Обмен веществ
Обмен веществ (метаболизм) складывается из процессов расщепления и синтеза — диссимиляции и ассимиляции, постоянно
протекающих в организме. Чтобы жизнь продолжалась, количество поступающей энергии должно превышать (или как минимум равняться)
количеству расходуемой энергии, поэтому диссимиляция и ассимиляция поддерживают определенный баланс друг с другом.

Энергетический обмен
Энергетический обмен (диссимиляция — от лат. dissimilis ‒ несходный) — обратная ассимиляции сторона обмена веществ, совокупность реакций, которые приводят к высвобождению энергии химических связей. Это реакции расщепления жиров,
белков, углеводов, нуклеиновых кислот до простых веществ.
Возможно три этапа диссимиляции: подготовительный, анаэробный и аэробный. Среда обитания определяет количество
этапов диссимиляции. Их может быть три, если организм обитает в кислородной среде, и два, если речь идет об
организме, обитающем в бескислородной среде (к примеру, в кишечнике).
Обсудим этапы энергетического обмена более подробно:
- Подготовительный этап
- Бескислородный этап (анаэробный) — гликолиз
- Кислородный этап (аэробный)
Подготовительный этап осуществляется ферментами в ЖКТ. В результате действия ферментов сложные вещества превращаются в более простые: полимеры распадаются на мономеры. Это сопровождается разрывом химических связей и выделением энергии, большая часть
которой рассеивается в виде тепла.
Под действием ферментов белки расщепляются на аминокислоты, жиры — на глицерин и жирные кислоты, сложные углеводы — до простых сахаров.
Этот этап является последним для организмов-анаэробов, обитающих в условиях, где кислород отсутствует. На этапе гликолиза
происходит расщепление молекулы глюкозы: образуется 2 молекулы АТФ и 2 молекулы пировиноградной кислоты (ПВК).
Происходит данный этап в цитоплазме клеток.
Этот этап доступен только для аэробов — организмов, живущих в кислородной среде. Из каждой молекулы ПВК, образовавшейся на
этапе гликолиза, синтезируется 18 молекул АТФ — в сумме с двух ПВК выход составляет 36 молекул АТФ.
Таким образом, суммарно с одной молекулы глюкозы можно получить 38 АТФ (гликолиз + кислородный этап).
Кислородный этап протекает на кристах митохондрий (складках, выпячиваниях внутренней мембраны), где наибольшая концентрация окислительных ферментов. Главную роль в этом процессе играет так называемый цикл Кребса, который подробно изучает биохимия.
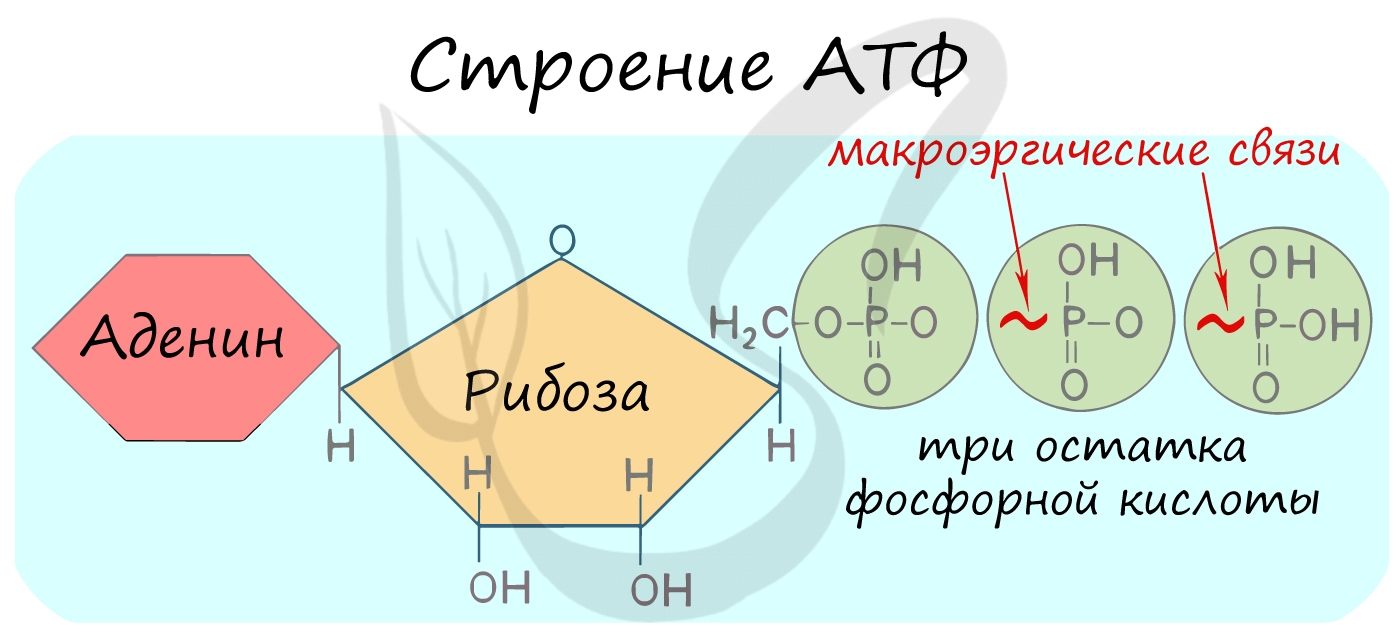
АТФ — аденозинтрифосфорная кислота
Трудно переоценить роль в клетке АТФ — универсального источника энергии. Молекула АТФ состоит из азотистого основания —
аденина, углевода — рибозы и трех остатков фосфорной кислоты.
Между остатками фосфорной кислоты находятся макроэргические связи — ковалентные связи, которые гидролизуются с выделением
большого количества энергии. Их принято обозначать типографическим знаком тильда «∽».
АТФ гидролизуется до АДФ (аденозиндифосфорная кислота), а затем и до АМФ (аденозинмонофосфорная кислота).
Гидролиз АТФ сопровождается выделением энергии (E) на каждом этапе и может быть представлен такой схемой:
- АТФ + H2O = АДФ + H3PO4 + E
- АДФ + H2O = АМФ + H3PO4 + E
- АМФ + H2O = аденин + рибоза + H3PO4 + E
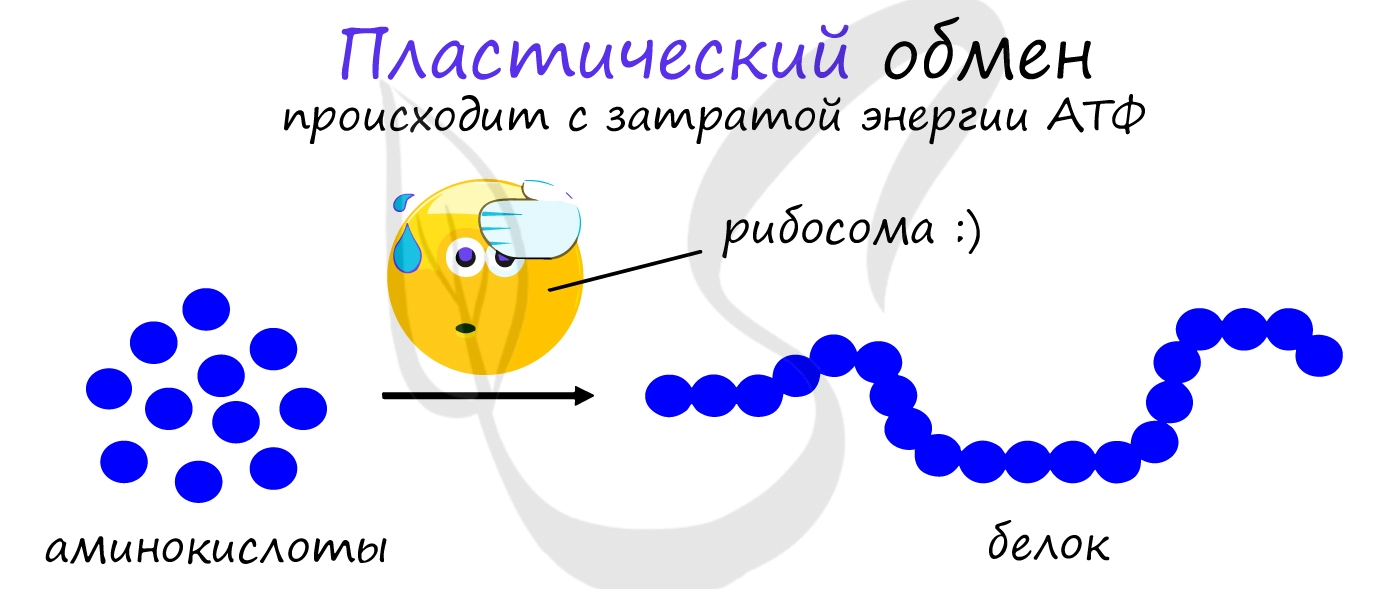
Пластический обмен
АТФ является универсальным источником энергии в клетке: энергия макроэргических связей АТФ используется для реакций
пластического обмена (ассимиляции), протекающих с затратой энергии: синтеза белка на рибосоме (трансляции),
удвоению ДНК (репликации) и т.д.
В результате пластического обмена в нашем организме происходит синтез белков, жиров и углеводов.
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Энергетический обмен
Энергетический обмен
(катаболизм, диссимиляция) — совокупность реакций расщепления органических
веществ, сопровождающихся выделением энергии. Энергия, освобождающаяся при
распаде органических веществ, не сразу используется клеткой, а запасается в
форме АТФ и других высокоэнергетических соединений. АТФ — универсальный
источник энергообеспечения клетки. Синтез АТФ происходит в клетках всех
организмов в процессе фосфорилирования — присоединения неорганического фосфата
к АДФ.
У аэробных
организмов (живущих в кислородной среде) выделяют три этапа энергетического
обмена: подготовительный, бескислородное окисление и кислородное окисление; у анаэробных
организмов (живущих в бескислородной среде) и аэробных при недостатке кислорода
— два этапа: подготовительный, бескислородное окисление.
Подготовительный этап
Заключается в
ферментативном расщеплении сложных органических веществ до простых: белковые
молекулы — до аминокислот, жиры — до глицерина и карбоновых кислот, углеводы —
до глюкозы, нуклеиновые кислоты — до нуклеотидов. Распад высокомолекулярных
органических соединений осуществляется или ферментами желудочно-кишечного
тракта или ферментами лизосом. Вся высвобождающаяся при этом энергия
рассеивается в виде тепла. Образовавшиеся небольшие органические молекулы могут
быть использованы в качестве «строительного материала» или могут подвергаться
дальнейшему расщеплению.
Бескислородное окисление, или гликолиз
Этот этап заключается в
дальнейшем расщеплении органических веществ, образовавшихся во время
подготовительного этапа, происходит в цитоплазме клетки и в присутствии
кислорода не нуждается. Главным источником энергии в клетке является глюкоза.
Процесс бескислородного неполного расщепления глюкозы — гликолиз.
Потеря электронов
называется окислением, приобретение — восстановлением, при этом донор
электронов окисляется, акцептор восстанавливается.
Следует отметить, что
биологическое окисление в клетках может происходить как с участием кислорода:
А + О2
→ АО2,
так и без его участия,
за счет переноса атомов водорода от одного вещества к другому. Например,
вещество «А» окисляется за счет вещества «В»:
АН2
+ В → А + ВН2
или за счет переноса
электронов, например, двухвалентное железо окисляется до трехвалентного:
Fe2+
→ Fe3+ + e—.
Гликолиз — сложный
многоступенчатый процесс, включающий в себя десять реакций. Во время этого
процесса происходит дегидрирование глюкозы, акцептором водорода служит
кофермент НАД+ (никотинамидадениндинуклеотид). Глюкоза в результате
цепочки ферментативных реакций превращается в две молекулы пировиноградной
кислоты (ПВК), при этом суммарно образуются 2 молекулы АТФ и восстановленная
форма переносчика водорода НАД·Н2:
С6Н12О6
+ 2АДФ + 2Н3РО4 + 2НАД+ → 2С3Н4О3
+ 2АТФ + 2Н2О + 2НАД·Н2.
Дальнейшая судьба ПВК
зависит от присутствия кислорода в клетке. Если кислорода нет, у дрожжей и
растений происходит спиртовое брожение, при котором сначала происходит
образование уксусного альдегида, а затем этилового спирта:
1.
С3Н4О3 → СО2 + СН3СОН,
2.
СН3СОН + НАД·Н2 → С2Н5ОН
+ НАД+.
3. У
животных и некоторых бактерий при недостатке кислорода происходит молочнокислое
брожение с образованием молочной кислоты:
4.
С3Н4О3 + НАД·Н2 → С3Н6О3
+ НАД+.
5. В
результате гликолиза одной молекулы глюкозы высвобождается 200 кДж, из которых
120 кДж рассеивается в виде тепла, а 80% запасается в связях АТФ.
6. Кислородное
окисление, или дыхание
7. Заключается
в полном расщеплении пировиноградной кислоты, происходит в митохондриях и при
обязательном присутствии кислорода.
8. Пировиноградная
кислота транспортируется в митохондрии (строение и функции митохондрий ). Здесь
происходит дегидрирование (отщепление водорода) и декарбоксилирование
(отщепление углекислого газа) ПВК с образованием двухуглеродной ацетильной
группы, которая вступает в цикл реакций, получивших название реакций цикла
Кребса. Идет дальнейшее окисление, связанное с дегидрированием и
декарбоксилированием. В результате на каждую разрушенную молекулу ПВК из
митохондрии удаляется три молекулы СО2; образуется пять пар атомов
водорода, связанных с переносчиками (4НАД·Н2, ФАД·Н2), а
также одна молекула АТФ.

