложных веществ из простых, например, белков из аминокислот.
После попадания в пищеварительную систему сложные вещества расщепляются под действием ферментов на более простые, которые внутри клеток превращаются сначала в пируват (пировиноградную кислоту), а затем — в ацетильный остаток. Все эти преобразования можно назвать подготовкой к ЦК, а образование остатка — его запуском или начальным этапом.
Дальнейшие стадии цикла трикарбоновых кислот являются частью катаболизма. Процесс идет каскадно. Каждый предыдущий этап запускает последующий, а промежуточные продукты химических реакций служат не только для продолжения цикла, но и при определенных потребностях организма могут пополнять запасы веществ, необходимых для синтеза новых соединений (анаболизма).
Клеточное дыхание
Для нормальной жизнедеятельности живым клеткам постоянно требуется энергия. Ее главный универсальный источник — аденозинтрифосфат (АТФ), способный встраиваться в белки организма напрямую. Это соединение получается в результате ряда реакций окисления, носящих общее название «клеточное дыхание». При этом происходит постепенный распад органических веществ вплоть до простейших неорганических — углекислого газа CO2 и воды H2O.
Структурное строение молекул АТФ содержит фосфорангидридные связи, которые имеют свойство накапливать высвобожденную при прохождении реакций клеточного дыхания энергию, поэтому называются макроэргическими. Так создаются энергетические запасы клеток, которые могут высвобождается при необходимости разрывом этих связей. Процесс синтеза АТФ и класса вспомогательных соединений включает три этапа:
- Гликолиз происходит в цитоплазме.
- В матриксе митохондрий проходят все химические реакции цикла Кребса.
- Окислительное фосфорилирование на внутренней мембране митохондрий.
Преобразование аденозиндифосфата (АДФ) в АТФ характерно для всех этапов. Но наибольшее суммарное количество молекул с макроэргическими связями образуется при фосфорилировании. Это не значит, что процессы гликолиза и ЦК менее важны. Многие соединения, образующиеся во время их протекания, участвуют в регуляции клеточного дыхания.
Описание процесса
Протекание ЦК достаточно экономно с точки зрения энергозатрат. Такой эффект достигается благодаря тому, что он связывает два метаболических направления. В процесс вовлекаются вещества, подлежащие утилизации, которые либо служат энергетическим «топливом», либо возвращаются в круг анаболизма. Подготовительная стадия ЦК заключается в распаде глюкозы, аминокислот и жирных кислот на молекулы пирувата или лактата.
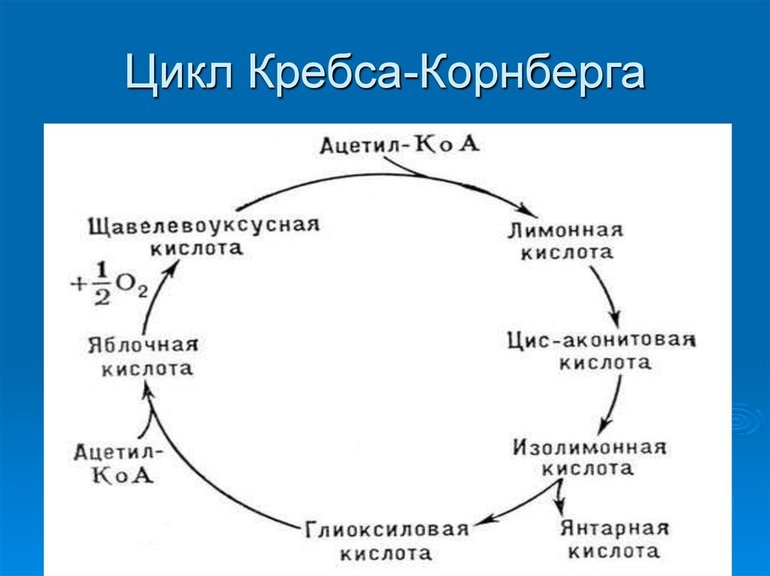
Органеллы митохондрий способны преобразовывать пируват в ацетильный остаток (ацетил-коэнзим А или ацетил-КоА), представляющий собой вместе с тиольной группой, которая может его переносить, кофермент А. Некоторое соединения могут сразу распадаться до ацетил-КоА, минуя стадию пирувата. При этом пировиноградная кислота может вовлекаться непосредственно в ЦК, не преобразуясь в ацетил-КоА.
Начальные этапы
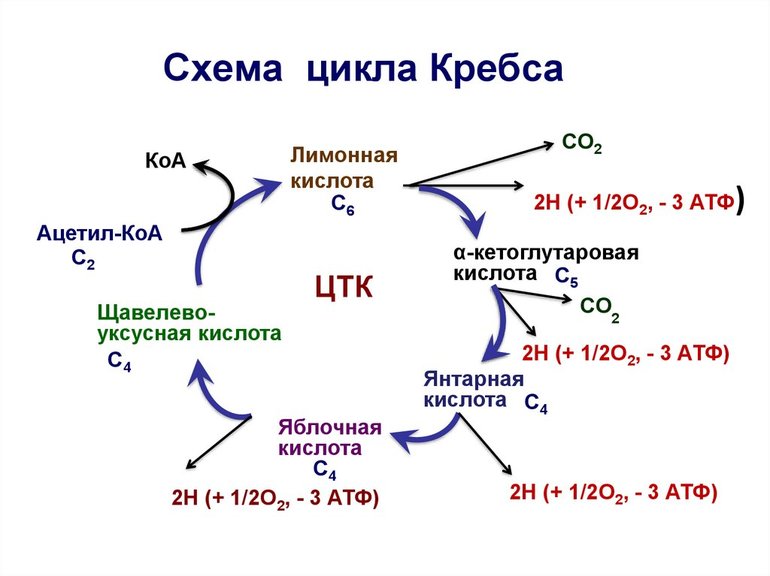
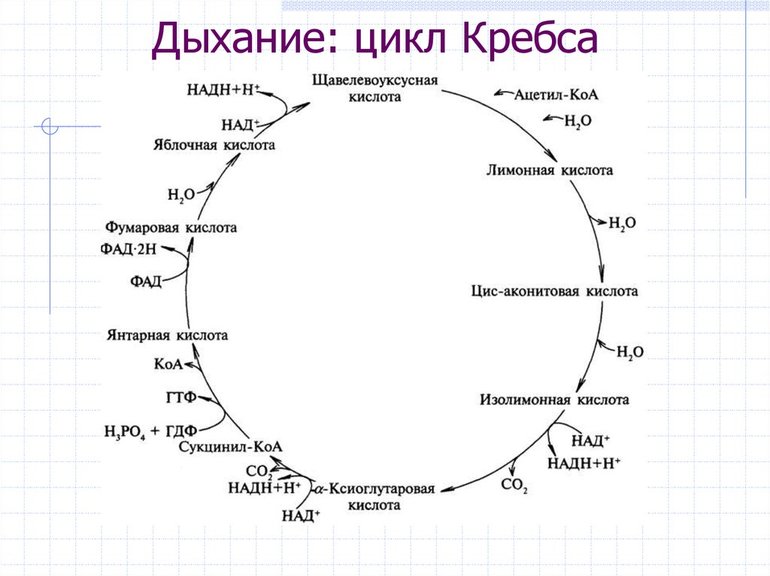
Первая стадия необратима и состоит из конденсации ацетил-КоА с четырехуглеродным веществом — оксалоацетатом (щавелевоуксусной кислотой или ЩУК), что приводит к образованию шестиуглеродного цитрата (лимонной кислоты). Во время реакции метильная группа ацетил-КоА соединяется с карбонильной группой ЩУК. Благодаря быстрому гидролизу промежуточного соединения цитроил-КоА этот этап проходит без затрат энергии извне.
На второй стадии образуется изоцитрат (изолимонная кислота) из цитрата через цис-аконитат. Это реакция обратимой изомеризации через образование промежуточной трикарбоновой кислоты, в которой катализатором выступает фермент аконитатгидратаза.
Далее происходит дегидрирование и декарбоксилирование изоцитрата до промежуточного соединения оксалосукцинат с выделением углекислого газа. После декарбоксилирования оксалосукцината образуется енольное соединение, которое перестраивается и превращается в пятиуглеродную кислоту — α-кетоглутарат (оксоглутарата), чем и завершает третью ступень ЦК. Четвертый этап — α-кетоглутарат декарбоксилирует и реагирует с ацетил-КоА. При этом получается сукцинил-КоА, соединение янтарной кислоты и коэнзима-А, выделяется СО2.
Замыкание цикла
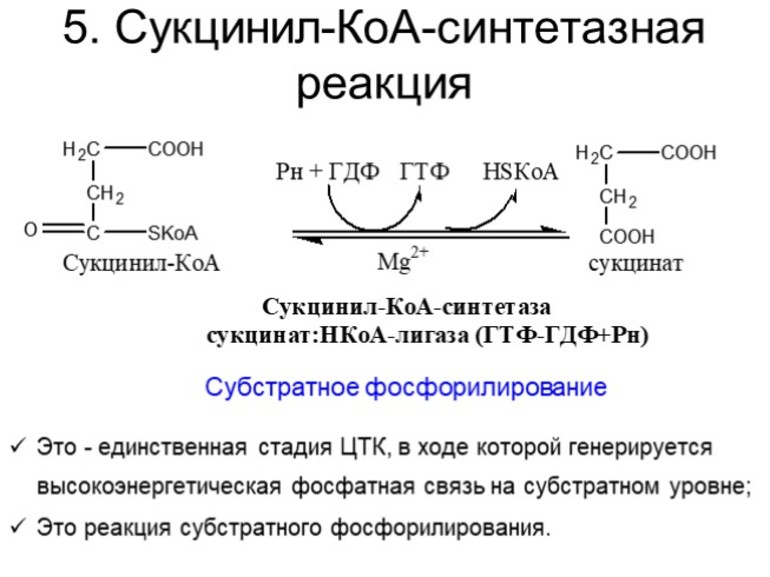
На пятой стадии сукцинил-КоА преобразуется в сукцинат (янтарную кислоту). Для этого этапа характерно субстратное фосфолирование, подобное синтезу АТФ при гликолизе. Введение в ЦК фосфорной группы РО3 становится возможным благодаря присутствию фермента ГДФ (гуанозиндифосфата) или АДФ (аденозиндифосфата), которые в процессе синтеза сукцината из дифосфатов становятся трифосфатами.
Начиная с шестой стадии, цикл начинает постепенно замыкаться. Сначала сукцинат под действием каталитического фермента сукцинатдегидрогеназы дегидрирует до фумарата. Дальнейшее дигидрирование приводит к седьмому этапу — образованию L-малата (яблочной кислоты) из фуратата через переходное соединение с карбанионом.
Последняя реакция цикла трикарбоновых кислот малат окисляется до щавелевоуксусной кислоты. Первая стадия следующего ЦК начинается с новой молекулы ацетил-КоА.
Значение и функции
Этот восьмиэтапный циклический процесс, итогом которого является окисление ацетильного остатка до углекислого газа, может показаться излишне сложным. Тем не менее, он имеет огромное значение в метаболизме промежуточных реакций и выполняет ряд функций. К ним относятся:
- энергетическая;
- анаболическая;
- катаболическая;
- транспортная.
Цикл Кребса участвуют в катаболизме жиров и углеводов. Соединения, образующиеся на разных стадиях процесса, участвуют в синтезе многих необходимых для организма веществ — глутамина, порфиринов, глицина, фенилаланина, цистеина и других. Когда промежуточные продукты покидают ЦК для участия в синтезе, происходит их замещение с помощью так называемых анаплеротических реакций, которые катализируются регуляторными ферментами, например, пируваткарбоксилазой.
Транспортная функция ЦК заключается в содействии гликолизу. Глюкозу невозможно превратить сразу в АТФ, поэтому механизм гликолиза действует поэтапно и сопровождается постоянным перемещением атомов и катионов водорода от одних соединений к другим. Для их транспортировки нужны специальные соединения, которые получаются на одной из стадий ЦТК. Участвующие в гликолизе коферменты цикла Кребса:
- НАД*H+(Никотинамидадениндинуклеотид с катионом водорода). Образуется на III стадии ЦК.
- ФАД*H2 (Флавинадениндинуклеотид с молекулой водорода). Появляется на V стадии ЦК.
Реакции ЦК имеют и большое клиническое значение. Хотя для людей не свойственны мутации, связанные с генами ферментов, участвующих в цикле, однако их редкие проявления губительны для здоровья. Они могут приводить к опухолям мышц и почек, нарушениям работы нервной системы.