Суммарная реакция
гликолиза и разрушения ПВК в митохондриях до водорода и углекислого газа
выглядит следующим образом:
С6Н12О6
+ 6Н2О → 6СО2 + 4АТФ + 12Н2.
Две молекулы АТФ
образуются в результате гликолиза, две — в цикле Кребса; две пары атомов
водорода (2НАДЧН2) образовались в результате гликолиза, десять пар — в цикле
Кребса.
Последним этапом
является окисление пар атомов водорода с участием кислорода до воды с
одновременным фосфорилированием АДФ до АТФ. Водород передается трем большим
ферментным комплексам (флавопротеины, коферменты Q, цитохромы) дыхательной
цепи, расположенным во внутренней мембране митохондрий. У водорода отбираются
электроны, которые в матриксе митохондрий в конечном итоге соединяются с
кислородом:
О2
+ e— → О2—.
Протоны закачиваются в
межмембранное пространство митохондрий, в «протонный резервуар». Внутренняя
мембрана непроницаема для ионов водорода, с одной стороны она заряжается
отрицательно (за счет О2—), с другой — положительно (за
счет Н+). Когда разность потенциалов на внутренней мембране
достигает 200 мВ, протоны проходят через канал фермента АТФ-синтетазы,
образуется АТФ, а цитохромоксидаза катализирует восстановление кислорода до
воды. Так в результате окисления двенадцати пар атомов водорода образуется 34
молекулы АТФ.

1 —
наружная мембрана; 2 — межмембранное пространство, протонный резервуар;
3 — цитохромы; 4 — АТФ-синтетаза.
При перфорации
внутренних митохондриальных мембран окисление НАД·Н2 продолжается,
но АТФ-синтетаза не работает и образования АТФ в дыхательной цепи не
происходит, энергия рассеивается в форме тепла (клетки «бурого жира»
млекопитающих).
Суммарная реакция
расщепления глюкозы до углекислого газа и воды выглядит следующим образом:
С6Н12О6
+ 6О2 → 6СО2 + 6Н2О + 38АТФ + Qт,
где Qт — тепловая
энергия.
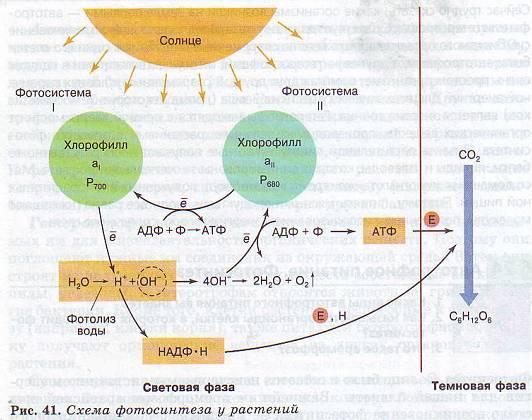
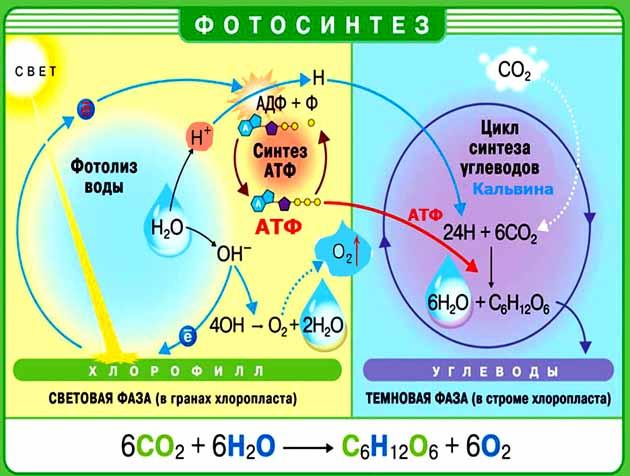
Фотосинтез
— синтез органических веществ из углекислого газа и воды с обязательным
использованием энергии света:
6СО2
+ 6Н2О + Qсвета → С6Н12О6
+ 6О2.
У высших растений
органом фотосинтеза является лист, органоидами фотосинтеза — хлоропласты
(строение хлоропластов — лекция
№7). В мембраны тилакоидов хлоропластов встроены фотосинтетические
пигменты: хлорофиллы и каротиноиды. Существует несколько разных типов
хлорофилла (a, b, c, d), главным является хлорофилл a. В молекуле
хлорофилла можно выделить порфириновую «головку» с атомом магния в центре и
фитольный «хвост». Порфириновая «головка» представляет собой плоскую структуру,
является гидрофильной и поэтому лежит на той поверхности мембраны, которая
обращена к водной среде стромы. Фитольный «хвост» — гидрофобный и за счет этого
удерживает молекулу хлорофилла в мембране.

Хлорофиллы поглощают
красный и сине-фиолетовый свет, отражают зеленый и поэтому придают растениям
характерную зеленую окраску. Молекулы хлорофилла в мембранах тилакоидов
организованы в фотосистемы. У
растений и синезеленых водорослей имеются фотосистема-1 и фотосистема-2, у
фотосинтезирующих бактерий — фотосистема-1. Только фотосистема-2 может
разлагать воду с выделением кислорода и отбирать электроны у водорода воды.
Фотосинтез — сложный
многоступенчатый процесс; реакции фотосинтеза подразделяют на две группы:
реакции световой фазы и реакции темновой
фазы.
Световая фаза
Эта фаза происходит
только в присутствии света в мембранах тилакоидов при участии хлорофилла,
белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта
света электроны хлорофилла возбуждаются, покидают молекулу и попадают на
внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно.
Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды,
находящейся во внутритилакоидном пространстве. Это приводит к распаду или
фотолизу воды:
Н2О
+ Qсвета → Н+ + ОН—.
Ионы гидроксила отдают
свои электроны, превращаясь в реакционноспособные радикалы •ОН:
ОН—
→ •ОН + е—.
Радикалы •ОН
объединяются, образуя воду и свободный кислород:
4НО• →
2Н2О + О2.
Кислород при этом
удаляется во внешнюю среду, а протоны накапливаются внутри тилакоида в
«протонном резервуаре». В результате мембрана тилакоида с одной стороны за счет
Н+ заряжается положительно, с другой за счет электронов —
отрицательно. Когда разность потенциалов между наружной и внутренней сторонами
мембраны тилакоида достигает 200 мВ, протоны проталкиваются через каналы
АТФ-синтетазы и происходит фосфорилирование АДФ до АТФ; атомарный водород идет
на восстановление специфического переносчика НАДФ+
(никотинамидадениндинуклеотидфосфат) до НАДФ·Н2:
2Н+
+ 2е— + НАДФ → НАДФ·Н2.
Таким образом, в
световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими
процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3)
образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2
транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.

1 —
строма хлоропласта; 2 — тилакоид граны.
Темновая фаза
Эта фаза протекает в
строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они
происходят не только на свету, но и в темноте. Реакции темновой фазы
представляют собой цепочку последовательных преобразований углекислого газа
(поступает из воздуха), приводящую к образованию глюкозы и других органических
веществ.
Первая реакция в этой
цепочке — фиксация углекислого газа; акцептором углекислого газа является
пятиуглеродный сахар рибулозобифосфат
(РиБФ); катализирует реакцию фермент рибулозобифосфат-карбоксилаза
(РиБФ-карбоксилаза). В результате карбоксилирования рибулозобисфосфата
образуется неустойчивое шестиуглеродное соединение, которое сразу же
распадается на две молекулы фосфоглицериновой кислоты
(ФГК). Затем происходит цикл реакций, в которых через ряд промежуточных
продуктов фосфоглицериновая кислота преобразуется в глюкозу. В этих реакциях
используются энергии АТФ и НАДФ·Н2, образованных в световую фазу;
цикл этих реакций получил название «цикл Кальвина»:
6СО2
+ 24Н+ + АТФ → С6Н12О6 + 6Н2О.
Кроме глюкозы, в
процессе фотосинтеза образуются другие мономеры сложных органических соединений
— аминокислоты, глицерин и жирные кислоты, нуклеотиды. В настоящее время
различают два типа фотосинтеза: С3— и С4-фотосинтез.
С3-фотосинтез

Это тип фотосинтеза,
при котором первым продуктом являются трехуглеродные (С3)
соединения. С3-фотосинтез был открыт раньше С4-фотосинтеза
(М. Кальвин). Именно С3-фотосинтез описан выше, в рубрике «Темновая
фаза». Характерные особенности С3-фотосинтеза: 1) акцептором
углекислого газа является РиБФ, 2) реакцию карбоксилирования РиБФ катализирует
РиБФ-карбоксилаза, 3) в результате карбоксилирования РиБФ образуется
шестиуглеродное соединение, которое распадается на две ФГК. ФГК
восстанавливается до триозофосфатов
(ТФ). Часть ТФ идет на регенерацию РиБФ, часть превращается в глюкозу.
Фотодыхание

Фотодыхание:
1 — хлоропласт; 2 — пероксисома; 3 — митохондрия.
Это светозависимое
поглощение кислорода и выделение углекислого газа. Еще в начале прошлого века
было установлено, что кислород подавляет фотосинтез. Как оказалось, для
РиБФ-карбоксилазы субстратом может быть не только углекислый газ, но и
кислород:
О2
+ РиБФ → фосфогликолат (2С) + ФГК (3С).
Фермент при этом
называется РиБФ-оксигеназой. Кислород является конкурентным ингибитором
фиксации углекислого газа. Фосфатная группа отщепляется, и фосфогликолат
становится гликолатом, который растение должно утилизировать. Он поступает в
пероксисомы, где окисляется до глицина. Глицин поступает в митохондрии, где
окисляется до серина, при этом происходит потеря уже фиксированного углерода в
виде СО2. В итоге две молекулы гликолата (2С + 2С) превращаются в
одну ФГК (3С) и СО2. Фотодыхание приводит к понижению урожайности С3-растений
на 30–40% (С3-растения — растения, для
которых характерен С3-фотосинтез).
С4-фотосинтез
С4-фотосинтез
— фотосинтез, при котором первым продуктом являются четырехуглеродные (С4)
соединения. В 1965 году было установлено, что у некоторых растений (сахарный
тростник, кукуруза, сорго, просо) первыми продуктами фотосинтеза являются
четырехуглеродные кислоты. Такие растения назвали С4-растениями.
В 1966 году австралийские ученые Хэтч и Слэк показали, что у С4-растений
практически отсутствует фотодыхание и они гораздо эффективнее поглощают углекислый
газ. Путь превращений углерода в С4-растениях стали называть путем
Хэтча-Слэка.
Для С4-растений
характерно особое анатомическое строение листа. Все проводящие пучки окружены
двойным слоем клеток: наружный — клетки мезофилла, внутренний — клетки обкладки.
Углекислый газ фиксируется в цитоплазме клеток мезофилла, акцептор — фосфоенолпируват
(ФЕП, 3С), в результате карбоксилирования ФЕП образуется оксалоацетат (4С).
Процесс катализируется ФЕП-карбоксилазой.
В отличие от РиБФ-карбоксилазы ФЕП-карбоксилаза обладает большим сродством к СО2
и, самое главное, не взаимодействует с О2. В хлоропластах мезофилла
много гран, где активно идут реакции световой фазы. В хлоропластах клеток
обкладки идут реакции темновой фазы.
Оксалоацетат (4С)
превращается в малат, который через плазмодесмы транспортируется в клетки
обкладки. Здесь он декарбоксилируется и дегидрируется с образованием пирувата,
СО2 и НАДФ·Н2.
Пируват возвращается в
клетки мезофилла и регенерирует за счет энергии АТФ в ФЕП. СО2 вновь
фиксируется РиБФ-карбоксилазой с образованием ФГК. Регенерация ФЕП требует
энергии АТФ, поэтому нужно почти вдвое больше энергии, чем при С3-фотосинтезе.
|
|
Строение С4-растений: |
|
|
С4-фотосинтез: |