Существует множество видов визуального и слухового отображения цикла Кребса — схемы с формулами, уравнения химических реакций, разнообразные таблицы и даже мнемонические способы для полного запоминания его главных «участников».
Читайте также:
- Эксперимент Милгрэма
- Плазмолиз и деплазмолиз
1. Матрикс митохондрии
содержит все ферменты цикла Кребса. В митохондриях ПВК превращается в ацетил-кофермент А (за счет присоединения КоА). В КоА входит адениловый нуклеотид, а также пантотеновая кислота (синтезируется в кишечнике человека бактериями). Надо помнить, что окисляться могут и жирные кислоты, и аминокислоты — но в результате всегда создается именно ацетил-КоА. При этом идет восстановление HAДН из НАД+ и выделяется СО2.
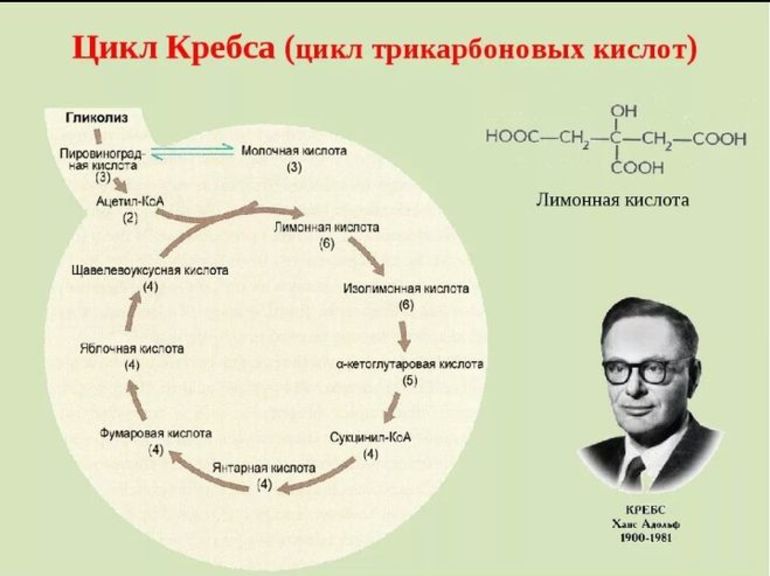
2. Ацетил-КоА объединяется с молекулой щавелеускусной кислоты, происходит образование лимонной кислоты.
3. Дальше лимонная кислота продолжает окисляться в процессе ферментных реакций.
1) Восстанавливаются 3 молекулы НАДН, одна молекула ФАДН2, и ГТФ (гуанозинтрифосфат).
2) ГТФ используется для фосфорилирования АДФ и образования АТФ.
3) Лимонная кислота утрачивает 2 углеродных атома, в результате чего возникают две молекулы СО2.
4. Пройдя одну за другой семь реакций, лимонная кислота вновь превращается в щавелеуксусную, а та, в свою очередь, вновь соединяется с ацетил-КоА. Цикл замыкается.
1) В лимонной кислоте словно бы сгорает присоединившийся ранее остаток ацетил-КоА.
2) Протоны водорода и электроны переносятся на акцепторы — НАД+ и ФАД.
3) В итоге энергия органических соединений аккумулируется в молекулах НАДН, ФАДН2, АТФ.
4) К тому же, подчеркнем, в цикле Кребса идет выделение CO2.
Цепь переноса электронов. Окислительное фосфорилирование
1. Электроны от НАДН и ФАДН2 переносятся по цепи переноса электронов (которая имеет много звеньев) к конечному акцептору — кислороду. В ходе этого процесса освобождается энергия электронов НАДН и ФАДН2, которая идет на синтез АТФ из АДФ. Именно потому данный процесс и назван фосфорилированием. При этом электроны концентрируются с внутренней стороны крист, а протоны с внешней (противоположно тому, как это было в хлоропластах).
2. Перенос электронов идет по цепи, расположенной с внутренней стороны внутренней мембраны митохондрий, где находятся ряд переносчиков. Самый сильный акцептор электронов — в конце цепи — кислород. Промежуточные переносчики: убихинон, ФМН, НАДН-дегидрогеназа, цитохромы b, с1, с, цитохромоксидаза — комплекс цитохромов а и а3 (в них есть атомы меди).
3. Куда идут атомы водорода и электроны? Атомы водорода, электроны от НАДН и ФАДН2 отправляются на внутреннюю сторону мембраны митохондрии. Протоны идут в межмембранное пространство, лежащее между двумя мембранами митохондрий, наружной и внутренней, формируя протонный резервуар. А электроны атома водорода остаются на внутренней стороне мембраны, где они и концентрируются. Так создается разность потенциалов.
4. Протоны двигаются через канал в молекуле фермента АТФ-синтетазы
при достижении разности потенциалов определенной величины. АТФ-синтетазы встроены во внутренние мембраны митохондрий. Они образуют АТФ из АДФ. Энергия протонов при движении их через канал фермента расходуется на синтез АТФ. По мере образования АТФ протонный резервуар теряет свою энергию.
5. Внутри митохондрий катионы водорода Н+, соединяясь с кислородом и электронами, образуют воду: 2Н+ + 0,5О2 = Н2О.
6. Подведем итог. В цикле трикарбоновых кислот образуются НАДН, ФАДН2, СО2. При окислительном фосфорилировании образуются 34 АТФ, 6Н2О. В результате окисление одной молекулы глюкозы дает 38 АТФ, 6 СО2, 6Н2О.
1) КПД кислородного этапа — 55 процентов.
2) 45 процентов энергии теряется в виде тепла.
3) АТФ идет на химическую работу (биосинтез), механическую (мышцы), осмотическую (накопление и вывод веществ), электрическую (нервная ткань).
Представьте себе, что все белки, жиры и углеводы, которые мы получаем с приемом пищи, распадаются на маленькие пазлы, которые собираются вновь только в одном определенном участке нашего организма — митохондриях. Там же происходит сортировка и параллельное протекание реакций созидания и разрушения. Разберемся подробнее:
Что такое цикл Кребса
Цикл Кребса — это цепочка химических реакций, происходящих в митохондриях каждой клетки нашего тела, которая называется циклом потому, что продолжается непрерывно. Она же является и общим конечным путем окисления ацетильных групп (в виде ацетил-КоА), в которые превращается в процессе распада большая часть органических молекул, играющих роль «клеточного топлива» или «субстратов окисления»: углеводов, жирных кислот и аминокислот.
Функции цикла Кребса:
- анаболическая (синтез новых органических веществ);
- энергетическая (питание организма)
- катаболическая (превращение некоторых веществ в катализаторы)
- транспортная (транспортировка водорода, участвующего в дыхании клеток).
Атомы водорода, высвобождающиеся в окислительно-восстановительных реакциях, доставляются в цепь переноса электронов при участии НАД- и ФАД- зависимых дегидрогеназ, в результате чего происходит образование 12 высокоэнергетических фосфатных связей: синтез 12 молекул аденозинтрифосфорной кислоты (АТФ) из аденозиндифосфорной кислоты (АДФ).
Как работает цикл Кребса?
В организме аминокислоты, жирные кислоты и пируват образуют ацетил-КоА.
Когда ацетил-КоА попадает в митохондриальный матрикс, он связывается с молекулой оксалацетата и превращается в лимонную кислоту (цитрат). Цитрат, в свою очередь, под действием фермента аконитазы превращается в цис-аконитат, оставляя молекулу воды.
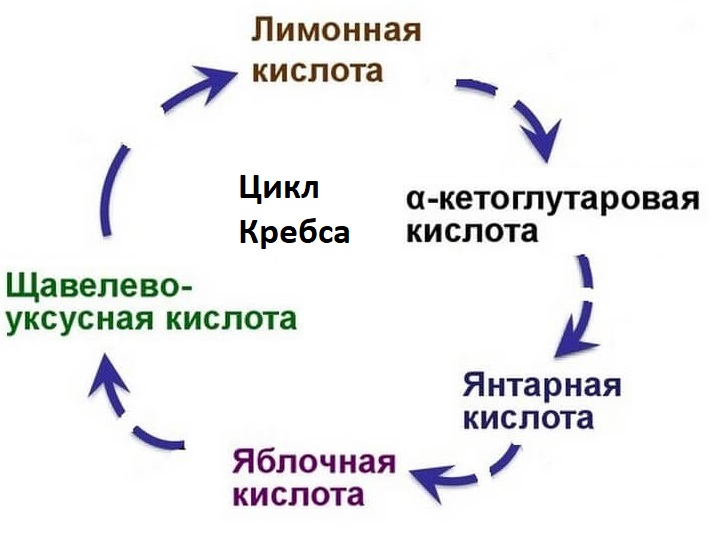
В свою очередь цис-аконитат превращается в изоцитрат под действием фермента изоцитратдегидрогеназы. Изоцитрат превращается в альфа-кетоглутарат под действием изоцитратдегидрогеназы.
Альфа-кетоглутарат превращается в сукцинил-КоА альфа-кетоглутаратдегидрогеназой и добавлением ацетил-КоА. Он подвергается сукцинату под действием сукцинат-тиокиназы. Сукцинатдегидрогеназа превращает его в фумарат. Фумарат превращается в L-малат через фумаразу. L-малат под действием фермента малатдегидрогеназы восстанавливает оксалацетат, который может снова вступать в реакцию с молекулой ацетил-КоА и повторять цикл.
Результатом этого цикла является образование CO2 и водорода, а также воды. Ионы на выходе из процесса участвуют в ресинтезе АТФ, что помогает организму восстановить еще один источник энергии — трифосфат аденозина.
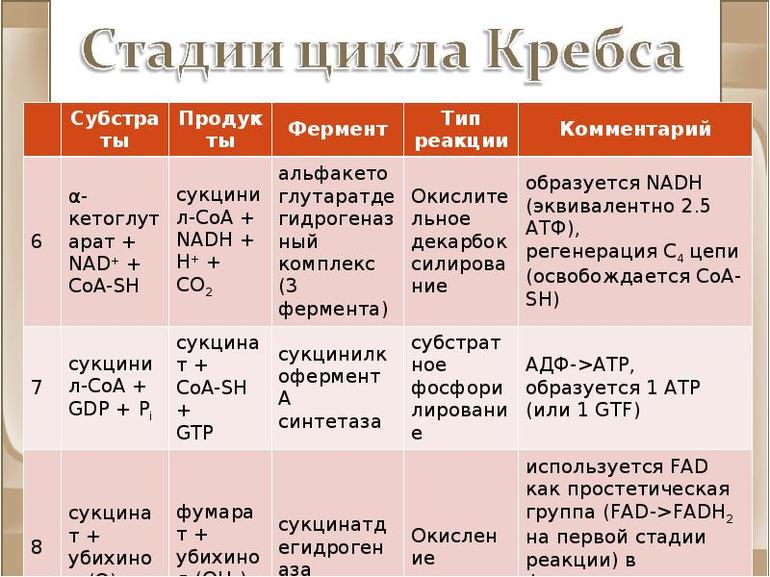
Стадии цикла Кребса
Окисление ацетильного остатка происходит в несколько стадий, образующих циклический процесс из 8 основных этапов:
|
Основные этапы цикла Кребса |
|
|
I этап |
Конденсация ацетил-КоА и оксалоацетата с образованием цитрата. |
|
Происходит реакция отщепление карбоксильной группы аминокислот, в процессе которой образуется ацетил-КоА *он выполняет функцию транспортировки углерода в различных обменных процессах. При соединении с молекулой щавелевой кислоты получается цитрат *фигурирует в буферных обменах. На данном этапе кофермент А полностью высвобождается, и получаем молекулу воды. Данная реакция необратима. |
|
|
II этап |
Превращение цитрата в изоцитрат. |
|
Дегидрирование (отщепление молекул воды) от цитрата, в результате которого получается цис-аконитат. И присоединяя молекулу воды, переходит в изолимонную кислоту (изоцитрат) |
|
|
III этап |
Превращение изоцитрата в а-кетоглутарат. |
|
Изолимонная кислота(изоцитрат) дегидрируется в присутствии НАД— зависимой изо-цитратдегидрогеназы.На выходе получаем альфа-кетоглутарат. *Альфа-кетоглутарат участвует в регуляции всасывания аминокислот, нормализует метаболизм и положительно влияет на антистрессорные процессы. Также образуется NADH (аллостерический фермент) |
|
|
IV этап |
Окисление α-кетоглутарата до сукцинил-КоА |
|
Окислительное декарбоксилирование а-кетоглутарата с образованием сукцинил-КоА — тиоэфира, содержащего высокоэнергетическую фосфатную связь. |
|
|
V этап |
Превращение сукцинил-КоА в сукцинат. |
|
Пятая реакция катализируется ферментом сукцинил-КоА-синтетазой. В ходе этой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата превращается в янтарную кислоту (сукцинат). Так же происходит образование высокоэргической фосфатной связи ГТФ за счет тиоэфирной связи сукцинил-КоА. |
|
|
VI этап |
Дегидрогенирование сукцината. Образование фумарата. |
|
Образовавшийся сукцинат превращается в фумарат под действием фермента сукцинат-дегидрогеназы. Единственная дегидрогеназная реакция цикла Кребса, в ходе которой осуществляется прямой перенос водорода с субстрата на флавопротеин без участия НАД+. |
|
|
VII этап |
Образование малата из фумарата. |
|
Под влиянием фермента фумаратгидратазы (фумаразы). Образовавшаяся при этом фумаровая кислота гидратируется, продуктом реакции является L-яблочная кислота (L-малат). |
|
|
VIII этап |
Превращение малата в оксалоацетат. |
|
Под влиянием митохондриальной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в оксалоацетат. Происходит полное «сгорание» одной молекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоянное поступление в систему ацетил-КоА. А коферменты (НАД+ и ФАД), перешедшие в восстановленное состояние, должны снова и снова окисляться. |
Реакции цикла Кребса по стадиям
Для облегчения запоминания ферментативных реакций цикла:
ЩУКа съела ацетат, получается цитрат
через цис-аконитат будет он изоцитрат
водороды отдав НАД, он теряет СО2
этому безмерно рад альфа-кетоглутарат
окисление грядёт: НАД похитит водород
В1 и липоат с коэнзимом А спешат,
отбирают СО2, а энергия едва
в сукциниле появилась сразу ГТФ родилась
и остался сукцинат. вот добрался он до ФАДа,
водороды тому надо водороды потеряв,
стал он просто фумарат. фумарат воды напился,
и в малат он превратился тут к малату НАД пришёл,
водороды приобрёл ЩУКа снова объявилась
и тихонько затаилась Караулить ацетат…
Понятие метаболизма
Метаболизм — совокупность всех химических реакций, протекающих в живом организме. Значение метаболизма состоит в создании необходимых организму веществ и обеспечении его энергией.
Выделяют две составные части метаболизма — катаболизм и анаболизм.
Составные части метаболизма
| Часть | Характеристика | Примеры | Затраты энергии |
| Катаболизм (энергетический обмен, диссимиляция) | Совокупность химических реакций, приводящих к образованию простых веществ из более сложных | Гидролиз полимеров до мономеров и расщепление последних до низкомолекулярных соединений углекислого газа, воды, аммиака и других веществ | Энергия выделяется |
| Анаболизм (пластический обмен, ассимиляция) | Совокупность химических реакций синтеза сложных веществ из более простых | Образование углеводов из углекислого газа и воды в процессе фотосинтеза, реакции матричного синтеза | Энергия поглощается |
Процессы пластического и энергетического обмена неразрывно связаны между собой. Все синтетические (анаболические) процессы нуждаются в энергии, поставляемой в ходе реакций диссимиляции. Сами же реакции расщепления (катаболизма) протекают лишь при участии ферментов, синтезируемых в процессе ассимиляции.
Роль ФТФ в метаболизме
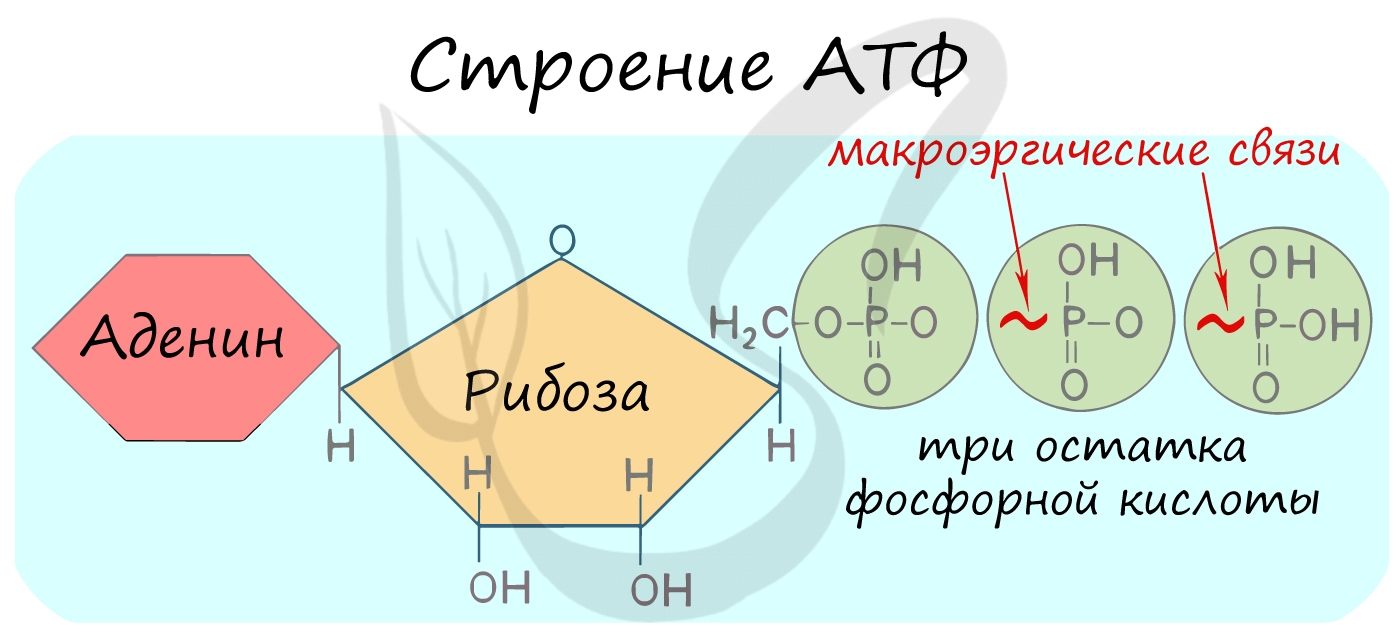
Энергия, высвобождающаяся при распаде органических веществ, не сразу используется клеткой, а запасается в форме высокоэнергетических соединений, как правило, в форме аденозинтрифосфата (АТФ). По своей химической природе АТФ относится к мононуклеотидам.
АТФ (аденозинтрифосфорная кислота) — мононуклеотид, состоящий из аденина, рибозы и трёх остатков фосфорной кислоты, соединяющихся между собой макроэргическими связями.
В этих связях запасена энергия, которая высвобождается при их разрыве:
АТФ + H2O → АДФ + H3PO4 + Q1
АДФ + H2O → АМФ + H3PO4 + Q2
АМФ + H2O → аденин + рибоза + H3PO4 + Q3,
где АТФ — аденозинтрифосфорная кислота; АДФ — аденозиндифосфорная кислота; АМФ — аденозинмонофосфорная кислота; Q1 = Q2 = 30,6 кДж; Q3 = 13,8 кДж.
Запас АТФ в клетке ограничен и пополняется благодаря процессу фосфорилирования. Фосфорилирование — присоединение остатка фосфорной кислоты к АДФ (АДФ + Ф → АТФ). Он происходит с разной интенсивностью при дыхании, брожении и фотосинтезе. АТФ обновляется чрезвычайно быстро (у человека продолжительность жизни одной молекулы АТФ менее 1 мин).
Энергия, накопленная в молекулах АТФ, используется организмом в анаболических реакциях (реакциях биосинтеза). Молекула АТФ является универсальным хранителем и переносчиком энергии для всех живых существ.
Энергетический обмен
Энергию, необходимую для жизнедеятельности, большинство организмов получают в результате процессов окисления органических веществ, то есть в результате катаболических реакций. Важнейшим соединением, выступающим в роли топлива, является глюкоза.
По отношению к свободному кислороду организмы делятся на три группы.
Классификация организмов по отношению к свободному кислороду
| Группа | Характеристика | Организмы |
| Аэробы (облигатные аэробы) | Организмы, способные жить только в кислородной среде | Животные, растения, некоторые бактерии и грибы |
| Анаэробы (облигатные анаэробы) | Организмы, неспособные жить в кислородной среде | Некоторые бактерии |
| Факультативные формы (факультативные анаэробы) | Организмы, способные жить как в присутствии кислорода, так и без него | Некоторые бактерии и грибы |
У облигатных аэробов и факультативных анаэробов в присутствии кислорода катаболизм протекает в три этапа: подготовительный, бес- кислородный и кислородный. В результате органические вещества распадаются до неорганических соединений. У облигатных анаэробов и факультативных анаэробов при недостатке кислорода катаболизм протекает в два первых этапа: подготовительный и бескислородный. В результате образуются промежуточные органические соединения, еще богатые энергией.
Этапы катаболизма
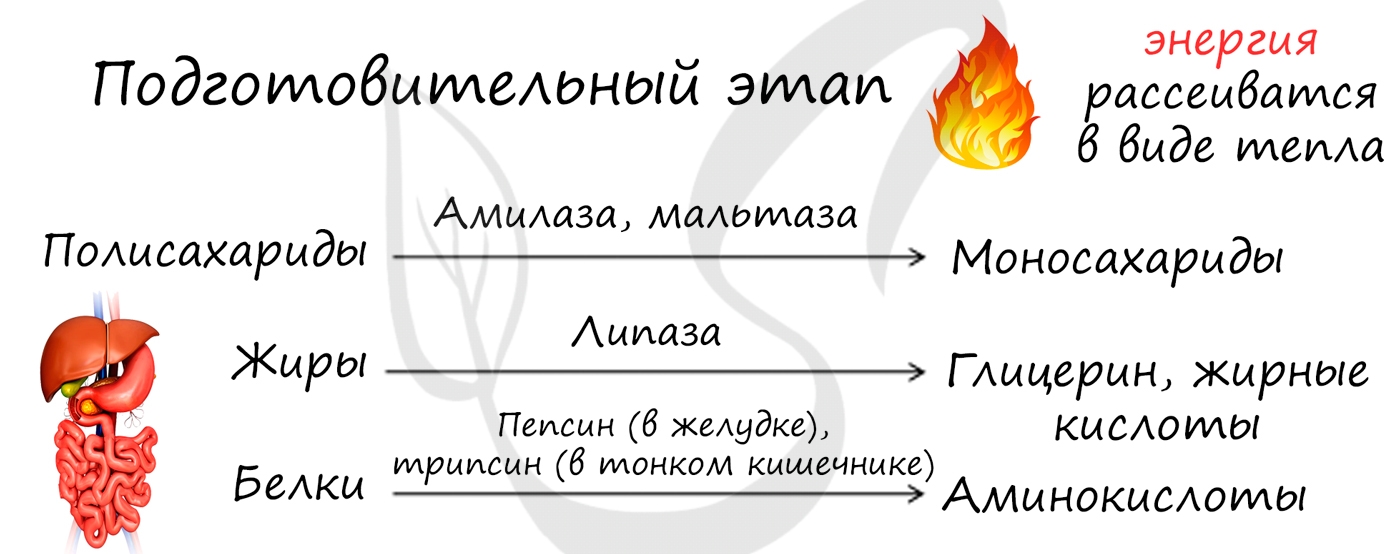
1. Первый этап — подготовительный — заключается в ферментативном расщеплении сложных органических соединений на более простые. Белки расщепляются до аминокислот, жиры — до глицерина и жирных кислот, полисахариды — до моносахаридов, нуклеиновые кислоты — до нуклеотидов. У многоклеточных организмов это происходит в желудочно-кишечном тракте, у одноклеточных — в лизосомах под действием гидролитических ферментов. Высвобождающаяся при этом энергия рассеивается в виде теплоты. Образовавшиеся органические соединения либо подвергаются дальнейшему окислению, либо используются клеткой для синтеза собственных органических соединений.
2. Второй этап — неполное окисление (бескислородный) — заключается в дальнейшем расщеплении органических веществ, осуществляется в цитоплазме клетки без участия кислорода. Главным источником энергии в клетке является глюкоза. Бескислородное, неполное окисление глюкозы называется гликолизом. В результате гликолиза одной молекулы глюкозы образуется по две молекулы пировиноградной кислоты (ПВК, пируват) CH3COCOOH, АТФ и воды, а также атомы водорода, которые связываются молекулой-переносчиком НАД+ и запасаются в виде НАД·Н.
Суммарная формула гликолиза имеет следующий вид:
C6H12O6 + 2H3PO4 + 2АДФ + 2НАД+ → 2C3Н4O3 + 2H2O + 2АТФ + 2НАД·Н.