Значение фотосинтеза
Благодаря фотосинтезу,
ежегодно из атмосферы поглощаются миллиарды тонн углекислого газа, выделяются
миллиарды тонн кислорода; фотосинтез является основным источником образования
органических веществ. Из кислорода образуется озоновый слой, защищающий живые
организмы от коротковолновой ультрафиолетовой радиации.
При фотосинтезе зеленый
лист использует лишь около 1% падающей на него солнечной энергии,
продуктивность составляет около 1 г органического вещества на 1 м2
поверхности в час.
Хемосинтез
Синтез органических
соединений из углекислого газа и воды, осуществляемый не за счет энергии света,
а за счет энергии окисления неорганических веществ, называется хемосинтезом.
К хемосинтезирующим организмам относятся некоторые виды бактерий.
Нитрифицирующие
бактерии окисляют аммиак до азотистой, а затем до азотной кислоты (NH3
→ HNO2 → HNO3).
Железобактерии
превращают закисное железо в окисное (Fe2+ → Fe3+).
Серобактерии
окисляют сероводород до серы или серной кислоты (H2S + ½O2
→ S + H2O, H2S + 2O2 → H2SO4).
В результате реакций
окисления неорганических веществ выделяется энергия, которая запасается
бактериями в форме макроэргических связей АТФ. АТФ используется для синтеза
органических веществ, который проходит аналогично реакциям темновой фазы
фотосинтеза.
Хемосинтезирующие
бактерии способствуют накоплению в почве минеральных веществ, улучшают
плодородие почвы, способствуют очистке сточных вод и др.
Понятие метаболизма
Метаболизм — совокупность всех химических реакций, протекающих в живом организме. Значение метаболизма состоит в создании необходимых организму веществ и обеспечении его энергией.
Выделяют две составные части метаболизма — катаболизм и анаболизм.
Составные части метаболизма
| Часть | Характеристика | Примеры | Затраты энергии |
| Катаболизм (энергетический обмен, диссимиляция) | Совокупность химических реакций, приводящих к образованию простых веществ из более сложных | Гидролиз полимеров до мономеров и расщепление последних до низкомолекулярных соединений углекислого газа, воды, аммиака и других веществ | Энергия выделяется |
| Анаболизм (пластический обмен, ассимиляция) | Совокупность химических реакций синтеза сложных веществ из более простых | Образование углеводов из углекислого газа и воды в процессе фотосинтеза, реакции матричного синтеза | Энергия поглощается |
Процессы пластического и энергетического обмена неразрывно связаны между собой. Все синтетические (анаболические) процессы нуждаются в энергии, поставляемой в ходе реакций диссимиляции. Сами же реакции расщепления (катаболизма) протекают лишь при участии ферментов, синтезируемых в процессе ассимиляции.
Роль ФТФ в метаболизме
Энергия, высвобождающаяся при распаде органических веществ, не сразу используется клеткой, а запасается в форме высокоэнергетических соединений, как правило, в форме аденозинтрифосфата (АТФ). По своей химической природе АТФ относится к мононуклеотидам.
АТФ (аденозинтрифосфорная кислота) — мононуклеотид, состоящий из аденина, рибозы и трёх остатков фосфорной кислоты, соединяющихся между собой макроэргическими связями.
В этих связях запасена энергия, которая высвобождается при их разрыве:
АТФ + H2O → АДФ + H3PO4 + Q1
АДФ + H2O → АМФ + H3PO4 + Q2
АМФ + H2O → аденин + рибоза + H3PO4 + Q3,
где АТФ — аденозинтрифосфорная кислота; АДФ — аденозиндифосфорная кислота; АМФ — аденозинмонофосфорная кислота; Q1 = Q2 = 30,6 кДж; Q3 = 13,8 кДж.
Запас АТФ в клетке ограничен и пополняется благодаря процессу фосфорилирования. Фосфорилирование — присоединение остатка фосфорной кислоты к АДФ (АДФ + Ф → АТФ). Он происходит с разной интенсивностью при дыхании, брожении и фотосинтезе. АТФ обновляется чрезвычайно быстро (у человека продолжительность жизни одной молекулы АТФ менее 1 мин).
Энергия, накопленная в молекулах АТФ, используется организмом в анаболических реакциях (реакциях биосинтеза). Молекула АТФ является универсальным хранителем и переносчиком энергии для всех живых существ.
Энергетический обмен
Энергию, необходимую для жизнедеятельности, большинство организмов получают в результате процессов окисления органических веществ, то есть в результате катаболических реакций. Важнейшим соединением, выступающим в роли топлива, является глюкоза.
По отношению к свободному кислороду организмы делятся на три группы.
Классификация организмов по отношению к свободному кислороду
| Группа | Характеристика | Организмы |
| Аэробы (облигатные аэробы) | Организмы, способные жить только в кислородной среде | Животные, растения, некоторые бактерии и грибы |
| Анаэробы (облигатные анаэробы) | Организмы, неспособные жить в кислородной среде | Некоторые бактерии |
| Факультативные формы (факультативные анаэробы) | Организмы, способные жить как в присутствии кислорода, так и без него | Некоторые бактерии и грибы |
У облигатных аэробов и факультативных анаэробов в присутствии кислорода катаболизм протекает в три этапа: подготовительный, бес- кислородный и кислородный. В результате органические вещества распадаются до неорганических соединений. У облигатных анаэробов и факультативных анаэробов при недостатке кислорода катаболизм протекает в два первых этапа: подготовительный и бескислородный. В результате образуются промежуточные органические соединения, еще богатые энергией.
Этапы катаболизма
1. Первый этап — подготовительный — заключается в ферментативном расщеплении сложных органических соединений на более простые. Белки расщепляются до аминокислот, жиры — до глицерина и жирных кислот, полисахариды — до моносахаридов, нуклеиновые кислоты — до нуклеотидов. У многоклеточных организмов это происходит в желудочно-кишечном тракте, у одноклеточных — в лизосомах под действием гидролитических ферментов. Высвобождающаяся при этом энергия рассеивается в виде теплоты. Образовавшиеся органические соединения либо подвергаются дальнейшему окислению, либо используются клеткой для синтеза собственных органических соединений.
2. Второй этап — неполное окисление (бескислородный) — заключается в дальнейшем расщеплении органических веществ, осуществляется в цитоплазме клетки без участия кислорода. Главным источником энергии в клетке является глюкоза. Бескислородное, неполное окисление глюкозы называется гликолизом. В результате гликолиза одной молекулы глюкозы образуется по две молекулы пировиноградной кислоты (ПВК, пируват) CH3COCOOH, АТФ и воды, а также атомы водорода, которые связываются молекулой-переносчиком НАД+ и запасаются в виде НАД·Н.
Суммарная формула гликолиза имеет следующий вид:
C6H12O6 + 2H3PO4 + 2АДФ + 2НАД+ → 2C3Н4O3 + 2H2O + 2АТФ + 2НАД·Н.
Далее при отсутствии в среде кислорода продукты гликолиза (ПВК и НАД·Н) перерабатываются либо в этиловый спирт — спиртовое брожение (в клетках дрожжей и растений при недостатке кислорода)
CH3COCOOH → СО2 + СН3СОН
СН3СОН + 2НАД·Н → С2Н5ОН + 2НАД+,
либо в молочную кислоту — молочнокислое брожение (в клетках животных при недостатке кислорода)
CH3COCOOH + 2НАД·Н → C3Н6O3 + 2НАД+.
При наличии в среде кислорода продукты гликолиза претерпевают дальнейшее расщепление до конечных продуктов.
3. Третий этап — полное окисление (дыхание) — заключается в окислении ПВК до углекислого газа и воды, осуществляется в митохондриях при обязательном участии кислорода.
Он состоит из трёх стадий:
А) образование ацетилкоэнзима А;
Б) окисление ацетилкоэнзима А в цикле Кребса;
В) окислительное фосфорилирование в электронотранспортной цепи.
А. На первой стадии ПВК переносится из цитоплазмы в митохондрии, где взаимодействует с ферментами матрикса и образует 1) диоксид углерода, который выводится из клетки; 2) атомы водорода, которые молекулами-переносчиками доставляются к внутренней мембране митохондрии; 3) ацетилкофермент А (ацетил-КоА).
Б. На второй стадии происходит окисление ацетилкоэнзима А в цикле Кребса. Цикл Кребса (цикл трикарбоновых кислот, цикл лимонной кислоты) — это цепь последовательных реакций, в ходе которых из одной молекулы ацетил-КоА образуются 1) две молекулы диоксида углерода, 2) молекула АТФ и 3) четыре пары атомов водорода, передаваемые на молекулы-переносчики — НАД и ФАД. Таким образом, в результате гликолиза и цикла Кребса молекула глюкозы расщепляется до СО2, а высвободившаяся при этом энергия расходуется на синтез 4 АТФ и накапливается в 10 НАД·Н и 4 ФАД·Н2.
В. На третьей стадии атомы водорода с НАД·Н и ФАД·Н2 окисляются молекулярным кислородом О2 с образованием воды. Один НАД·Н способен образовывать 3 АТФ, а один ФАД·Н2–2 АТФ. Таким образом, выделяющаяся при этом энергия запасается в виде ещё 34 АТФ.
Этот процесс протекает следующим образом. Атомы водорода концентрируются около наружной стороны внутренней мембраны митохондрии. Они теряют электроны, которые по цепи молекул-переносчиков (цитохромов) электронотранспортной цепи (ЭТЦ) переносятся на внутреннюю сторону внутренней мембраны, где соединяются с молекулами кислорода:
О2 + е— → О2—.
В результате деятельности ферментов цепи переноса электронов внутренняя мембрана митохондрий изнутри заряжается отрицательно (за счёт О2—), а снаружи — положительно (за счёт Н+), так что между её поверхностями создаётся разность потенциалов. Во внутреннюю мембрану митохондрий встроены молекулы фермента АТФ- синтетазы, обладающие ионным каналом. Когда разность потенциалов на мембране достигает критического уровня, положительно заряженные частицы H+ силой электрического поля начинают проталкиваться через канал АТФазы и, оказавшись на внутренней поверхности мембраны, взаимодействуют с кислородом, образуя воду:
1/2О2— +2H+ → Н2О.
Энергия ионов водорода H+, транспортирующихся через ионный канал внутренней мембраны митохондрии, используется для фосфорилирования АДФ в АТФ:
АДФ + Ф → АТФ.
Такое образование АТФ в митохондриях при участии кислорода называется окислительным фосфорилированием.
Суммарное уравнение расщепления глюкозы в процессе клеточного дыхания:
C6H12O6 + 6O2 + 38H3PO4 + 38АДФ → 6CO2 + 44H2O + 38АТФ.
Таким образом, в ходе гликолиза образуются 2 молекулы АТФ, в ходе клеточного дыхания — ещё 36 молекул АТФ, в целом при пол- ном окислении глюкозы — 38 молекул АТФ.
Пластический обмен
Пластический обмен, или ассимиляция, представляет собой совокупность реакций, обеспечивающих синтез сложных органических соединений из более простых (фотосинтез, хемосинтез, биосинтез белка и др.).
Гетеротрофные организмы строят собственные органические вещества из органических компонентов пищи. Гетеротрофная ассимиляция сводится, по существу, к перестройке молекул:
органические вещества пищи (белки, жиры, углеводы) → простые органические молекулы (аминокислоты, жирные кислоты, моносахариды) → макромолекулы тела (белки, жиры, углеводы).
Автотрофные организмы способны полностью самостоятельно синтезировать органические вещества из неорганических молекул, потребляемых из внешней среды. В процессе фото- и хемосинтеза происходит образование простых органических соединений, из которых в дальнейшем синтезируются макромолекулы:
неорганические вещества (СО2, Н2О) → простые органические молекулы (аминокислоты, жирные кислоты, моносахариды) → макромолекулы тела (белки, жиры, углеводы).
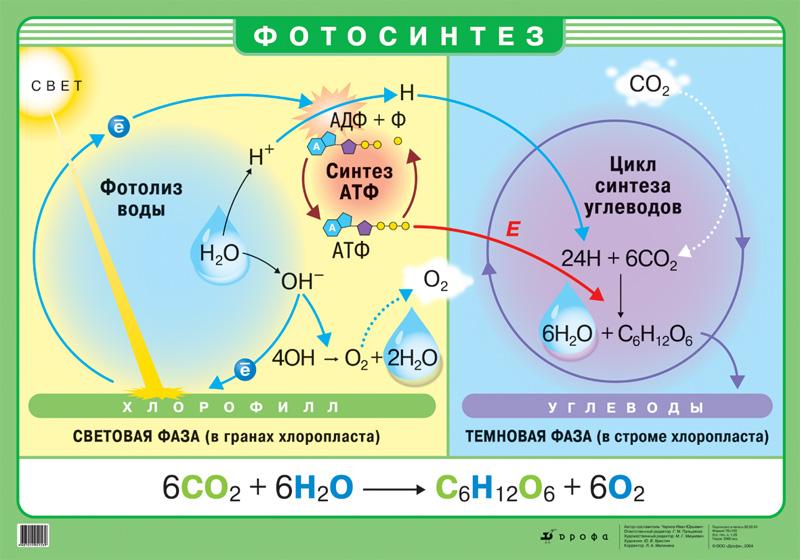
Фотосинтез
Фотосинтез — синтез органических соединений из неорганических за счёт энергии света. Суммарное уравнение фотосинтеза:
Фотосинтез протекает при участии фотосинтезирующих пигментов, обладающих уникальным свойством преобразования энергии солнечного света в энергию химической связи в виде АТФ. Фотосинтезирующие пигменты представляют собой белковоподобные вещества. Наиболее важным является пигмент хлорофилл. У эукариот фотосинтезирующие пигменты встроены во внутреннюю мембрану пластид, у прокариот — во впячивания цитоплазматической мембраны.
Строение хлоропласта очень похоже на строение митохондрии. Во внутренней мембране тилакоидов гран содержатся фотосинтетические пигменты, а также белки цепи переноса электронов и молекулы фермента АТФ-синтетазы.
Процесс фотосинтеза состоит из двух фаз: световой и темновой.
1. Световая фаза фотосинтеза протекает только на свету в мембране тилакоидов граны.
К ней относятся поглощение хлорофиллом квантов света, образование молекулы АТФ и фотолиз воды.
Под действием кванта света (hv) хлорофилл теряет электроны, переходя в возбуждённое состояние:
Эти электроны передаются переносчиками на наружную, то есть обращенную к матриксу поверхность мембраны тилакоидов, где накапливаются.
Одновременно внутри тилакоидов происходит фотолиз воды, то есть её разложение под действием света:
Образующиеся электроны передаются переносчиками к молекулам хлорофилла и восстанавливают их. Молекулы хлорофилла возвращаются в стабильное состояние.
Протоны водорода, образовавшиеся при фотолизе воды, накапливаются внутри тилакоида, создавая Н+-резервуар. В результате внутренняя поверхность мембраны тилакоида заряжается положительно (за счёт Н+), а наружная — отрицательно (за счёт е—). По мере накопления по обе стороны мембраны противоположно заряженных частиц нарастает разность потенциалов. При достижении критической величины разности потенциалов сила электрического поля начинает проталкивать протоны через канал АТФ-синтетазы. Выделяющаяся при этом энергия используется для фосфорилирования молекул АДФ:
АДФ + Ф → АТФ.
Образование АТФ в процессе фотосинтеза под действием энергии света называется фотофосфорилированием.
Ионы водорода, оказавшись на наружной поверхности мембраны тилакоида, встречаются там с электронами и образуют атомарный водород, который связывается с молекулой-переносчиком водорода НАДФ (никотинамидадениндинуклеотидфосфат):
2Н+ + 4е– + НАДФ+ → НАДФ·Н2.
Таким образом, во время световой фазы фотосинтеза происходят три процесса: образование кислорода вследствие разложения воды, синтез АТФ и образование атомов водорода в форме НАДФ·Н2. Кислород диффундирует в атмосферу, а АТФ и НАДФ·Н2 участвуют в процессах темновой фазы.
2. Темновая фаза фотосинтеза протекает в матриксе хлоропласта как на свету, так и в темноте и представляет собой ряд последовательных преобразований СО2, поступающего из воздуха, в цикле Кальвина. Осуществляются реакции темновой фазы за счёт энергии АТФ. В цикле Кальвина СО2 связывается с водородом из НАДФ·Н2 с образованием глюкозы.
В процессе фотосинтеза кроме моносахаридов (глюкоза и др.) синтезируются мономеры других органических соединений — аминокислоты, глицерин и жирные кислоты. Таким образом, благодаря фотосинтезу растения обеспечивают себя и всё живое на Земле необходимыми органическими веществами и кислородом.
Сравнительная характеристика фотосинтеза и дыхания эукариот представлена в таблице.
Сравнительная характеристика фотосинтеза и дыхания эукариот
| Признак | Фотосинтез | Дыхание |
| Уравнение реакции | 6СО2 + 6Н2О + энергия света → C6H12O6 + 6O2 | C6H12O6 + 6O2 → 6СО2 + 6Н2О + энергия (АТФ) |
| Исходные вещества | Углекислый газ, вода | Органические вещества, кислород |
| Продукты реакции | Органические вещества, кислород | Углекислый газ, вода |
| Значение в круговороте веществ | Синтез органических веществ из неорганических | Разложение органических веществ до неорганических |
| Превращение энергии | Превращение энергии света в энергию химических связей органических веществ | Превращение энергии химических связей органических веществ в энергию макроэргических связей АТФ |
| Важнейшие этапы | Световая и темновая фаза (включая цикл Кальвина) | Неполное окисление (гликолиз) и полное окисление (включая цикл Кребса) |
| Место протекания процесса | Хлоропласты | Гиалоплазма (неполное окисление) и митохондрии (полное окисление) |
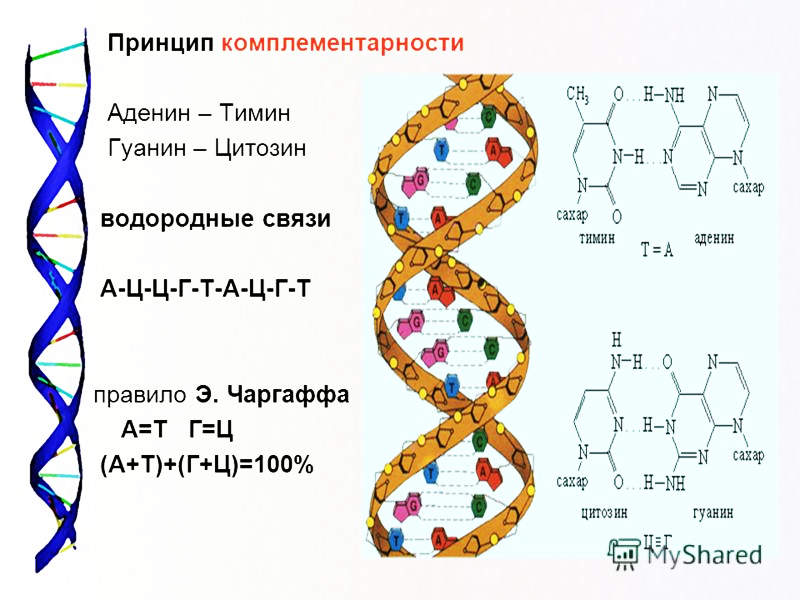
Генетическая информация у всех организмов хранится в виде определённой последовательности нуклеотидов ДНК (или РНК у РНК-содержащих вирусов). Прокариоты содержат генетическую информацию в виде одной молекулы ДНК. В эукариотических клетках генетический материал распределён в нескольких молекулах ДНК, организованных в хромосомы.
ДНК состоит из кодирующих и некодирующих участков. Кодирующие участки кодируют РНК. Некодирующие области ДНК выполняют структурную функцию, позволяя участкам генетического материала упаковываться определённым образом, или регуляторную функцию, участвуя во включении генов, направляющих синтез белка.
Кодирующими участками ДНК являются гены. Ген — участок молекулы ДНК, кодирующей синтез одной мРНК (и соответственно полипептида), рРНК или тРНК.
Участок хромосомы, где расположен ген называется локусом. Совокупность генов клеточного ядра представляет собой генотип, совокупность генов гаплоидного набора хромосом — гено́м, совокупность генов внеядерных ДНК (митохондрий, пластид, цитоплазмы) — плазмон.
Реализация информации, записанной в генах, через синтез белков называется экспрессией (проявлением) генов. Генетическая информация хранится в виде определённой последовательности нуклеотидов ДНК, а реализуется в виде последовательности аминокислот в белке. Посредниками, переносчиками информации выступают РНК. То есть реализация генетической информации происходит следующим образом:
ДНК → РНК → белок.
Этот процесс осуществляется в два этапа:
1) транскрипция;
2) трансляция.
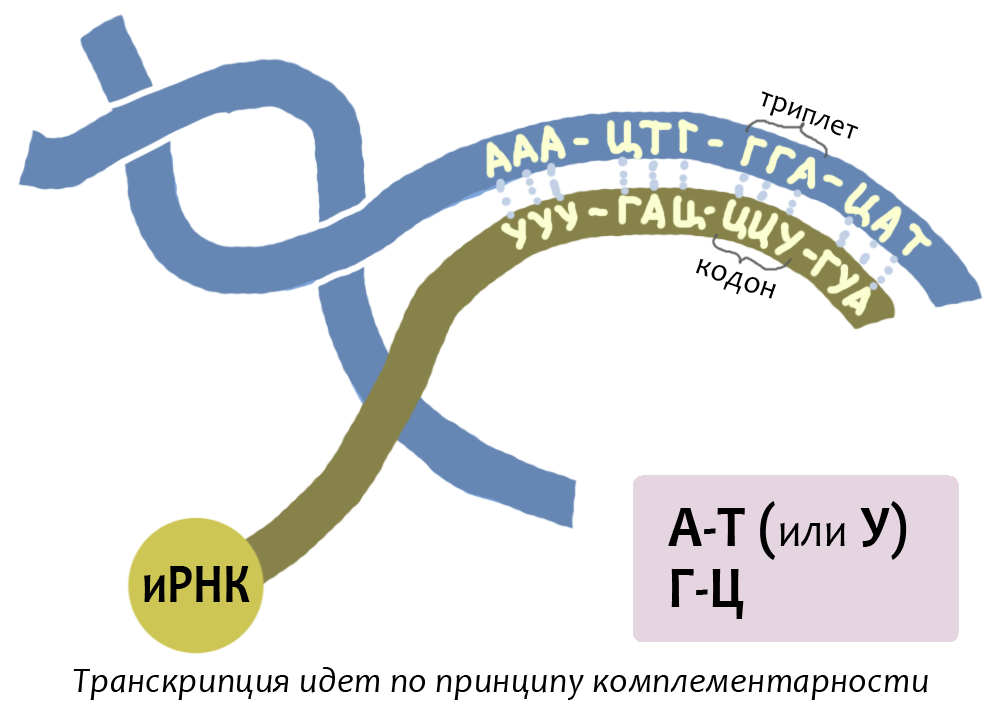
Транскрипция (от лат. transcriptio — переписывание) — синтез РНК с использованием ДНК в качестве матрицы. В результате образуются мРНК, тРНК и рРНК. Процесс транскрипции требует больших затрат энергии в виде АТФ и осуществляется ферментом РНК-полимеразой.
Одновременно транскрибируется не вся молекула ДНК, а лишь отдельные её отрезки. Такой отрезок (транскриптон) начинается промотором — участком ДНК, куда присоединяется РНК-полимераза и откуда начинается транскрипция, а заканчивается терминатором — участком ДНК, содержащим сигнал окончания транскрипции. Транскриптон — это ген с точки зрения молекулярной биологии.
Транскрипция, как и репликация, основана на способности азотистых оснований нуклеотидов к комплементарному связыванию. На время транскрипции двойная цепь ДНК разрывается, и синтез РНК осуществляется по одной цепи ДНК.