Далее при отсутствии в среде кислорода продукты гликолиза (ПВК и НАД·Н) перерабатываются либо в этиловый спирт — спиртовое брожение (в клетках дрожжей и растений при недостатке кислорода)
CH3COCOOH → СО2 + СН3СОН
СН3СОН + 2НАД·Н → С2Н5ОН + 2НАД+,
либо в молочную кислоту — молочнокислое брожение (в клетках животных при недостатке кислорода)
CH3COCOOH + 2НАД·Н → C3Н6O3 + 2НАД+.
При наличии в среде кислорода продукты гликолиза претерпевают дальнейшее расщепление до конечных продуктов.
3. Третий этап — полное окисление (дыхание) — заключается в окислении ПВК до углекислого газа и воды, осуществляется в митохондриях при обязательном участии кислорода.
Он состоит из трёх стадий:
А) образование ацетилкоэнзима А;
Б) окисление ацетилкоэнзима А в цикле Кребса;
В) окислительное фосфорилирование в электронотранспортной цепи.
А. На первой стадии ПВК переносится из цитоплазмы в митохондрии, где взаимодействует с ферментами матрикса и образует 1) диоксид углерода, который выводится из клетки; 2) атомы водорода, которые молекулами-переносчиками доставляются к внутренней мембране митохондрии; 3) ацетилкофермент А (ацетил-КоА).
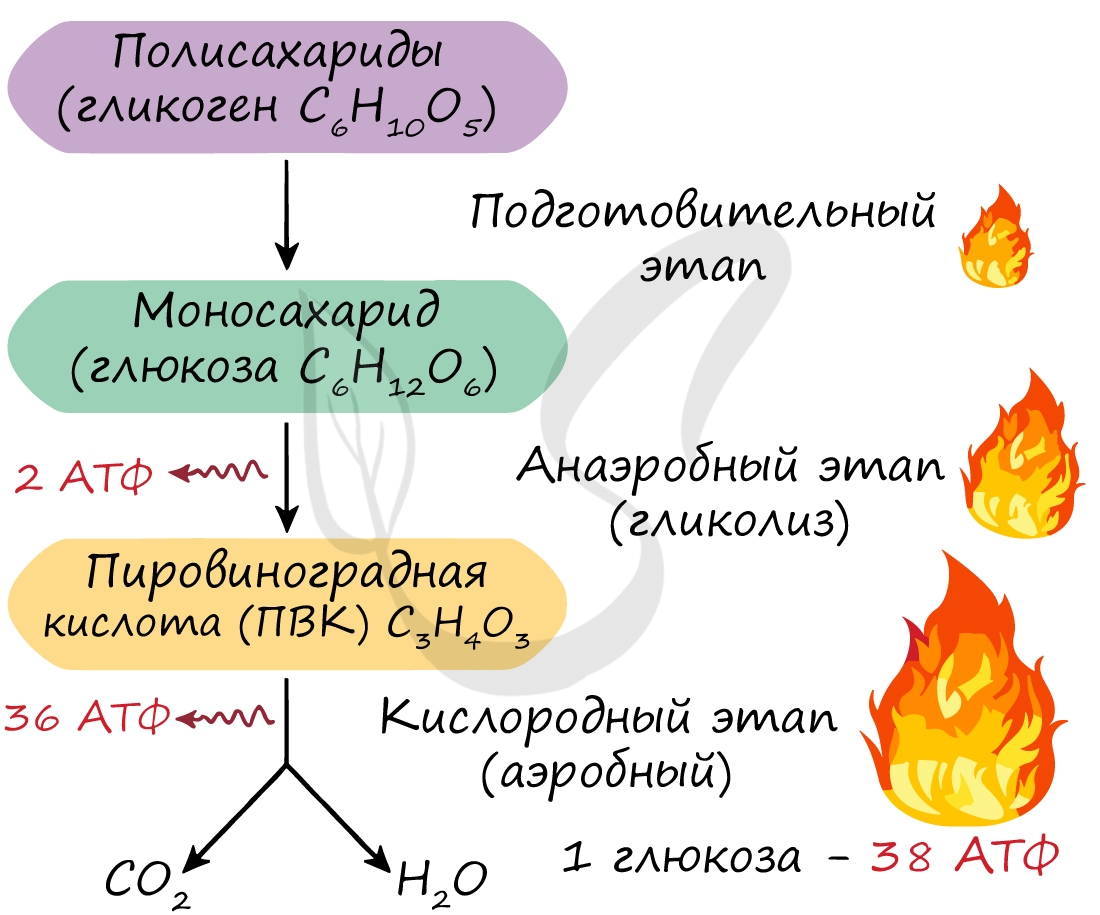
Б. На второй стадии происходит окисление ацетилкоэнзима А в цикле Кребса. Цикл Кребса (цикл трикарбоновых кислот, цикл лимонной кислоты) — это цепь последовательных реакций, в ходе которых из одной молекулы ацетил-КоА образуются 1) две молекулы диоксида углерода, 2) молекула АТФ и 3) четыре пары атомов водорода, передаваемые на молекулы-переносчики — НАД и ФАД. Таким образом, в результате гликолиза и цикла Кребса молекула глюкозы расщепляется до СО2, а высвободившаяся при этом энергия расходуется на синтез 4 АТФ и накапливается в 10 НАД·Н и 4 ФАД·Н2.
В. На третьей стадии атомы водорода с НАД·Н и ФАД·Н2 окисляются молекулярным кислородом О2 с образованием воды. Один НАД·Н способен образовывать 3 АТФ, а один ФАД·Н2–2 АТФ. Таким образом, выделяющаяся при этом энергия запасается в виде ещё 34 АТФ.
Этот процесс протекает следующим образом. Атомы водорода концентрируются около наружной стороны внутренней мембраны митохондрии. Они теряют электроны, которые по цепи молекул-переносчиков (цитохромов) электронотранспортной цепи (ЭТЦ) переносятся на внутреннюю сторону внутренней мембраны, где соединяются с молекулами кислорода:
О2 + е— → О2—.
В результате деятельности ферментов цепи переноса электронов внутренняя мембрана митохондрий изнутри заряжается отрицательно (за счёт О2—), а снаружи — положительно (за счёт Н+), так что между её поверхностями создаётся разность потенциалов. Во внутреннюю мембрану митохондрий встроены молекулы фермента АТФ- синтетазы, обладающие ионным каналом. Когда разность потенциалов на мембране достигает критического уровня, положительно заряженные частицы H+ силой электрического поля начинают проталкиваться через канал АТФазы и, оказавшись на внутренней поверхности мембраны, взаимодействуют с кислородом, образуя воду:
1/2О2— +2H+ → Н2О.
Энергия ионов водорода H+, транспортирующихся через ионный канал внутренней мембраны митохондрии, используется для фосфорилирования АДФ в АТФ:
АДФ + Ф → АТФ.
Такое образование АТФ в митохондриях при участии кислорода называется окислительным фосфорилированием.
Суммарное уравнение расщепления глюкозы в процессе клеточного дыхания:
C6H12O6 + 6O2 + 38H3PO4 + 38АДФ → 6CO2 + 44H2O + 38АТФ.
Таким образом, в ходе гликолиза образуются 2 молекулы АТФ, в ходе клеточного дыхания — ещё 36 молекул АТФ, в целом при пол- ном окислении глюкозы — 38 молекул АТФ.
Пластический обмен
Пластический обмен, или ассимиляция, представляет собой совокупность реакций, обеспечивающих синтез сложных органических соединений из более простых (фотосинтез, хемосинтез, биосинтез белка и др.).
Гетеротрофные организмы строят собственные органические вещества из органических компонентов пищи. Гетеротрофная ассимиляция сводится, по существу, к перестройке молекул:
органические вещества пищи (белки, жиры, углеводы) → простые органические молекулы (аминокислоты, жирные кислоты, моносахариды) → макромолекулы тела (белки, жиры, углеводы).
Автотрофные организмы способны полностью самостоятельно синтезировать органические вещества из неорганических молекул, потребляемых из внешней среды. В процессе фото- и хемосинтеза происходит образование простых органических соединений, из которых в дальнейшем синтезируются макромолекулы:
неорганические вещества (СО2, Н2О) → простые органические молекулы (аминокислоты, жирные кислоты, моносахариды) → макромолекулы тела (белки, жиры, углеводы).
Фотосинтез
Фотосинтез — синтез органических соединений из неорганических за счёт энергии света. Суммарное уравнение фотосинтеза:
Фотосинтез протекает при участии фотосинтезирующих пигментов, обладающих уникальным свойством преобразования энергии солнечного света в энергию химической связи в виде АТФ. Фотосинтезирующие пигменты представляют собой белковоподобные вещества. Наиболее важным является пигмент хлорофилл. У эукариот фотосинтезирующие пигменты встроены во внутреннюю мембрану пластид, у прокариот — во впячивания цитоплазматической мембраны.
Строение хлоропласта очень похоже на строение митохондрии. Во внутренней мембране тилакоидов гран содержатся фотосинтетические пигменты, а также белки цепи переноса электронов и молекулы фермента АТФ-синтетазы.
Процесс фотосинтеза состоит из двух фаз: световой и темновой.
1. Световая фаза фотосинтеза протекает только на свету в мембране тилакоидов граны.
К ней относятся поглощение хлорофиллом квантов света, образование молекулы АТФ и фотолиз воды.
Под действием кванта света (hv) хлорофилл теряет электроны, переходя в возбуждённое состояние:
Эти электроны передаются переносчиками на наружную, то есть обращенную к матриксу поверхность мембраны тилакоидов, где накапливаются.
Одновременно внутри тилакоидов происходит фотолиз воды, то есть её разложение под действием света:
Образующиеся электроны передаются переносчиками к молекулам хлорофилла и восстанавливают их. Молекулы хлорофилла возвращаются в стабильное состояние.
Протоны водорода, образовавшиеся при фотолизе воды, накапливаются внутри тилакоида, создавая Н+-резервуар. В результате внутренняя поверхность мембраны тилакоида заряжается положительно (за счёт Н+), а наружная — отрицательно (за счёт е—). По мере накопления по обе стороны мембраны противоположно заряженных частиц нарастает разность потенциалов. При достижении критической величины разности потенциалов сила электрического поля начинает проталкивать протоны через канал АТФ-синтетазы. Выделяющаяся при этом энергия используется для фосфорилирования молекул АДФ:
АДФ + Ф → АТФ.
Образование АТФ в процессе фотосинтеза под действием энергии света называется фотофосфорилированием.
Ионы водорода, оказавшись на наружной поверхности мембраны тилакоида, встречаются там с электронами и образуют атомарный водород, который связывается с молекулой-переносчиком водорода НАДФ (никотинамидадениндинуклеотидфосфат):
2Н+ + 4е– + НАДФ+ → НАДФ·Н2.
Таким образом, во время световой фазы фотосинтеза происходят три процесса: образование кислорода вследствие разложения воды, синтез АТФ и образование атомов водорода в форме НАДФ·Н2. Кислород диффундирует в атмосферу, а АТФ и НАДФ·Н2 участвуют в процессах темновой фазы.
2. Темновая фаза фотосинтеза протекает в матриксе хлоропласта как на свету, так и в темноте и представляет собой ряд последовательных преобразований СО2, поступающего из воздуха, в цикле Кальвина. Осуществляются реакции темновой фазы за счёт энергии АТФ. В цикле Кальвина СО2 связывается с водородом из НАДФ·Н2 с образованием глюкозы.
В процессе фотосинтеза кроме моносахаридов (глюкоза и др.) синтезируются мономеры других органических соединений — аминокислоты, глицерин и жирные кислоты. Таким образом, благодаря фотосинтезу растения обеспечивают себя и всё живое на Земле необходимыми органическими веществами и кислородом.
Сравнительная характеристика фотосинтеза и дыхания эукариот представлена в таблице.
Сравнительная характеристика фотосинтеза и дыхания эукариот
| Признак | Фотосинтез | Дыхание |
| Уравнение реакции | 6СО2 + 6Н2О + энергия света → C6H12O6 + 6O2 | C6H12O6 + 6O2 → 6СО2 + 6Н2О + энергия (АТФ) |
| Исходные вещества | Углекислый газ, вода | Органические вещества, кислород |
| Продукты реакции | Органические вещества, кислород | Углекислый газ, вода |
| Значение в круговороте веществ | Синтез органических веществ из неорганических | Разложение органических веществ до неорганических |
| Превращение энергии | Превращение энергии света в энергию химических связей органических веществ | Превращение энергии химических связей органических веществ в энергию макроэргических связей АТФ |
| Важнейшие этапы | Световая и темновая фаза (включая цикл Кальвина) | Неполное окисление (гликолиз) и полное окисление (включая цикл Кребса) |
| Место протекания процесса | Хлоропласты | Гиалоплазма (неполное окисление) и митохондрии (полное окисление) |
Генетическая информация у всех организмов хранится в виде определённой последовательности нуклеотидов ДНК (или РНК у РНК-содержащих вирусов). Прокариоты содержат генетическую информацию в виде одной молекулы ДНК. В эукариотических клетках генетический материал распределён в нескольких молекулах ДНК, организованных в хромосомы.