В процессе транскрипции последовательность нуклеотидов ДНК переписывается на синтезирующуюся молекулу мРНК, которая выступает в качестве матрицы в процессе биосинтеза белка.
Гены прокариот состоят только из кодирующих нуклеотидных последовательностей.
Гены эукариот состоят из чередующихся кодирующих (экзонов) и некодирующих (интронов) участков.
После транскрипции участки мРНК, соответствующие интронам, удаляются в ходе сплайсинга, являющегося составной частью процессинга.
Процессинг — процесс формирования зрелой мРНК из её предшественника пре-мРНК. Он включает два основных события. 1.Присоединение к концам мРНК коротких последовательностей нуклеотидов, обозначающих место начала и место конца трансляции. Сплайсинг — удаление неинформативных последовательностей мРНК, соответствующих интронам ДНК. В результате сплайсинга молекулярная масса мРНК уменьшается в 10 раз.
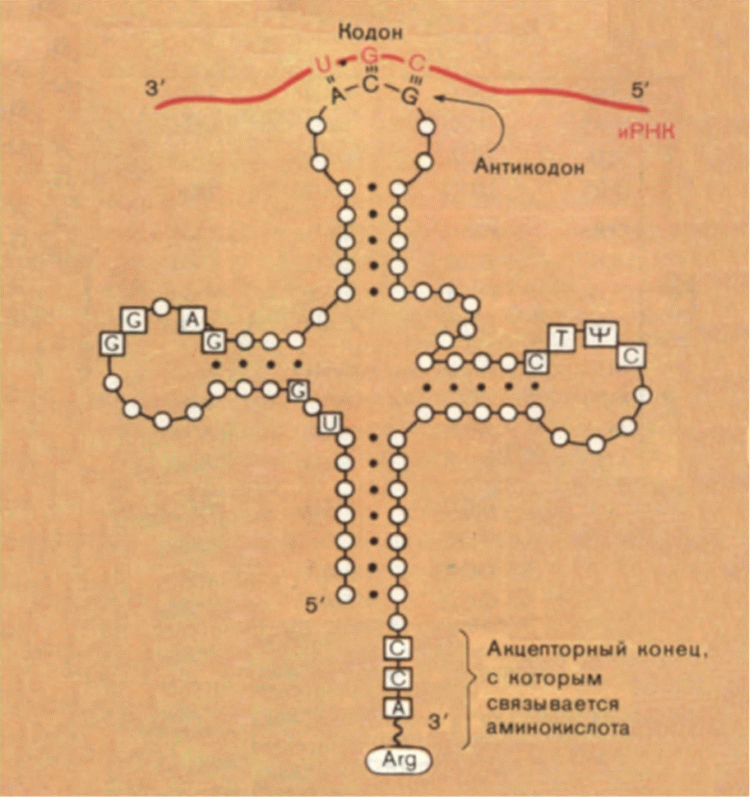
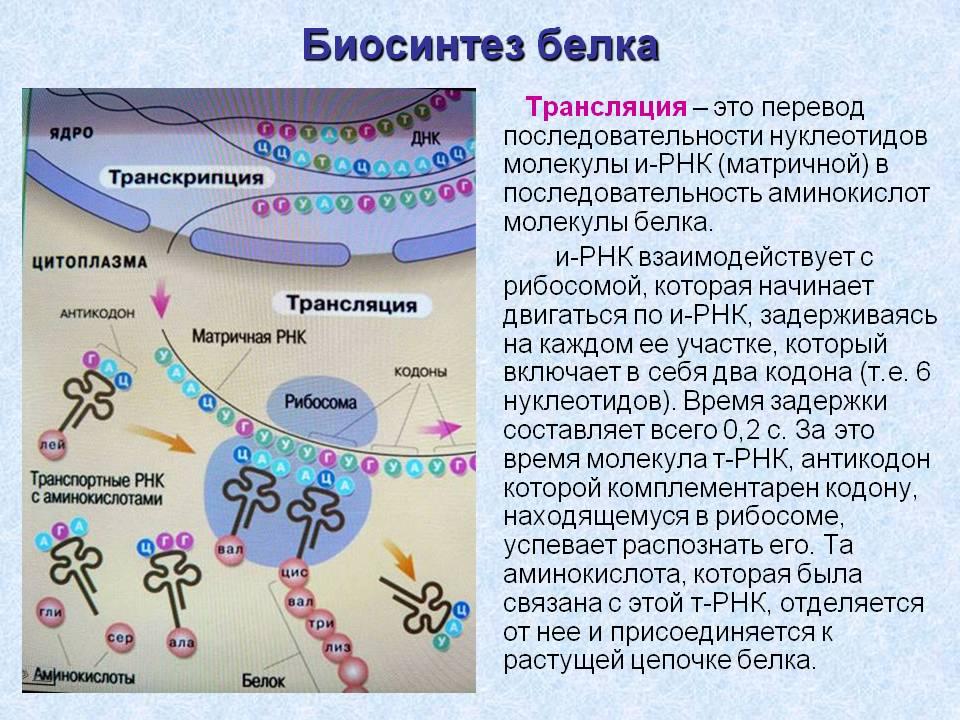
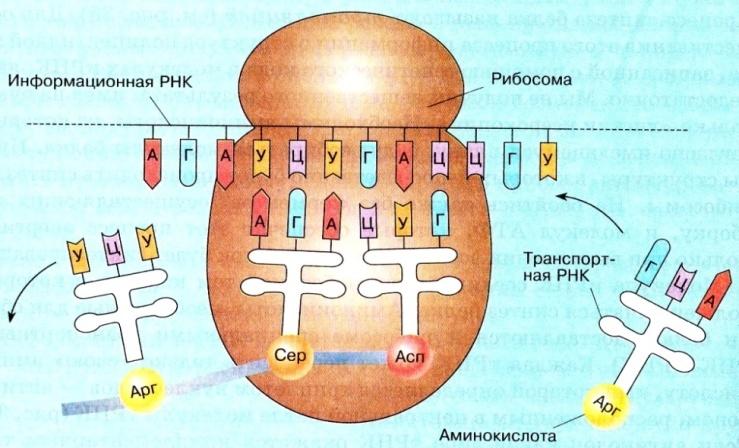
Трансляция (от лат. translatio — перевод) — синтез полипептидной цепи с использованием мРНК в роли матрицы.
В трансляции участвуют все три типа РНК: мРНК является информационной матрицей; тРНК доставляют аминокислоты и узнают кодоны; рРНК вместе с белками образуют рибосомы, которые удерживают мРНК, тРНК и белок и осуществляют синтез полипептидной цепи.
Этапы трансляции
| Этап | Характеристика |
| Инициация | Сборка комплекса, участвующего в синтезе полипептидной цепи. Малая субчастица рибосомы соединяется с инициаторной мет-трнк, а затем с мрнк, после чего происходит образование целой рибосомы, состоящей из малой и большой субчастиц. |
| Элонгация | Удлинение полипептидной цепи. Рибосома перемещается вдоль мрнк, что сопровождается многократным повторением цикла присоединения очередной аминокислоты к растущей полипептидной цепи. |
| Терминация | Завершение синтеза полипептидной молекулы. Рибосома достигает одного из трёх стоп-кодонов мрнк, а так как не существует трнк с антикодонами, комплементарными стоп-кодонам, синтез полипептидной цепи прекращается. Она высвобождается и отделяется от рибосомы. Рибосомные субчастицы диссоциируют, отделяются от мрнк и могут принять участие в синтезе следующей полипептидной цепи. |
Реакции матричного синтеза. К реакциям матричного синтеза относятся
- самоудвоение ДНК (репликация);
- образование мРНК, тРНК и рРНК на молекуле ДНК (транскрипция);
- биосинтез белка на мРНК (трансляция).
Все эти реакции объединяет то, что молекула ДНК в одном случае или молекула мРНК в другом выступают в роли матрицы, на которой происходит образование одинаковых молекул. Реакции матричного синтеза являются основой способности живых организмов к воспроизведению себе подобных.
Регуляция экспрессии генов. Тело многоклеточного организма построено из разнообразных клеточных типов. Они отличаются структурой и функциями, то есть дифференцированы. Различия проявляются в том, что помимо белков, необходимых любой клетке организма, клетки каждого типа синтезируют ещё и специализированные белки: в эпидермисе образуется кератин, в эритроцитах — гемоглобин и т. д. Клеточная дифференцировка обусловлена изменением набора экспрессируемых генов и не сопровождается какими-либо необратимыми изменениями в структуре самих последовательностей ДНК.
«Обмен веществ и превращения энергии. Ферменты»
Раздел ЕГЭ: 2.5. Обмен веществ и превращения энергии — свойства живых организмов. Энергетический обмен и пластический обмен, их взаимосвязь. Стадии энергетического обмена. Брожение и дыхание. Фотосинтез, его значение, космическая роль. Фазы фотосинтеза. Световые и темновые реакции фотосинтеза, их взаимосвязь. Хемосинтез. Роль хемосинтезирующих бактерий на Земле.
Клетку можно уподобить миниатюрной химической фабрике, на которой происходят сотни и тысячи химических реакций. Обмен веществ — совокупность химических превращений, направленных на сохранение и самовоспроизведение биологических систем. Он включает в себя:
- поступление веществ в организм в процессе питания и дыхания,
- внутриклеточный обмен веществ, или метаболизм,
- выделение конечных продуктов обмена.
Метаболизм складывается из двух одновременно протекающих в клетке процессов: пластического и энергетического обменов.
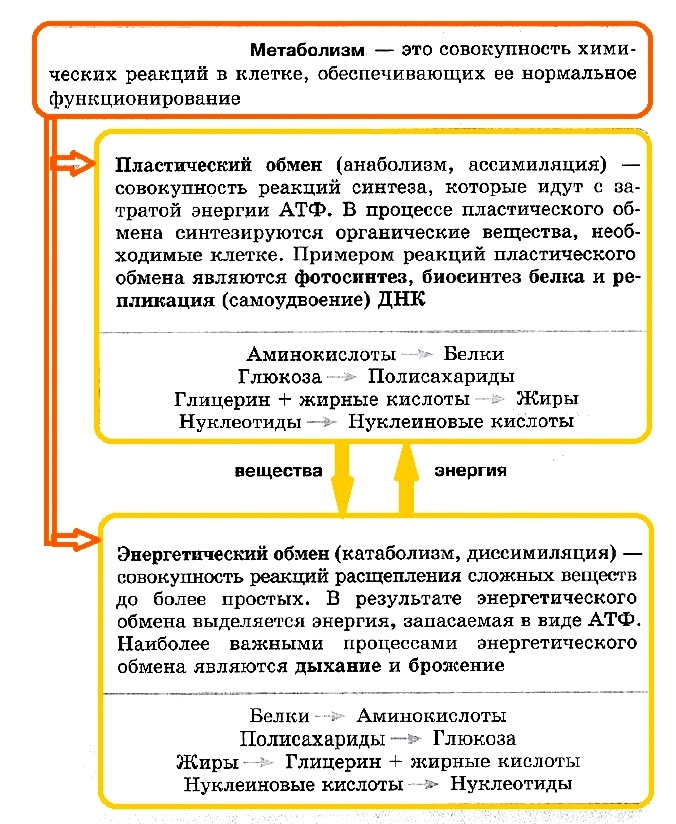
Энергетический обмен и пластический обмен
Пластический обмен (анаболизм, ассимиляция) представляет собой совокупность реакций синтеза, которые идут с затратой энергии АТФ. В процессе пластического обмена синтезируются органические вещества, необходимые клетке. Примерами реакций пластического обмена являются фотосинтез, биосинтез белка и репликация (самоудвоение) ДНК.
Энергетический обмен (катаболизм, диссимиляция) — это совокупность реакций расщепления сложных веществ до более простых. В результате энергетического обмена выделяется энергия, запасаемая в виде АТФ. Наиболее важными процессами энергетического обмена являются дыхание и брожение.
Пластический и энергетический обмены неразрывно связаны, поскольку в процессе пластического обмена синтезируются органические вещества и для этого необходима энергия АТФ, а в процессе энергетического обмена органические вещества расщепляются и высвобождается энергия, которая затем будет израсходована на процессы синтеза.
Энергию организмы получают в процессе питания, а высвобождают ее и переводят в доступную форму в основном в процессе дыхания.
Схема общего обмена веществ
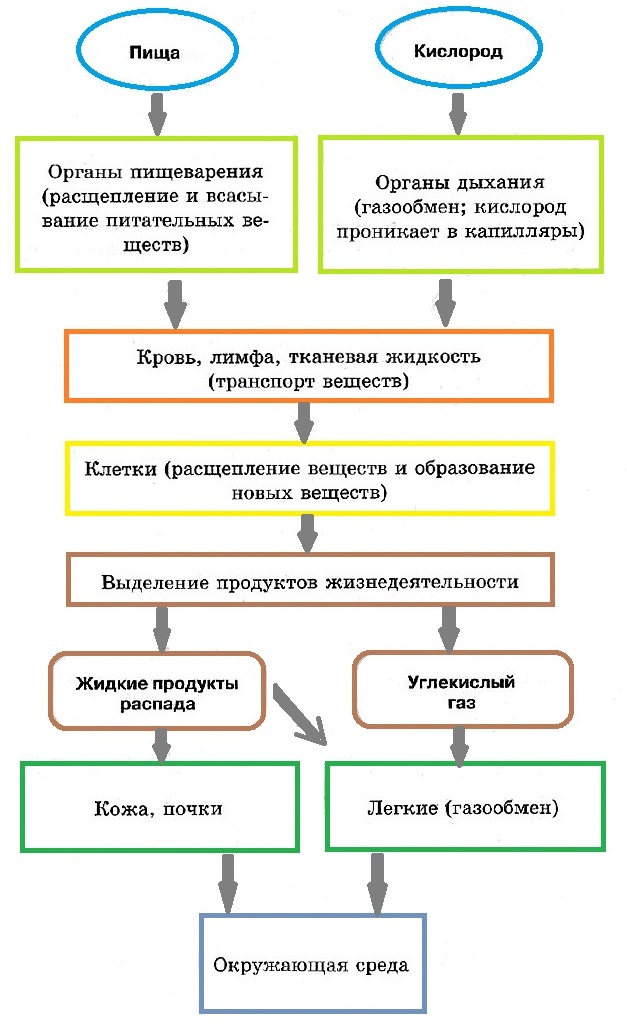
Ферменты
Протекание химических реакций в живых организмах обеспечивается благодаря биологическим катализаторам белковой природы — ферментам, или энзимам. Как и другие катализаторы, ферменты ускоряют протекание химических реакций в клетке в десятки и сотни тысяч раз, а иногда и вообще делают их возможными, но не изменяют при этом ни природы, ни свойств конечного продукта (продуктов) реакции и не изменяются сами. Ферменты могут быть как простыми, так и сложными белками, в состав которых, кроме белковой части, входит и небелковая — кофактор (кофермент). Примерами ферментов являются амилаза слюны, расщепляющая полисахариды при длительном пережевывании, и пепсин, обеспечивающий переваривание белков в желудке.
Ферменты отличаются от катализаторов небелковой природы высокой специфичностью действия, а также возможностью регуляции действия за счет изменения условий протекания реакции либо взаимодействия с ними различных веществ. К тому же и условия, в которых протекает ферментный катализ, существенно отличаются от тех, при которых идет неферментный: оптимальной для функционирования ферментов в организме человека является температура 37° С, а давление должно быть близким к атмосферному.
Механизм действия ферментов заключается в снижении энергии активации веществ (субстратов), вступающих в реакцию, за счет образования промежуточных фермент-субстратных комплексов.
Это конспект для 10-11 классов по теме «Обмен веществ и превращения энергии. Ферменты».
Читайте также другие конспекты, относящиеся к разделу ЕГЭ 2.5:
- Стадии энергетического обмена. Брожение и дыхание.
- Фотосинтез, его значение, космическая роль. Фазы фотосинтеза. Световые и темновые реакции фотосинтеза, их взаимосвязь. Хемосинтез. Роль хемосинтезирующих бактерий на Земле
Энергетический обмен.
(катаболизм (в переводе — разрушение) , диссимиляция)
Автор статьи — Л.В. Окольнова.
Энергетический обмен — это часть процесса обмена веществ (метаболизма).
Этими терминами называют:
— распад сложного вещества (полимера) на более простые (мономеры);
— окисление веществ;
— превращение органических веществ в неорганические;
Обязательное условие — выделение тепла и энергии (АТФ)
Самые часто встречающиеся катаболические процессы в организмах:
— пищеварение;
— дыхание;
— разложение редуцентами органических веществ до неорганических;
— брожение.
Все живые организмы в природе по типу дыхания делятся на 2 группы:
|
Аэробы (+О2) |
Анаэробы (-О2) |
|
используют 02 для дыхания и обмена веществ |
живут в бескислородной среде |
|
большинство животных |
бактерии (кроме фотосинтезирующих) |
|
растения |
грибы |
|
некоторые микроорганизмы |
паразитические животные |
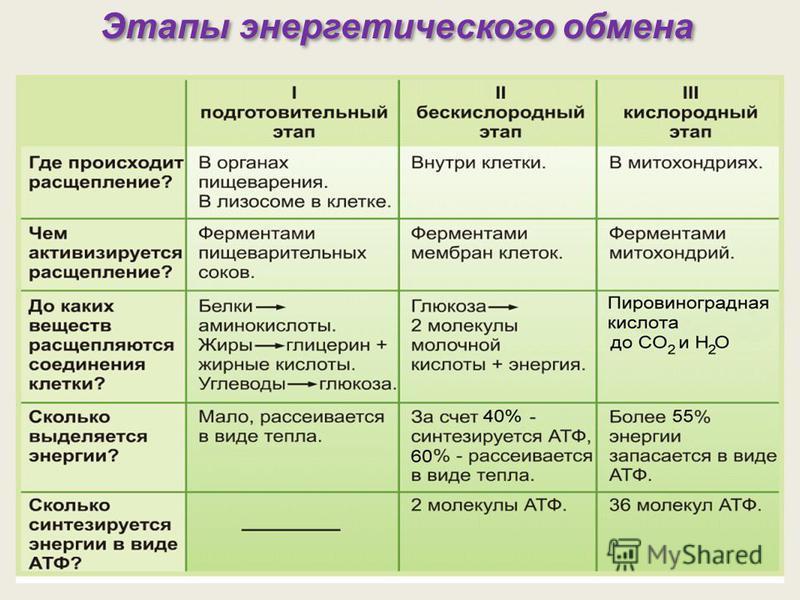
Стадии энергетического обмена аэробов:
3 этапа энергетического обмена:
— подготовительный;
— бескислородный;
— кислородный.
Для анаэробов:
2 этапа энергетического обмена:
— подготовительный;
— бескислородный.
Рассмотрим аэробный энергетический обмен:
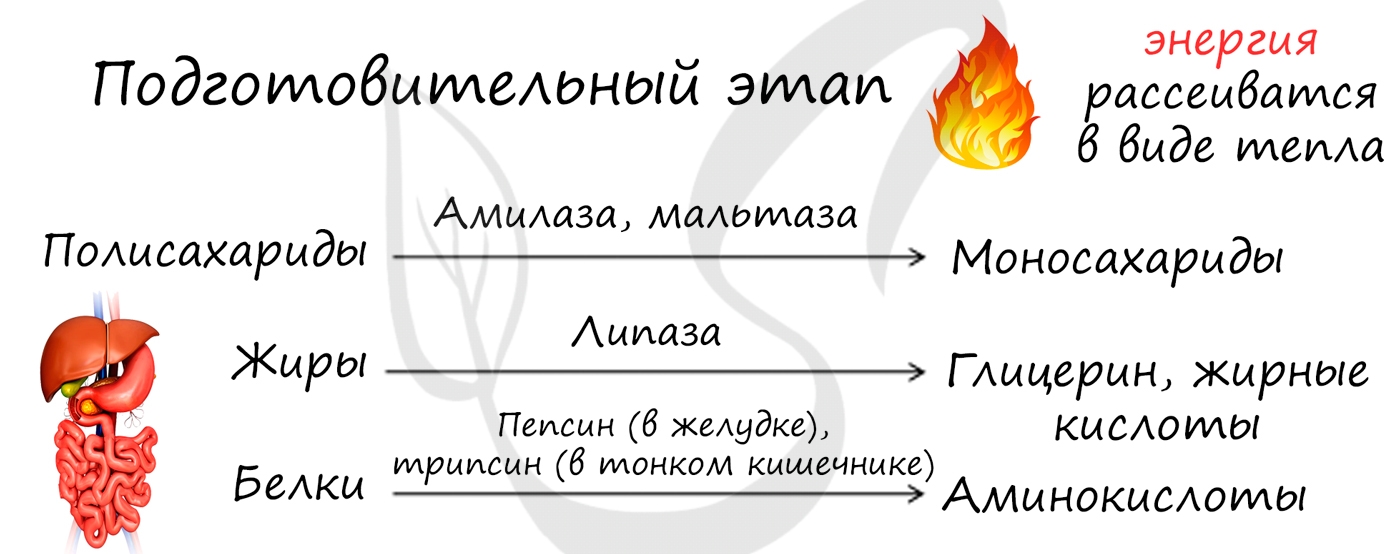
1 этап —подготовительный.
Все живые существа потребляют пищу органические вещества в виде крупных молекул — полимеров.
Первое, что необходимо для пищеварения — расщепить эти полимеры на более простые и небольшие составляющие — мономеры.
Расщепляются (диссимилируют) вещества под действием ферментов и в определенной среде. Причем, для каждого вещества существует свой фермент (это называется специфичностью ферментов).
У многоклеточных организмов это происходит в пищеварительной системе, у одноклеточных — прямо в клетке в лизосомах.

У многоклеточных организмов мономеры всасываются в кровь, разносятся кровью к тканям и органам и поступают в клетки для следующего этапа.
У одноклеточных — идут в запас в аппарат Гольджи, в рибосомы — для синтеза новых белков и глюкоза — в цитоплазму для следующего этапа.
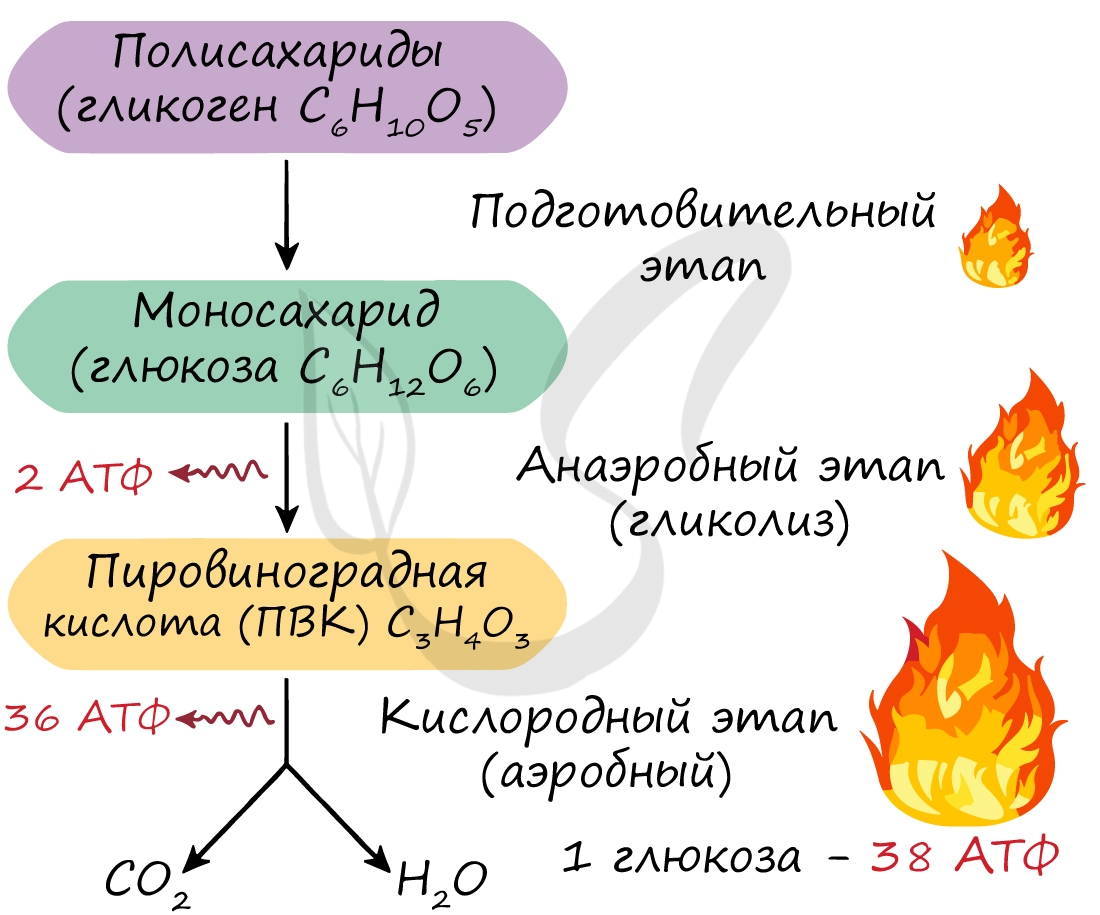
2 этап — в цитоплазме клеток — бескислородный.
(его рассматривают только на примере углеводов).