ДНК состоит из кодирующих и некодирующих участков. Кодирующие участки кодируют РНК. Некодирующие области ДНК выполняют структурную функцию, позволяя участкам генетического материала упаковываться определённым образом, или регуляторную функцию, участвуя во включении генов, направляющих синтез белка.
Кодирующими участками ДНК являются гены. Ген — участок молекулы ДНК, кодирующей синтез одной мРНК (и соответственно полипептида), рРНК или тРНК.
Участок хромосомы, где расположен ген называется локусом. Совокупность генов клеточного ядра представляет собой генотип, совокупность генов гаплоидного набора хромосом — гено́м, совокупность генов внеядерных ДНК (митохондрий, пластид, цитоплазмы) — плазмон.
Реализация информации, записанной в генах, через синтез белков называется экспрессией (проявлением) генов. Генетическая информация хранится в виде определённой последовательности нуклеотидов ДНК, а реализуется в виде последовательности аминокислот в белке. Посредниками, переносчиками информации выступают РНК. То есть реализация генетической информации происходит следующим образом:
ДНК → РНК → белок.
Этот процесс осуществляется в два этапа:
1) транскрипция;
2) трансляция.
Транскрипция (от лат. transcriptio — переписывание) — синтез РНК с использованием ДНК в качестве матрицы. В результате образуются мРНК, тРНК и рРНК. Процесс транскрипции требует больших затрат энергии в виде АТФ и осуществляется ферментом РНК-полимеразой.
Одновременно транскрибируется не вся молекула ДНК, а лишь отдельные её отрезки. Такой отрезок (транскриптон) начинается промотором — участком ДНК, куда присоединяется РНК-полимераза и откуда начинается транскрипция, а заканчивается терминатором — участком ДНК, содержащим сигнал окончания транскрипции. Транскриптон — это ген с точки зрения молекулярной биологии.
Транскрипция, как и репликация, основана на способности азотистых оснований нуклеотидов к комплементарному связыванию. На время транскрипции двойная цепь ДНК разрывается, и синтез РНК осуществляется по одной цепи ДНК.
В процессе транскрипции последовательность нуклеотидов ДНК переписывается на синтезирующуюся молекулу мРНК, которая выступает в качестве матрицы в процессе биосинтеза белка.
Гены прокариот состоят только из кодирующих нуклеотидных последовательностей.
Гены эукариот состоят из чередующихся кодирующих (экзонов) и некодирующих (интронов) участков.
После транскрипции участки мРНК, соответствующие интронам, удаляются в ходе сплайсинга, являющегося составной частью процессинга.
Процессинг — процесс формирования зрелой мРНК из её предшественника пре-мРНК. Он включает два основных события. 1.Присоединение к концам мРНК коротких последовательностей нуклеотидов, обозначающих место начала и место конца трансляции. Сплайсинг — удаление неинформативных последовательностей мРНК, соответствующих интронам ДНК. В результате сплайсинга молекулярная масса мРНК уменьшается в 10 раз.
Трансляция (от лат. translatio — перевод) — синтез полипептидной цепи с использованием мРНК в роли матрицы.
В трансляции участвуют все три типа РНК: мРНК является информационной матрицей; тРНК доставляют аминокислоты и узнают кодоны; рРНК вместе с белками образуют рибосомы, которые удерживают мРНК, тРНК и белок и осуществляют синтез полипептидной цепи.
Этапы трансляции
| Этап | Характеристика |
| Инициация | Сборка комплекса, участвующего в синтезе полипептидной цепи. Малая субчастица рибосомы соединяется с инициаторной мет-трнк, а затем с мрнк, после чего происходит образование целой рибосомы, состоящей из малой и большой субчастиц. |
| Элонгация | Удлинение полипептидной цепи. Рибосома перемещается вдоль мрнк, что сопровождается многократным повторением цикла присоединения очередной аминокислоты к растущей полипептидной цепи. |
| Терминация | Завершение синтеза полипептидной молекулы. Рибосома достигает одного из трёх стоп-кодонов мрнк, а так как не существует трнк с антикодонами, комплементарными стоп-кодонам, синтез полипептидной цепи прекращается. Она высвобождается и отделяется от рибосомы. Рибосомные субчастицы диссоциируют, отделяются от мрнк и могут принять участие в синтезе следующей полипептидной цепи. |
Реакции матричного синтеза. К реакциям матричного синтеза относятся
- самоудвоение ДНК (репликация);
- образование мРНК, тРНК и рРНК на молекуле ДНК (транскрипция);
- биосинтез белка на мРНК (трансляция).
Все эти реакции объединяет то, что молекула ДНК в одном случае или молекула мРНК в другом выступают в роли матрицы, на которой происходит образование одинаковых молекул. Реакции матричного синтеза являются основой способности живых организмов к воспроизведению себе подобных.
Регуляция экспрессии генов. Тело многоклеточного организма построено из разнообразных клеточных типов. Они отличаются структурой и функциями, то есть дифференцированы. Различия проявляются в том, что помимо белков, необходимых любой клетке организма, клетки каждого типа синтезируют ещё и специализированные белки: в эпидермисе образуется кератин, в эритроцитах — гемоглобин и т. д. Клеточная дифференцировка обусловлена изменением набора экспрессируемых генов и не сопровождается какими-либо необратимыми изменениями в структуре самих последовательностей ДНК.
Обмен веществ
Обмен веществ (метаболизм) складывается из процессов расщепления и синтеза — диссимиляции и ассимиляции, постоянно
протекающих в организме. Чтобы жизнь продолжалась, количество поступающей энергии должно превышать (или как минимум равняться)
количеству расходуемой энергии, поэтому диссимиляция и ассимиляция поддерживают определенный баланс друг с другом.

Энергетический обмен
Энергетический обмен (диссимиляция — от лат. dissimilis ‒ несходный) — обратная ассимиляции сторона обмена веществ, совокупность реакций, которые приводят к высвобождению энергии химических связей. Это реакции расщепления жиров,
белков, углеводов, нуклеиновых кислот до простых веществ.
Возможно три этапа диссимиляции: подготовительный, анаэробный и аэробный. Среда обитания определяет количество
этапов диссимиляции. Их может быть три, если организм обитает в кислородной среде, и два, если речь идет об
организме, обитающем в бескислородной среде (к примеру, в кишечнике).
Обсудим этапы энергетического обмена более подробно:
- Подготовительный этап
- Бескислородный этап (анаэробный) — гликолиз
- Кислородный этап (аэробный)
Подготовительный этап осуществляется ферментами в ЖКТ. В результате действия ферментов сложные вещества превращаются в более простые: полимеры распадаются на мономеры. Это сопровождается разрывом химических связей и выделением энергии, большая часть
которой рассеивается в виде тепла.
Под действием ферментов белки расщепляются на аминокислоты, жиры — на глицерин и жирные кислоты, сложные углеводы — до простых сахаров.
Этот этап является последним для организмов-анаэробов, обитающих в условиях, где кислород отсутствует. На этапе гликолиза
происходит расщепление молекулы глюкозы: образуется 2 молекулы АТФ и 2 молекулы пировиноградной кислоты (ПВК).
Происходит данный этап в цитоплазме клеток.
Этот этап доступен только для аэробов — организмов, живущих в кислородной среде. Из каждой молекулы ПВК, образовавшейся на
этапе гликолиза, синтезируется 18 молекул АТФ — в сумме с двух ПВК выход составляет 36 молекул АТФ.
Таким образом, суммарно с одной молекулы глюкозы можно получить 38 АТФ (гликолиз + кислородный этап).
Кислородный этап протекает на кристах митохондрий (складках, выпячиваниях внутренней мембраны), где наибольшая концентрация окислительных ферментов. Главную роль в этом процессе играет так называемый цикл Кребса, который подробно изучает биохимия.
АТФ — аденозинтрифосфорная кислота
Трудно переоценить роль в клетке АТФ — универсального источника энергии. Молекула АТФ состоит из азотистого основания —
аденина, углевода — рибозы и трех остатков фосфорной кислоты.
Между остатками фосфорной кислоты находятся макроэргические связи — ковалентные связи, которые гидролизуются с выделением
большого количества энергии. Их принято обозначать типографическим знаком тильда «∽».
АТФ гидролизуется до АДФ (аденозиндифосфорная кислота), а затем и до АМФ (аденозинмонофосфорная кислота).
Гидролиз АТФ сопровождается выделением энергии (E) на каждом этапе и может быть представлен такой схемой:
- АТФ + H2O = АДФ + H3PO4 + E
- АДФ + H2O = АМФ + H3PO4 + E
- АМФ + H2O = аденин + рибоза + H3PO4 + E
Пластический обмен
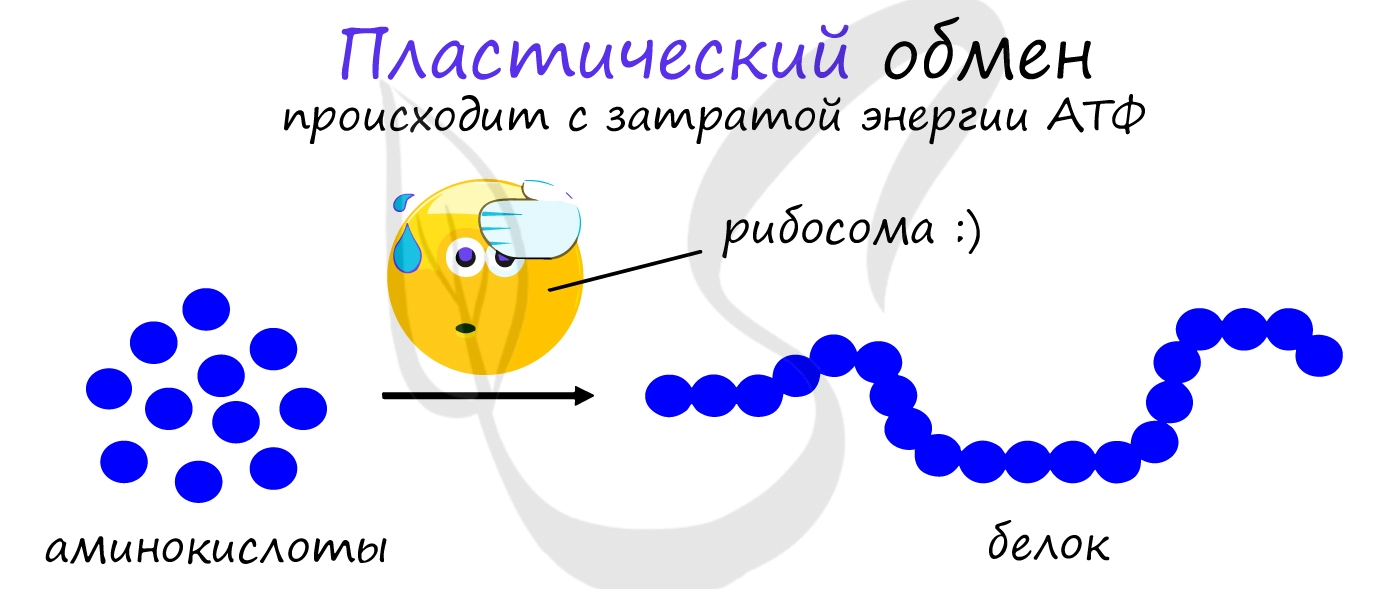
АТФ является универсальным источником энергии в клетке: энергия макроэргических связей АТФ используется для реакций
пластического обмена (ассимиляции), протекающих с затратой энергии: синтеза белка на рибосоме (трансляции),
удвоению ДНК (репликации) и т.д.
В результате пластического обмена в нашем организме происходит синтез белков, жиров и углеводов.
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Энергетический обмен
Энергетический обмен
(катаболизм, диссимиляция) — совокупность реакций расщепления органических
веществ, сопровождающихся выделением энергии. Энергия, освобождающаяся при
распаде органических веществ, не сразу используется клеткой, а запасается в
форме АТФ и других высокоэнергетических соединений. АТФ — универсальный
источник энергообеспечения клетки. Синтез АТФ происходит в клетках всех
организмов в процессе фосфорилирования — присоединения неорганического фосфата
к АДФ.
У аэробных
организмов (живущих в кислородной среде) выделяют три этапа энергетического
обмена: подготовительный, бескислородное окисление и кислородное окисление; у анаэробных
организмов (живущих в бескислородной среде) и аэробных при недостатке кислорода
— два этапа: подготовительный, бескислородное окисление.
Подготовительный этап
Заключается в
ферментативном расщеплении сложных органических веществ до простых: белковые
молекулы — до аминокислот, жиры — до глицерина и карбоновых кислот, углеводы —
до глюкозы, нуклеиновые кислоты — до нуклеотидов. Распад высокомолекулярных
органических соединений осуществляется или ферментами желудочно-кишечного
тракта или ферментами лизосом. Вся высвобождающаяся при этом энергия
рассеивается в виде тепла. Образовавшиеся небольшие органические молекулы могут
быть использованы в качестве «строительного материала» или могут подвергаться
дальнейшему расщеплению.
Бескислородное окисление, или гликолиз
Этот этап заключается в
дальнейшем расщеплении органических веществ, образовавшихся во время
подготовительного этапа, происходит в цитоплазме клетки и в присутствии
кислорода не нуждается. Главным источником энергии в клетке является глюкоза.
Процесс бескислородного неполного расщепления глюкозы — гликолиз.
Потеря электронов
называется окислением, приобретение — восстановлением, при этом донор
электронов окисляется, акцептор восстанавливается.
Следует отметить, что
биологическое окисление в клетках может происходить как с участием кислорода:
А + О2
→ АО2,
так и без его участия,
за счет переноса атомов водорода от одного вещества к другому. Например,
вещество «А» окисляется за счет вещества «В»:
АН2
+ В → А + ВН2
или за счет переноса
электронов, например, двухвалентное железо окисляется до трехвалентного:
Fe2+
→ Fe3+ + e—.
Гликолиз — сложный
многоступенчатый процесс, включающий в себя десять реакций. Во время этого
процесса происходит дегидрирование глюкозы, акцептором водорода служит
кофермент НАД+ (никотинамидадениндинуклеотид). Глюкоза в результате
цепочки ферментативных реакций превращается в две молекулы пировиноградной
кислоты (ПВК), при этом суммарно образуются 2 молекулы АТФ и восстановленная
форма переносчика водорода НАД·Н2:
С6Н12О6
+ 2АДФ + 2Н3РО4 + 2НАД+ → 2С3Н4О3
+ 2АТФ + 2Н2О + 2НАД·Н2.
Дальнейшая судьба ПВК
зависит от присутствия кислорода в клетке. Если кислорода нет, у дрожжей и
растений происходит спиртовое брожение, при котором сначала происходит
образование уксусного альдегида, а затем этилового спирта:
1.
С3Н4О3 → СО2 + СН3СОН,
2.
СН3СОН + НАД·Н2 → С2Н5ОН
+ НАД+.
3. У
животных и некоторых бактерий при недостатке кислорода происходит молочнокислое
брожение с образованием молочной кислоты:
4.
С3Н4О3 + НАД·Н2 → С3Н6О3
+ НАД+.
5. В
результате гликолиза одной молекулы глюкозы высвобождается 200 кДж, из которых
120 кДж рассеивается в виде тепла, а 80% запасается в связях АТФ.
6. Кислородное
окисление, или дыхание
7. Заключается
в полном расщеплении пировиноградной кислоты, происходит в митохондриях и при
обязательном присутствии кислорода.
8. Пировиноградная
кислота транспортируется в митохондрии (строение и функции митохондрий ). Здесь
происходит дегидрирование (отщепление водорода) и декарбоксилирование
(отщепление углекислого газа) ПВК с образованием двухуглеродной ацетильной
группы, которая вступает в цикл реакций, получивших название реакций цикла
Кребса. Идет дальнейшее окисление, связанное с дегидрированием и
декарбоксилированием. В результате на каждую разрушенную молекулу ПВК из
митохондрии удаляется три молекулы СО2; образуется пять пар атомов
водорода, связанных с переносчиками (4НАД·Н2, ФАД·Н2), а
также одна молекула АТФ.

Суммарная реакция
гликолиза и разрушения ПВК в митохондриях до водорода и углекислого газа
выглядит следующим образом:
С6Н12О6
+ 6Н2О → 6СО2 + 4АТФ + 12Н2.
Две молекулы АТФ
образуются в результате гликолиза, две — в цикле Кребса; две пары атомов
водорода (2НАДЧН2) образовались в результате гликолиза, десять пар — в цикле
Кребса.
Последним этапом
является окисление пар атомов водорода с участием кислорода до воды с
одновременным фосфорилированием АДФ до АТФ. Водород передается трем большим
ферментным комплексам (флавопротеины, коферменты Q, цитохромы) дыхательной
цепи, расположенным во внутренней мембране митохондрий. У водорода отбираются
электроны, которые в матриксе митохондрий в конечном итоге соединяются с
кислородом:
О2
+ e— → О2—.
Протоны закачиваются в
межмембранное пространство митохондрий, в «протонный резервуар». Внутренняя
мембрана непроницаема для ионов водорода, с одной стороны она заряжается
отрицательно (за счет О2—), с другой — положительно (за
счет Н+). Когда разность потенциалов на внутренней мембране
достигает 200 мВ, протоны проходят через канал фермента АТФ-синтетазы,
образуется АТФ, а цитохромоксидаза катализирует восстановление кислорода до
воды. Так в результате окисления двенадцати пар атомов водорода образуется 34
молекулы АТФ.

1 —
наружная мембрана; 2 — межмембранное пространство, протонный резервуар;
3 — цитохромы; 4 — АТФ-синтетаза.
При перфорации
внутренних митохондриальных мембран окисление НАД·Н2 продолжается,
но АТФ-синтетаза не работает и образования АТФ в дыхательной цепи не
происходит, энергия рассеивается в форме тепла (клетки «бурого жира»
млекопитающих).
Суммарная реакция
расщепления глюкозы до углекислого газа и воды выглядит следующим образом:
С6Н12О6
+ 6О2 → 6СО2 + 6Н2О + 38АТФ + Qт,
где Qт — тепловая
энергия.
Фотосинтез
— синтез органических веществ из углекислого газа и воды с обязательным
использованием энергии света:
6СО2
+ 6Н2О + Qсвета → С6Н12О6
+ 6О2.
У высших растений
органом фотосинтеза является лист, органоидами фотосинтеза — хлоропласты
(строение хлоропластов — лекция
№7). В мембраны тилакоидов хлоропластов встроены фотосинтетические
пигменты: хлорофиллы и каротиноиды. Существует несколько разных типов
хлорофилла (a, b, c, d), главным является хлорофилл a. В молекуле
хлорофилла можно выделить порфириновую «головку» с атомом магния в центре и
фитольный «хвост». Порфириновая «головка» представляет собой плоскую структуру,
является гидрофильной и поэтому лежит на той поверхности мембраны, которая
обращена к водной среде стромы. Фитольный «хвост» — гидрофобный и за счет этого
удерживает молекулу хлорофилла в мембране.

Хлорофиллы поглощают
красный и сине-фиолетовый свет, отражают зеленый и поэтому придают растениям
характерную зеленую окраску. Молекулы хлорофилла в мембранах тилакоидов
организованы в фотосистемы. У
растений и синезеленых водорослей имеются фотосистема-1 и фотосистема-2, у
фотосинтезирующих бактерий — фотосистема-1. Только фотосистема-2 может
разлагать воду с выделением кислорода и отбирать электроны у водорода воды.
Фотосинтез — сложный
многоступенчатый процесс; реакции фотосинтеза подразделяют на две группы:
реакции световой фазы и реакции темновой
фазы.
Световая фаза
Эта фаза происходит
только в присутствии света в мембранах тилакоидов при участии хлорофилла,
белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта
света электроны хлорофилла возбуждаются, покидают молекулу и попадают на
внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно.
Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды,
находящейся во внутритилакоидном пространстве. Это приводит к распаду или
фотолизу воды:
Н2О
+ Qсвета → Н+ + ОН—.
Ионы гидроксила отдают
свои электроны, превращаясь в реакционноспособные радикалы •ОН:
ОН—
→ •ОН + е—.
Радикалы •ОН
объединяются, образуя воду и свободный кислород:
4НО• →
2Н2О + О2.
Кислород при этом
удаляется во внешнюю среду, а протоны накапливаются внутри тилакоида в
«протонном резервуаре». В результате мембрана тилакоида с одной стороны за счет
Н+ заряжается положительно, с другой за счет электронов —
отрицательно. Когда разность потенциалов между наружной и внутренней сторонами
мембраны тилакоида достигает 200 мВ, протоны проталкиваются через каналы
АТФ-синтетазы и происходит фосфорилирование АДФ до АТФ; атомарный водород идет
на восстановление специфического переносчика НАДФ+
(никотинамидадениндинуклеотидфосфат) до НАДФ·Н2:
2Н+
+ 2е— + НАДФ → НАДФ·Н2.
Таким образом, в
световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими
процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3)
образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2
транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.

1 —
строма хлоропласта; 2 — тилакоид граны.
Темновая фаза
Эта фаза протекает в
строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они
происходят не только на свету, но и в темноте. Реакции темновой фазы
представляют собой цепочку последовательных преобразований углекислого газа
(поступает из воздуха), приводящую к образованию глюкозы и других органических
веществ.
Первая реакция в этой
цепочке — фиксация углекислого газа; акцептором углекислого газа является
пятиуглеродный сахар рибулозобифосфат
(РиБФ); катализирует реакцию фермент рибулозобифосфат-карбоксилаза
(РиБФ-карбоксилаза). В результате карбоксилирования рибулозобисфосфата
образуется неустойчивое шестиуглеродное соединение, которое сразу же
распадается на две молекулы фосфоглицериновой кислоты
(ФГК). Затем происходит цикл реакций, в которых через ряд промежуточных
продуктов фосфоглицериновая кислота преобразуется в глюкозу. В этих реакциях
используются энергии АТФ и НАДФ·Н2, образованных в световую фазу;
цикл этих реакций получил название «цикл Кальвина»:
6СО2
+ 24Н+ + АТФ → С6Н12О6 + 6Н2О.
Кроме глюкозы, в
процессе фотосинтеза образуются другие мономеры сложных органических соединений
— аминокислоты, глицерин и жирные кислоты, нуклеотиды. В настоящее время
различают два типа фотосинтеза: С3— и С4-фотосинтез.
С3-фотосинтез

Это тип фотосинтеза,
при котором первым продуктом являются трехуглеродные (С3)
соединения. С3-фотосинтез был открыт раньше С4-фотосинтеза
(М. Кальвин). Именно С3-фотосинтез описан выше, в рубрике «Темновая
фаза». Характерные особенности С3-фотосинтеза: 1) акцептором
углекислого газа является РиБФ, 2) реакцию карбоксилирования РиБФ катализирует
РиБФ-карбоксилаза, 3) в результате карбоксилирования РиБФ образуется
шестиуглеродное соединение, которое распадается на две ФГК. ФГК
восстанавливается до триозофосфатов
(ТФ). Часть ТФ идет на регенерацию РиБФ, часть превращается в глюкозу.
Фотодыхание

Фотодыхание:
1 — хлоропласт; 2 — пероксисома; 3 — митохондрия.
Это светозависимое
поглощение кислорода и выделение углекислого газа. Еще в начале прошлого века
было установлено, что кислород подавляет фотосинтез. Как оказалось, для
РиБФ-карбоксилазы субстратом может быть не только углекислый газ, но и
кислород:
О2
+ РиБФ → фосфогликолат (2С) + ФГК (3С).
Фермент при этом
называется РиБФ-оксигеназой. Кислород является конкурентным ингибитором
фиксации углекислого газа. Фосфатная группа отщепляется, и фосфогликолат
становится гликолатом, который растение должно утилизировать. Он поступает в
пероксисомы, где окисляется до глицина. Глицин поступает в митохондрии, где
окисляется до серина, при этом происходит потеря уже фиксированного углерода в
виде СО2. В итоге две молекулы гликолата (2С + 2С) превращаются в
одну ФГК (3С) и СО2. Фотодыхание приводит к понижению урожайности С3-растений
на 30–40% (С3-растения — растения, для
которых характерен С3-фотосинтез).