3 этап — в митохондриях — кислородный.
Этот процесс сложный, многостадийный, обязательно участвуют ферменты, мы его рассмотрим схематично.

Все процессы суммарно:
| подготовительный | расщепление полимеров | в пищеварительной системе у многоклеточных,
в лизосомах у одноклеточных |
крахмал -> глюкоза, выделяется тепло |
| бескислородный | расщепление глюкозы | в цитоплазме | глюкоза -> пировиноградная кислота + 2АТФ |
| кислородный | расщепление пировиноградной кислоты | в митохондриях | пировиноградная кислота -> CO2 + H2O + 36 АТФ |

Спасибо за то, что пользуйтесь нашими материалами.
Информация на странице «Энергетический обмен.» подготовлена нашими авторами специально, чтобы помочь вам в освоении предмета и подготовке к ЕГЭ и ОГЭ.
Чтобы успешно сдать необходимые и поступить в высшее учебное заведение или техникум нужно использовать все инструменты: учеба, контрольные, олимпиады, онлайн-лекции, видеоуроки, сборники заданий.
Также вы можете воспользоваться другими статьями из разделов нашего сайта.
Публикация обновлена:
09.03.2023

Видео YouTube

СТАДИИ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
Энергетический обмен (катаболизм, диссимиляция) — это процессы расщепления ве ществ с высвобождением энергии. Высвобожденная энергия преобразуется в энергию АТФ. Наиболее важными процессами энергетического обмена являются дыхание и брожение.

Энергетический обмен – это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ. Синтезированная АТФ становится универсальным источником энергии для жизнедеятельности организмов. Она образуется в результате реакции фосфорилирования – присоединения остатков фосфорной кислоты к молекуле АДФ. На эту реакцию расходуется энергия, которая затем накапливается в макроэргических связях молекулы АТФ, при распаде молекулы АТФ или при ее гидролизе до АДФ клетка получает около 40 кДж энергии.
АТФ – постоянный источник энергии для клетки, она мобильно может доставлять химическую энергию в любую часть клетки. Когда клетке необходима энергия – достаточно гидролизовать молекулу АТФ. Энергия выделяется в результате реакции диссимиляции (расщепления органических веществ), в зависимости от специфики организма и условий его обитания энергетический обмен проходит в два или три этапа. Большинство живых организмов относятся к аэробам, использующим для обмена веществ кислород, который поступает из окружающей среды. Для аэробов энергетический обмен проходит в три этапа:
— подготовительный;
— бескислородный;
— кислородный.
В организмах, которые обитают в бескислородной среде и не нуждаются в кислороде для энергетического обмена – анаэробах и аэробах, при недостатке кислорода проходят энергетический обмен в два этапа:
— подготовительный;
— бескислородный.
Количество энергии, которое выделяется при двухэтапном варианте намного меньше, чем в трехэтапном.
ЭТАПЫ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
Подготовительный этап – во время него крупные пищевые полимерные молекулы распадаются на более мелкие фрагменты. В желудочно-кишечном тракте многоклеточных организмов он осуществляется пищеварительными ферментами, у одноклеточных – ферментами лизосом. Полисахариды распадаются на ди- и моносахариды, белки – до аминокислот, жиры – до глицерина и жирных кислот. В ходе этих превращений энергии выделяется мало, она рассеивается в виде тепла, и АТФ не образуется. Образующиеся в ходе подготовительного этапа соединения-мономеры могут участвовать в реакциях пластического обмена (в дальнейшем из них синтезируются вещества, необходимые для клетки) или подвергаться дальнейшему расщеплению с целью получения энергии.
Большинство клеток в первую очередь используют углеводы, жиры остаются в первом резерве и используются по окончания запаса углеводов. Хотя есть и исключения: в клетках скелетных мышц при наличии жирных кислот и глюкозы предпочтение отдается жирным кислотам. Белки расходуются в последнюю очередь, когда запас углеводов и жиров будет исчерпан – при длительном голодании.
Бескислородный этап (гликолиз) – происходит в цитоплазме клеток. Главным источником энергии в клетке является глюкоза. Ее бескислородное расщепление называют анаэробным гликолизом. Он состоит из ряда последовательных реакций по превращению глюкозы в лактат. Его присутствие в мышцах хорошо известно уставшим спортсменам. Этот этап заключается в ферментативном расщеплении органических веществ, полученных в ходе первого этапа. Так как глюкоза является наиболее доступным субстратом для клетки как продукт расщепления полисахаридов, то второй этап можно рассмотреть на примере ее бескислородного расщепления – гликолиза (Рис. 1).

Рис. 1. Бескислородный этап
Гликолиз – многоступенчатый процесс бескислородного расщепления молекулы глюкозы, содержащей шесть атомов углерода, до двух молекул пировиноградной кислоты (пируват). Реакция гликолиза катализируется многими ферментами и протекает в цитоплазме клетки. В ходе гликолиза при расщеплении одного моля глюкозы выделяется около 200 кДж энергии, 60 % ее рассеивается в виде тепла, 40 % – для синтезирования двух молекул АТФ из двух молекул АДФ. При наличии кислорода в среде пировиноградная кислота из цитоплазмы переходит в митохондрии и участвует в третьем этапе энергетического обмена. Если кислорода в клетке нет, то пировиноградная кислота преобразуется в животных клетках или превращается в молочную кислоту.
В микроорганизмах, которые существуют без доступа кислорода – получают энергию в процессе брожения, начальный этап аналогичен гликолизу: распад глюкозы до двух молекул пировиноградной кислоты, и далее она зависит от ферментов, которые находятся в клетке – пировиноградная кислота может преобразовываться в спирт, уксусную кислоту, пропионовую и молочную кислоту. В отличие от того, что происходит в животных тканях, у микроорганизмов этот процесс носит название молочнокислого брожения. Все продукты брожения широко используются в практической деятельности человека: это вино, квас, пиво, спирт, кисломолочные продукты. При брожении, так же как и при гликолизе, выделяется всего две молекулы АТФ.
Кислородный этап стал возможен после накопления в атмосфере достаточного количества молекулярного кислорода, он происходит в митохондриях клеток. Он очень сложен по сравнению с гликолизом, это процесс многостадийный и идет при участии большого количества ферментов. В результате третьего этапа энергетического обмена из двух молекул пировиноградной кислоты формируется углекислый газ, вода и 36 молекул АТФ (Рис. 2).

Рис. 2. Митохондрия
Две молекулы АТФ запасаются в ходе бескислородного расщепления молекулами глюкозы, поэтому суммарный энергетический обмен в клетке в случае распада глюкозы можно представить как:
С6Н12О6 + 6О2 + 38АДФ + 38Н3РО4 = 6СО2 + 44Н2О + 38АТФ
В результате окисления одной молекулы глюкозы шестью молекулами кислорода образуется шесть молекул углекислого газа и выделяется тридцать восемь молекул АТФ.
Мы видим, что в трехэтапном варианте энергетического обмена выделяется гораздо больше энергии, чем в двухэтапном варианте – 38 молекул АТФ против 2.


БРОЖЕНИЕ

В отсутствие кислорода или при его недостатке про исходит брожение. Брожение является эволюционно бо лее ранним способом получения энергии, чем дыхание, однако оно энергетически менее выгодно, поскольку ко нечными продуктами брожения являются органические вещества, богатые энергией. Существует несколько видов брожения, названия которых определяются конечными продуктами: молочнокислое, спиртовое, уксуснокислое и др. Так, в скелетных мышцах в отсутствие кислорода протекает молочнокислое брожение, в ходе которого пировиноградная кислота восстанавли вается до молочной кислоты. При этом восстановленные ранее коферменты НАДН расходу ются на восстановление пирувата:

Энергетическая эффективность молочнокислого брожения составляет две молекулыАТФ, образованные в процессе окисления глюкозы до пирувата.
Для многих микроорганизмов брожение является единственным способом ассимиляции энергии. Большинство таких организмов живет в анаэробных условиях и погибает в присутствии кислорода, но есть и такие, которые нормально существуют и в присутствии кислорода, и без него. Например, дрожжевые грибы при спиртовом брожении окисляют пировиноградную кислоту до этилового спирта и оксида углерода (IV):


ВИДЕО ДОМА
Видео YouTube
Вопросы части с

 Метаболизм
Метаболизм
(постоянно происходящий в клетках живых организмов
обмен веществ и энергии)




пластический обмен энергетический обмен
(анаболизм) (ассимиляция) (катаболизм) (диссимиляция)
(биологичекое окисление)
 Реакции биологического синтеза Совокупность реакций расщепления
Реакции биологического синтеза Совокупность реакций расщепления
 сложных молекул на более простые
сложных молекул на более простые
—липидов в эндоплазматической сети, —расщепление липидов, полисахаридов,
—белков на рибосомах, белков и нуклеиновых кислот в лизосомах,
—полисахаридов в комплексе Гольджи эукариот —простых углеводов и жирных кислот
и в цитоплазме прокариот, в митохондриях

—углеводов в пластидах растений.