С4-фотосинтез
С4-фотосинтез
— фотосинтез, при котором первым продуктом являются четырехуглеродные (С4)
соединения. В 1965 году было установлено, что у некоторых растений (сахарный
тростник, кукуруза, сорго, просо) первыми продуктами фотосинтеза являются
четырехуглеродные кислоты. Такие растения назвали С4-растениями.
В 1966 году австралийские ученые Хэтч и Слэк показали, что у С4-растений
практически отсутствует фотодыхание и они гораздо эффективнее поглощают углекислый
газ. Путь превращений углерода в С4-растениях стали называть путем
Хэтча-Слэка.
Для С4-растений
характерно особое анатомическое строение листа. Все проводящие пучки окружены
двойным слоем клеток: наружный — клетки мезофилла, внутренний — клетки обкладки.
Углекислый газ фиксируется в цитоплазме клеток мезофилла, акцептор — фосфоенолпируват
(ФЕП, 3С), в результате карбоксилирования ФЕП образуется оксалоацетат (4С).
Процесс катализируется ФЕП-карбоксилазой.
В отличие от РиБФ-карбоксилазы ФЕП-карбоксилаза обладает большим сродством к СО2
и, самое главное, не взаимодействует с О2. В хлоропластах мезофилла
много гран, где активно идут реакции световой фазы. В хлоропластах клеток
обкладки идут реакции темновой фазы.
Оксалоацетат (4С)
превращается в малат, который через плазмодесмы транспортируется в клетки
обкладки. Здесь он декарбоксилируется и дегидрируется с образованием пирувата,
СО2 и НАДФ·Н2.
Пируват возвращается в
клетки мезофилла и регенерирует за счет энергии АТФ в ФЕП. СО2 вновь
фиксируется РиБФ-карбоксилазой с образованием ФГК. Регенерация ФЕП требует
энергии АТФ, поэтому нужно почти вдвое больше энергии, чем при С3-фотосинтезе.
|
|
Строение С4-растений: |
|
|
С4-фотосинтез: |


Значение фотосинтеза
Благодаря фотосинтезу,
ежегодно из атмосферы поглощаются миллиарды тонн углекислого газа, выделяются
миллиарды тонн кислорода; фотосинтез является основным источником образования
органических веществ. Из кислорода образуется озоновый слой, защищающий живые
организмы от коротковолновой ультрафиолетовой радиации.
При фотосинтезе зеленый
лист использует лишь около 1% падающей на него солнечной энергии,
продуктивность составляет около 1 г органического вещества на 1 м2
поверхности в час.
Хемосинтез
Синтез органических
соединений из углекислого газа и воды, осуществляемый не за счет энергии света,
а за счет энергии окисления неорганических веществ, называется хемосинтезом.
К хемосинтезирующим организмам относятся некоторые виды бактерий.
Нитрифицирующие
бактерии окисляют аммиак до азотистой, а затем до азотной кислоты (NH3
→ HNO2 → HNO3).
Железобактерии
превращают закисное железо в окисное (Fe2+ → Fe3+).
Серобактерии
окисляют сероводород до серы или серной кислоты (H2S + ½O2
→ S + H2O, H2S + 2O2 → H2SO4).
В результате реакций
окисления неорганических веществ выделяется энергия, которая запасается
бактериями в форме макроэргических связей АТФ. АТФ используется для синтеза
органических веществ, который проходит аналогично реакциям темновой фазы
фотосинтеза.
Хемосинтезирующие
бактерии способствуют накоплению в почве минеральных веществ, улучшают
плодородие почвы, способствуют очистке сточных вод и др.
Метаболизм состоит из двух взаимно противоположных, но взаимосвязанных процессов пластического и энергетического обмена.
Энергетический обмен необходим организму для образования энергии, которая, в свою очередь, будет израсходована на важные биологические процессы, происходящие в клетках, тканях, органах, в том числе и на пластический обмен.
Все наши движения, мыслительные и физиологические процессы (пищеварение, кровообращение, выделение), любое проявление жизнедеятельности требуют затрат энергии.
Энергетический обмен также называют катаболизм или диссимиляцией. Это достаточно длительный процесс, который происходит вплоть до того момента, пока все питательные вещества, поступившие в организм, не расщепятся до углекислого газа, воды или других простых соединений, которые организм уже не сможет использовать.
Этот процесс аналогичен горению, при котором выделяется вода, углекислый газ и огромное количество энергии.
Катаболизм- это прежде всего многоступенчатый процесс, он не нуждается в высоких температурах, а выделившаяся энергия по большей части не переходит в тепловую, чтобы безвозвратно рассеяться, а запасается для дальнейших нужд в виде молекул АТФ.
Все это делает этот процесс невероятно эффективным и уникальным!
Эта информация доступна зарегистрированным пользователям
Энергетический обмен— это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ.
Каким же образом энергия реакции расщепления используется клеткой?
Ученые обнаружили, что любая деятельность клетки всегда точно совпадает во времени с распадом молекул АТФ.
К примеру, при синтезе белков, углеводов, жиров в клетке идет активный распад АТФ.
В результате опытов было обнаружено, что любая работа мышц сопровождается активным расщеплением АТФ в их клетках.
Ученые сделали вывод, что именно АТФ является непосредственным источником энергии, необходимой для сокращения мышц и для синтеза сложных соединений.
Известно, что в среднем содержание АТФ в клетках составляет от 0,05% до 0,5% ее массы, то есть запас молекул АТФ в организме ограничен и после распада АТФ должно произойти его восстановление.
Многоуровневый процесс энергетического обмена- это последовательные реакции восстановления молекул АТФ, которые происходят при участии ферментов.
Это можно сравнить с аккумулятором для телефона: когда его заряд садится, то устройство необходимо вновь зарядить.
Если в клетке постоянно измерять содержание АТФ, то его количество существенно не изменяется, но количество углеводов, белков, жиров будет уменьшаться. Это объясняется тем, что реакции расщепления углеводов, белков, жиров и других веществ обеспечивают быстрое и полное восстановление израсходованной АТФ.
В каждой клетке нашего организма в течение суток АТФ примерно 10 тысяч раз распадается и вновь заново образуется.
Таким образом, АТФ- это единый и универсальный источник энергии для функциональной деятельности клетки.
Следует отметить, что возможна передача энергии из одних частей клетки в другие.
Синтез АТФ может происходить в одном месте и в одно время, а использоваться может в другом месте и в другое время.
Синтез АТФ в основном происходит в митохондриях. Образовавшаяся там АТФ по каналам эндоплазматической сети направляется в те места клетки, где возникает потребность в энергии.
Эта информация доступна зарегистрированным пользователям
Это одно из проявлений высочайшей организованности и упорядоченности всех химических реакций, протекающих в клетке.
Растения могут преобразовывать энергию солнечных лучей в АТФ на первом этапе фотосинтеза. Хемосинтезирующие бактерии способны запасать энергию в форме АТФ, получаемую при реакциях окисления различных неорганических соединений.
Следует отметить, что фотосинтезирующие и хемосинтезирующие организмы также способны получать энергию благодаря окислению органических веществ, синтезированных в собственных клетках из неорганических соединений.
У гетеротрофов (животных, грибов) образование АТФ идет в клетках при помощи реакций окисления органических веществ, поступающих вместе с пищей.
В клетках растений:
Крахмал →глюкоза → АТФ
В клетках животных:
гликоген → глюкоза → АТФ
Энергетический обмен делится на три последовательных этапа:
- подготовительный этап
- бескислородный этап
- кислородный этап
Подготовительный этап
Вся пища, которая поступает в наш организм, подвергается ферментативному расщеплению, при котором:
- белки расщепляются до аминокислот
- липиды расщепляются до глицерина и жирных кислот
- сложные углеводы (крахмал) расщепляются до глюкозы
На этом этапе вся выделившаяся при расщеплении веществ энергия рассеивается в виде тепла.
Эта информация доступна зарегистрированным пользователям
У одноклеточных животных подготовительный этап протекает в клетках, где и происходит расщепление сложных органических веществ на простые вещества под действием ферментов лизосом.
У многоклеточных организмов расщепление веществ начинает происходить в пищеварительном канале, а далее в клетках под действием лизосом.
Эта информация доступна зарегистрированным пользователям
Образовавшиеся небольшие органические молекулы могут быть использованы в качестве «строительного материала» или могут подвергаться дальнейшему расщеплению (гликолизу).
Вывод: на первом этапе энергетического обмена происходит распад сложных органических веществ на простые, с выделением энергии, которая вся рассеивается в виде тепла.
Эта информация доступна зарегистрированным пользователям
Ключевое место в метаболизме всех типов клеток занимают реакции с участием сахаров, например, глюкозы, потому что процесс расщепления глюкозы идет наиболее быстро и легче, ведь организму необходимо достаточно быстро восстанавливать энергетические затраты.
Аминокислоты и белки использовать для образования энергии слишком не выгодно, так как большая их часть является структурными компонентами клеток. В этом случае организм разрушал бы сам себя.
Жиры могут использоваться для получения энергии, но главным образом после того, как израсходовались запасы углеводов, ведь жиры из-за своей гидрофобности очень медленно окисляются и малоподвижны в клетках. При этом из жиров в отсутствие кислорода АТФ получить нельзя, а из глюкозы можно.
Поэтому организм выбирает наиболее выгодный путь получения энергии в виде молекул АТФ за счет расщепления, в первую очередь, глюкозы.
Второй этап энергетического обмена называют бескислородным, так как процесс расщепления глюкозы и образования молекул АТФ идет без участия кислорода.
Гликолиз (от греч. «гликос» сладкий, «лизис»- расщепление) — последовательное расщепление глюкозы.
Гликолиз идет в цитоплазме клеток без участия кислорода. Он состоит из последовательных реакций, каждая из которых катализируется общим ферментом.
В ходе реакций гликолиза молекула глюкозы С6Н12О6 распадается на две трехуглеродные молекулы пировиноградной кислоты (ПВК)— С3Н4О3, при этом суммарно образуются две молекулы АТФ и вода.
Акцептором (лат. accipio- «я принимаю, получаю») водорода в реакции гликолиза служит кофермент НАД+.
НАД+ (никотинамидадениндинуклеотид) — кофермент, имеющийся во всех живых клетках.
НАД+ переносит электроны из одной реакции в другую.
НАД+ является окислителем и забирает электрон от другой молекулы и один водород, восстанавливаясь в НАД H, который далее служит восстановителем и уже отдаёт электроны.
Уравнение реакции гликолиза:
Эта информация доступна зарегистрированным пользователям
Дальнейшая судьба ПВК может быть различной и зависит от того, какой тип извлечения энергии предпочитают организмы: анаэробный (бескислородный) или аэробный (кислородный).
Например, паразитические черви, живущие в кишечнике организмов хозяев, выбирают бескислородный путь преобразования ПВК, так как они мало подвижны и их клеткам хватает энергии, которая образуется при гликолизе глюкозы.
Эти виды паразитов выбирают именно такой путь преобразования энергии еще и потому, что при распаде глюкозы образуются ядовитые вещества (ацетон, уксусная кислота и этиловый спирт), которые действуют угнетающе на организм хозяина и ослабляют его иммунитет, что, в свою очередь, помогает паразиту существовать в агрессивной для него среде.
Эта информация доступна зарегистрированным пользователям
Образовавшаяся в результате гликолиза пировиноградная кислота подвергается дальнейшему преобразованию уже на внутренней мембране митохондрий, то есть переходит на третий этап энергетического обмена.
Вывод: на втором этапе энергетического обмена, гликолизе, из 1 молекулы глюкозы образуется 2 молекулы ПВК и 2 молекулы АТФ.
Эта информация доступна зарегистрированным пользователям
Если в клетку прекратилась подача кислорода, то ПВК подвергается брожению, к примеру, в клетках растений, которые были затоплены во время весенних паводков.
В зависимости от того, какие конечные продукты образуются, выделяют несколько видов брожения.
Рассмотрим основные виды:
1. Спиртовое брожение
Встречается в основном у дрожжей и растений.
Конечными продуктами являются этанол и углекислый газ.
Дрожжи:
Эта информация доступна зарегистрированным пользователям
При доступе кислорода процесс брожения ослабевает, на смену ему приходит дыхание.
Подавление спиртового брожения кислородом называется эффектом Пастера.
Спиртовое брожение используется в пищевой промышленности: хлебопечении, виноделии.
При этом типе брожения сначала происходит образование уксусного альдегида, а затем этилового спирта:
Эта информация доступна зарегистрированным пользователям
2. Молочнокислое брожение
Осуществляется с помощью лактобактерий, бифидобактерий, стрептококков.
Из ПВК они образуют молочную кислоту, ацетон, янтарную и уксусную кислоту.
Молочнокислые бактерии широко используются в молочной промышленности для получения молочнокислых продуктов, а также в создании пробиотиков.
Эта информация доступна зарегистрированным пользователям
У животных и человека при недостатке кислорода также может происходить молочнокислое брожение с образованием молочной кислоты.
В мышцах есть запасы углеводов в виде гликогена. При долгой и усиленной работе кровь не успевает снабдить мышцы достаточным количеством кислорода, в результате чего мышечные клетки вынуждены переходить на бескислородный способ получения АТФ.
При этом образуется молочная кислота, вызывающая боли в мышцах.
Эта информация доступна зарегистрированным пользователям
Квашение- разновидность молочнокислого брожения, в процессе которого образуется молочная кислота, оказывающая на продукты (наряду с добавляемой поваренной солью) консервирующее и размягчающее действие.
Квашение применяется при консервировании овощей и в кожевенном производстве.
Эта информация доступна зарегистрированным пользователям
3. Маслянокислое брожение
Масляная кислота, бутанол, ацетон, уксусная и ряд других органических кислот являются продуктами сбраживания углеводов бактериями- сахаролитическими анаэробами.
Благодаря определению наличия тех или иных кислот в клетке можно установить, какие бактерии образовали эти кислоты.
Знание механизмов брожения имеет большое практическое значение не только для живых организмов, но и для человека:
- для разработки методов диагностики инфекционных заболеваний, по набору ферментов или кислот, которые образовались во время брожения
- для создания современных биотехнологий молочнокислых продуктов, сыра, хлеба, вина и многих других продуктов питания
Недостатком процессов брожения является извлечение незначительной доли той энергии, которая заключена в связях органических молекул.
Для бактерий, паразитических видов, живущих в бескислородной среде, энергии, образующейся в результате брожения или гликолиза, достаточно для существования, поэтому они, в отличие от человека, не нуждаются в кислороде.
Также брожение является жизненно важным процессом для хвойных растений. В зимний период устьица хвои закупориваются смолой и газообмен с окружающей средой практически прекращается, в этом случае для получения энергии в клетках активно идет процесс спиртового брожения.
Эта информация доступна зарегистрированным пользователям
Эта информация доступна зарегистрированным пользователям
Окислительное фосфорилирование (дыхание)- процесс синтеза АТФ с обязательным участием комплекса ферментов, идет на внутренних мембранах митохондрий в присутствии кислорода.
Эта информация доступна зарегистрированным пользователям
Для нормального течения кислородного процесса необходима целостность мембран митохондрий.
В процессе такого клеточного дыхания энергия может переходить из химической в тепловую.
Кислородное дыхание гораздо эффективнее гликолиза, так как полное окисление органических веществ приводит к выделению большого количества энергии, причем примерно 60% ее запасается в молекулах АТФ, а 40% рассеивается в виде тепла.
Образовавшаяся в процессе гликолиза пировиноградная кислота поступает в митохондрии. Здесь она превращается в богатое энергией вещество ацетилкофермент А (Ацетил-КоА).
Ацетил-КоА взаимодействует с молекулой щавелевоуксусной кислоты, образуя лимонную кислоту, которая подвергается дальнейшим превращениям, заканчивающимся образованием щавелевоуксусной кислоты.
Эта кислота вновь взаимодействует с Ацетил-КоА, и цикл превращений повторяется.
Этот сложный процесс получил название цикл Кребса или цикл трикарбоновых кислот, еще его называют «циклом лимонной кислоты».
Цикл превращения трикарбоновых кислот открыл немецкий биохимик Ханс Кребс (1900- 1981), за что в 1953 году совместно с другими исследователями он получил Нобелевскую премию.
В цикле Кребса образуются молекулы СО2 (выводятся из клетки) и атомы водорода Н.
Специальные молекулы-переносчики НАД доставляют атомы Н к внутренней мембране митохондрий, имеющей внутреннюю и наружную поверхности.
Здесь атомы Н теряют электрон и превращаются в протоны Н+
Н — ē → Н+
Свободные электроны перемещаются по цепи переноса электронов на внутреннюю поверхность внутренней мембраны и присоединяются к атому кислорода О2:
О2 + ē= О2—
Далее 4 протона Н+соединяются с О2—в результате образуются две молекулы воды, которую еще называют метаболической водой, то есть выработанной организмом, а не полученной извне.
4Н+ + О2— = 2 Н2О + Q(т)
Обратите внимание, что в данной реакции образуется тепловая энергия (Qт)
Эта информация доступна зарегистрированным пользователям
Сиреневой стрелочкой показано, как белки дыхательной цепи забирают электроны у НАД Н.
Оказывается, белки дыхательной цепи за счет разницы энергии входящего электрона и выходящего могут протаскивать сквозь мембрану протоны Н+ в пространство между мембранами, где формируется положительный заряд.
Таким образом, между внутренней и наружной поверхностями внутренней мембраны митохондрий возникает разность потенциалов и формируется большая потенциальная энергия, за счет которой происходит образование АТФ.
Красная стрелка показывает, как белки дыхательной цепи переносят протоны Н+ в пространство между мембранами.
АТФ- синтаза (фермент, синтезируюший АТФ) пропускает протоны Н+ через свой активный центр и за счет этой энергии из АДФ и фосфорной кислоты образуется молекула АТФ.
В матриксе митохондрий протоны Н+соединяются с анионами О-2, образуя воду (Н2О).
АДФ + Ф → АТФ+ Н2О
Таким образом, в процессе дыхания в митохондриях образуются бедные энергией вещества: СО2 и Н2О, и освобождается большое количество энергии.
При полном расщеплении двух молекул ПВК, (синтезированных из одной молекулы глюкозы в процессе гликолиза), образуется 36 молекул АТФ.
Для большинства животных процессы дыхания являются основным путем получения энергии, необходимой для их жизнедеятельности.
ПВК + О2 → СО2 + Н2О + Qт (энергия)
Эта информация доступна зарегистрированным пользователям
Организм человека функционирует в аэробных условиях: 90% энергии он получает при участии кислорода.
Итак, повторим этапы энергетического обмена:
|
Подготовительный этап |
Бескислородный этап (гликолиз) |
Кислородный этап |
|
|
Где происходит |
В органах пищеварения, в клетках под действием ферментов |
Внутри клетки, в цитоплазме |
На внутренних мембранах митохондрий |
|
На какие вещества расщепляются соединения клетки |
Белки до аминокислот. Жиры до глицерина. Жирные кислоты и углеводы до глюкозы |
Глюкоза (С6Н12О6) до: ПВК (С3Н4О3) Энергия (Qт) |
ПВК (С3Н4О3) до: СО2 Н2О |
|
Сколько выделяется энергии |
Мало, рассеивается в виде тепла. |
40% энергии идет на синтез АТФ. 60% рассеивается в виде тепла |
Более 60% энергии запасается в виде АТФ |
|
Сколько синтезируется энергии в виде АТФ |
Не образуется |
2 молекулы АТФ |
36 молекул АТФ |
Общий вывод: объединяя анаэробные и аэробные процессы образования АТФ при расщеплении глюкозы, можно записать следующее суммарное уравнение:
Эта информация доступна зарегистрированным пользователям
Эта информация доступна зарегистрированным пользователям
Задача 1
В процессе гликолиза в клетках эукариот образовались 120 молекул пировиноградной кислоты (ПВК).
Какое количество молекул глюкозы подверглось расщеплению и сколько молекул АТФ образуется при полном ее окислении? Ответ поясните.
Решение:
1) В процессе гликолиза при расщеплении 1 молекулы глюкозы образуются 2 молекулы пировиноградной кислоты и выделяется энергия, которой хватает на синтез двух молекул АТФ
2) Так как образовалось 120 молекул пировиноградной кислоты, то расщеплению подверглось 120 : 2 = 60 молекул глюкозы
3) При полном окислении в расчете на одну молекулу глюкозы образуется 38 молекул АТФ
4) Следовательно, при полном окислении 60 молекул глюкозы образуются
38 х 60 = 2280 молекул АТФ
Задача 2
Гликолизу подверглись две молекулы глюкозы, окислению только одна.
Определите количество образовавшихся в результате этого процесса молекул АТФ и углекислого газа.
Решение:
Для решения используем уравнения 2 этапа (гликолиза) и 3 этапа (кислородного) энергетического обмена.
1) При гликолизе одной молекулы глюкозы образуются 2 молекулы АТФ
2) По условию задачи гликолизу подверглось 2 молекулы глюкозы:
2х2= 4 молекулы АТФ образовалось
3) При окислении (кислородный этап) из одной молекулы глюкозы образуются 36 молекул АТФ
4) По условию задачи окислению подверглась только одна молекула глюкозы:
1 х 36= 36 молекул АТФ образовалось
5) Таким образом образовалось 4 + 36 = 40 молекул АТФ
6) Углекислый газ образуется только на 3 этапе, при полном окислении одной молекулы глюкозы образуется 6 молекул СО2
Ответ: образовалось 40 молекул АТФ и 6 молекул СО2
Задача 3
В процессе окислительного фосфорилирования образовалось 1728 молекул АТФ.
Определите, какое количество молекул глюкозы подверглось расщеплению и сколько молекул АТФ образовалось в результате подготовительного и бескислородного этапов катаболизма.
Решение:
1) При окислительном фосфорилировании (третий этап энергетического обмена) из одной молекулы глюкозы образуется 36 молекул АТФ, следовательно, гидролизу подверглось:
1728 : 36 = 48 молекул глюкозы
2) При гликолизе она молекула глюкозы расщепляется до двух молекул пировиноградной кислоты (ПВК) с образованием двух молекул АТФ, следовательно, образовалось:
48 х 2 = 96 молекул АТФ
3) При полном окислении одной молекулы глюкозы образуется 38 молекул АТФ, следовательно, при полном окислении 48 молекул глюкозы образовалось:
48 х 38 = 1824 молекулы АТФ (1728 АТФ третьего этапа + 96 АТФ второго этапа = 1824)
Задача 4
Сколько молекул АТФ будет синтезировано в клетках в процессе гликолиза, если происходит окисление участка молекулы крахмала, содержащего 50 остатков глюкозы?
Решение:
1) В процессе гликолиза при окислении одной молекулы глюкозы образуется 2молекулы АТФ
2) Из фрагмента молекулы крахмала, содержащего 50 остатков глюкозы в результате гликолиза образуется 100 молекул АТФ (50 х 2 = 100)
Задача 5
В процессе полного расщепления глюкозы образовалось 342 молекулы АТФ.
Сколько молекул глюкозы поверглось расщеплению?
Сколько молекул АТФ образовалось в результате гликолиза?
Объясните полученный результат.
Решение:
1) В результате полного расщепления одной молекулы глюкозы образуется 38 молекул АТФ
2) Расщеплению поверглись 342 : 38 = 9 молекул глюкозы
3) Гликолиз– бескислородный этап катаболизма, в результате которого из одной молекулы глюкозы образуется 2 молекулы АТФ
4) Следовательно, в результате гликолиза образовалось 9 х 2 = 18 молекул АТФ
Задача 6
При беге со средней скоростью мышцы ног расходуют за 1 минуту 24 кДж энергии.
Определите сколько граммов глюкозы израсходуют мышцы ног за 25 минут бега, если кислород доставляется кровью к мышцам в достаточном количестве?
Для сведения: один моль содержит 180 грамм глюкозы и из этого количества образуется 38 молекул АТФ.
Решение:
1) Определим сколько всего энергии было израсходовано за 25 минут бега:
25 минут х 24 кДж = 600 кДж энергии
2) Определим сколько молекул АТФ образовалось, учитывая, что 1 молекула АТФ образует 40 кДж энергии:
600 : 40 = 15 молекул АТФ
3) Определим сколько глюкозы было израсходовано. Из условия ясно, что 1 моль глюкозы образует 38 молекул АТФ:
С6Н12О6 + 6 О2→ 6 СО2 + 6 Н2О + 38 АТФ
Составляем пропорцию:
1 моль (C6H12O6) — 38 (АТФ)
Х моль (C6H12O6) — 15 (АТФ)
Х=1 х 15/38
Х= 0,4 моль (C6H12O6)
4) Определяем, сколько грамм глюкозы содержится в 0,4 молях, для этого опять составляем пропорцию:
1 моль (C6H12O6) = 180 гр
0,4 моль (C6H12O6) = Х гр
Х=180 х 0,4/1
Х = 72 гр (C6H12O6)
Ответ:71 гр глюкозы израсходуют мышцы при беге продолжительность 25 минут
Эта информация доступна зарегистрированным пользователям