 Значение
Значение
(Для обеспечения реакций синтеза клетке (В результате процессов катаболизма
требуются существенные затраты энергии, высвобождается энергия. Существенная ее
получаемой при расщеплении веществ.) часть запасается в виде высокоэнергетичных
химических связей АТФ. Запасы АТФ
позволяют организму быстро и эффективно
обеспечивать различные процессы
жизнедеятельности.)
Пластический и энергетический обмены неразрывно взаимосвязаны. Процессы расщепления осуществляют энергетическое обеспечение процессов синтеза, а также поставляют необходимые для синтеза строительные вещества. Правильный обмен веществ поддерживает постоянство химического состава биологических систем, их внутренней среды.
Способность организмов сохранять внутренние параметры неизменными носит название гомеостаза.
Процессы метаболизма происходят в соответствии с генетической программой клетки, реализуя ее наследственную информацию.
Энергетический обмен в клетке. Синтез АТФ
Человек и животные получают энергию за счет окисления органических соединений, поступающих с пищей. Биологическое окисление веществ — это, по сути, медленное горение. Конечные продукты сгорания дров (целлюлозы) — углекислый газ и вода. Полное окисление органических веществ (углеводов и липидов) в клетках также происходит до воды и углекислого газа. В отличие от горения, процесс биологического окисления происходит постепенно. Высвобождающаяся энергия также постепенно запасается в виде химических связей синтезируемых соединений. Некоторая ее часть рассеивается в клетках, поддерживая необходимую для жизнедеятельности температуру.
Синтез АТФ происходит главным образом в митохондриях (у растений еще и в хлоропластах) и обеспечивается в основном энергией, выделяющейся при расщеплении глюкозы, но могут использоваться и другие простые органические соединения — сахара, жирные кислоты и пр.
Гликолиз. Процесс расщепления глюкозы в живых организмах носит название гликолиза (греч. glykys сладкий + lysis расщепление).
Рассмотрим основные его этапы.
На первой, предварительной стадии в лизосомах происходит образование простых органических молекул путем расщепления ди- и полисахаридов. Выделяющееся при этом небольшое количество энергии рассеивается в виде тепла.
Второй этап гликолиза происходит в цитоплазме без участия кислорода и называется анаэробным (бескислородным — греч. ana без + aerвоздух) гликолизом — неполным окислением глюкозы без участия кислорода. Бескислородный гликолиз представляет собой сложный многоступенчатый процесс из десяти последовательных реакций. Каждая реакция катализируется специальным ферментом. В итоге глюкоза расщепляется до пировиноградной кислоты (ПВК):
С6Н12О6(глюкоза) + 2Н3РО4 + 2АДФ = 2С3Н4О3(ПВК) + 2АТФ + 2Н2О
Глюкоза в этом процессе не только расщепляется, но и окисляется (теряет атомы водорода). В мышцах человека и животных две молекулы ПВК, приобретая атомы водорода, восстанавливаются в молочную кислоту С3Н6О3. Этим же продуктом заканчивается гликолиз у молочнокислых бактерий и грибков, применяемый для приготовления кислого молока, простокваши, кефира, а также при силосовании кормов в животноводстве. Процесс превращения ПВК в клетках микроорганизмов и растений в устойчивые конечные продукты называют брожением.
Так, дрожжевые грибки расщепляют ПВК на этиловый спирт и углекислый газ. Этот процесс, называемый спиртовым брожением, используют для приготовления кваса, пива и вина. Брожение других микроорганизмов завершается образованием ацетона, уксусной кислоты и т.д.
Главным результатом анаэробного гликолиза во всех организмах является образование двух молекул АТФ. Высвобождающаяся при расщеплении глюкозы энергия относительно невелика — 200 кДж/моль. Высокоэнергетичные связи АТФ запасают 40% этой величины. Остальные 60% рассеиваются в виде тепла. Основной выход энергии и молекул АТФ происходит на третьем, кислородном этапе гликолиза, называемом еще аэробным дыханием.
III этап. Кислородный гликолиз (аэробное дыхание). При наличии достаточного количества кислорода дальнейший процесс расщепления ПВК происходит уже не в цитоплазме, а в митохондриях, и включает несколько десятков последовательных реакций, каждая из которых обслуживается своим комплексом ферментов.
Молекулы ПВК под действием ферментов (и кофермента НАД — никотинамидадениндинуклеотида) поэтапно окисляются сначала до уксусной кислоты, а затем, в так называемом цикле Кребса (или трикарбоновых кислот), до углекислого газа и воды (медленное горение). В процессе окисления образуются сложные молекулярные соединения с присоединенными к ним атомами водорода. Молекулы-переносчики подхватывают и перемещают электроны этих атомов по длинной цепи ферментов от одного к другому. На каждом шаге электроны вступают в окислительно-восстановительные реакции и отдают свою энергию, которая идет на перемещение протонов на внешнюю сторону внутренней мембраны митохондрии.
В результате оставшиеся протоны и перемещенные электроны оказываются на разных сторонах внутренней мембраны. На мембране создается разность потенциалов.
Фермент, синтезирующий АТФ (АТФ-синтетаза), встроен во внутреннюю мембрану по всей ее толщине. Этот фермент имеет характерную особенность: небольшой каналец в молекулярной структуре. При накоплении на мембране разности потенциалов примерно в 200 мВ ионы Н+ начинают протискиваться через каналец в молекуле АТФ-синтетазы. В процессе энергичного продвижения ионов через фермент происходит синтез АТФ из АДФ с участием фосфорной кислоты.
В химических реакциях кислородного гликолиза освобождается большое количество энергии — 2600 кДж/моль. Существенная ее часть (55%) запасается в высокоэнергетичных связях образующихся молекул АТФ. Остальные 45% рассеиваются в виде тепла (поэтому при выполнении физической работы нам жарко).
Итоговое уравнение кислородной стадии выглядит следующим образом:
2С3Н6О3(молочн.кислота) + 6О2 + 36Н3РО4 + 36АДФ = 6СО2 + 42Н2О + 36АТФ
Таким образом, кислородное расщепление резко увеличивает эффективность энергетического обмена и играет основную роль в аккумулировании энергии. Если гликолиз без участия кислорода дает только 2 молекулы АТФ, то кислородный гликолиз обеспечивает синтез 36 молекул АТФ. В итоге в полном цикле гликолиза на каждую молекулу глюкозы образуется 38 молекул АТФ.
При среднесуточных энергетических затратах в 10 тыс. кДж в организме человека ежедневно синтезируется около 170 кг АТФ, а содержится всего около 50 г АТФ, следовательно, возобновление запаса происходит с частотой 3400 раз в сутки!
Митохондрии способны использовать для синтеза АТФ не только расщепление глюкозы. В их матриксе содержатся также ферменты, расщепляющие жирные кислоты. Особенностью этого цикла является большой энергетический выход — 51 молекула АТФ на каждую молекулу жирной кислоты. Не случайно медведи и другие животные, впадая в спячку, запасают именно жиры. Любопытно, что часть запасаемого жира имеет у них бурый цвет. Такие жировые клетки содержат множество митохондрий необычного строения: их внутренние мембраны пронизаны порами. Ионы водорода свободно проходят через эти поры, и синтез АТФ в клетках бурого жира не происходит. Вся энергия, освобождающаяся в процессе кислородного расщепления жирных кислот, выделяется в виде большого количества тепла, согревающего животных во время долгой зимней спячки.
Бурый жир составляет не более 1-2% массы тела, но повышает производство тепла до 400 Вт на каждый килограмм веса (теплопроизводство человека в состоянии покоя составляет 1 Вт/кг). Запасают жир и верблюды. При постоянном дефиците влаги это вдвойне выгодно, поскольку расщепление жиров дает еще и большое количество воды.
Кроме глюкозы и жирных кислот, митохондрии способны расщеплять аминокислоты, но они — дорогое топливо. Аминокислоты являются важным строительным материалом, из них организм синтезирует свои белки. К тому же использование аминокислот для синтеза АТФ требует предварительного удаления аминогруппы NН2 с образованием токсичного аммиака. Белки и составляющие их аминокислоты используются клеткой для получения энергии только в крайнем случае.
Этиловый спирт тоже может использоваться митохондриями для синтеза АТФ. Но спирт как «топливо» имеет для организма человека свои недостатки, постоянное употребление алкоголя приводит к тяжелым расстройствам, например, к жировому перерождению печени — циррозу.
Фотосинтез — превращение энергии света в энергию химических связей
Автотрофные организмы. В отличие от человека и животных, все зеленые растения и часть бактерий способны синтезировать органические вещества из неорганических соединений. Такой тип обмена веществ называется автотрофным (греч. autos сам + trophe пища). В зависимости от вида энергии, используемой автотрофами для синтеза органических молекул, их делят на фототрофов и хемотрофов. Фототрофы используют энергию солнечного света, а хемотрофы — химическую энергию, высвобождающуюся при окислении ими различных неорганических соединений.
Зеленые растения являются фототрофами. Их хлоропласты содержат хлорофилл, позволяющий растениям осуществлять фотосинтез — преобразование энергии солнечного света в энергию химических связей синтезируемых органических соединений. Из всего спектра солнечного излучения молекулы хлорофилла поглощают красную и синюю часть, а зеленая составляющая достигает сетчатки наших глаз. Поэтому большинство растений мы видим зелеными.
Для осуществления фотосинтеза растения поглощают из атмосферы углекислый газ, а из водоемов и почвы — воду, неорганические соли азота и фосфора. Итоговое уравнение фотосинтеза выглядит довольно просто:
6СО2 + 6Н2О = С6Н12О6(глюкоза) + 6О2,
но всем хорошо известно, что при смешивании углекислого газа и воды глюкоза не образуется. Фотосинтез — сложный многоступенчатый процесс, для прохождения которого необходим не только солнечный свет и хлорофилл, но и ряд ферментов, энергия АТФ и молекулы-переносчики. Выделяют две фазы фотосинтеза — световую и темновую.
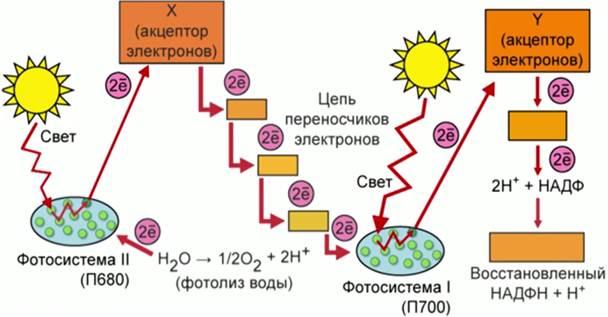
С в е т о в а я ф а з а фотосинтеза начинается с освещения растений светом. Солнечные фотоны, передавая свою энергию молекуле хлорофилла, переводят молекулу в возбужденное состояние: ее электроны, получая дополнительную энергию, переходят на более высокие орбиты. Отрыв таких возбужденных электронов может происходить значительно легче, чем невозбужденных. Молекулы-переносчики захватывают их и перемещают на другую сторону мембраны тилакоида.
Молекулы хлорофилла восполняют потерю электронов, отрывая их от молекул воды. В результате вода расщепляется на протоны и молекулярный кислород:
2Н2О – 4е = 4Н + О2
Процесс расщепления молекул воды до молекулярного кислорода, протонов и электронов под действием света называют фотолизом. Молекулярный кислород легко диффундирует сквозь мембраны тилакоидов и выделяется в атмосферу. Протоны неспособны к проникновению через мембрану и остаются внутри.
Таким образом, снаружи мембраны накапливаются электроны, доставленные молекулами-переносчиками с возбужденных молекул хлорофилла, а внутри — протоны, образовавшиеся в результате фотолиза воды. Возникает разность потенциалов. В мембраны тилакоидов хлоропласта, так же как и во внутренние мембраны митохондрий, встроены ферменты-синтетазы, осуществляющие синтез АТФ. В молекулярной структуре синтетаз растений также имеется каналец, через который могут проходить протоны. При достижении на мембране критической разности потенциалов протоны, влекомые силой электрического поля, протискиваются по канальцу АТФ-синтетазы, затрачивая энергию на синтез АТФ. Соединяясь на другой стороне мембраны с электронами, протоны образуют атомарный водород.
Фотосинтез в хлоропластах весьма эффективен: он дает в 30 раз больше АТФ, чем кислородный гликолиз в митохондриях тех же растений.
Таким образом, во время световой фазы фотосинтеза происходят следующие главные процессы: выделение в атмосферу свободного кислорода, синтез АТФ и образование атомарного водорода.
Протекание дальнейших реакций может происходить и в темноте, потому носит название темновой фазы.
Т е м н о в а я ф а з а. Реакции этой фазы происходят в строме хлоропласта при участии атомарного водорода и АТФ, образовавшихся в световой фазе, а также ферментов, восстанавливающих СО2 до простого сахара — триозы (глицеральдегида) — и синтезирующих из нее глюкозу:
6СО2 + 24Н = С6Н12О6(глюкоза) + 6Н2О
Для образования одной молекулы глюкозы требуется 18 молекул АТФ. Комплекс реакций темновой фазы, осуществляемых ферментами (и коферментом НАД), носит название цикла Кальвина.
Кроме глюкозы, из триозы могут синтезироваться жирные кислоты, аминокислоты и пр. Углеводы и жирные кислоты далее транспортируются в лейкопласты, где из них формируются запасные питательные вещества — крахмал и жиры.
С наступлением темноты растения продолжают процесс фотосинтеза, используя запасенные на свету соединения. Когда этот запас исчерпывается, прекращается и фотосинтез. В ночной темноте растения напоминают по типу обмена веществ животных: они поглощают кислород из атмосферы (дышат) и окисляют при помощи его запасенные днем питательные вещества. На дыхание растения используют в 20-30 раз меньше кислорода, чем выделяют в атмосферу в процессе фотосинтеза.
Количество энергии, производимой растениями, значительно превышает количество тепла, выделяющегося при сжигании всем населением планеты горючих полезных ископаемых. Ежегодно растительность планеты дает 200 млрд. т кислорода и 150 млрд. т органических соединений, необходимых человеку и животным.
Хемосинтез
Большинство бактерий лишены хлорофилла. Некоторые из них являются хемотрофами: для синтеза органических веществ они используют не энергию света, а энергию, высвобождающуюся при окислении неорганических соединений. Такой способ получения энергии и синтеза органических веществ назвали хемосинтезом (греч. chemia химия). Явление хемосинтеза открыто в 1887 г. русским микробиологом С. Н. Виноградским.
Нитрифицирующие бактерии. В корневищах растений, главным образом, бобовых, живут особые клубеньковые бактерии. Они способны усваивать недоступный растениям атмосферный азот и обогащать почву аммиаком. Нитрифицирующие бактерии окисляют аммиак клубеньковых бактерий до азотистой кислоты и далее — азотистую до азотной. В результате растения получают соли азотной кислоты, необходимые для синтеза аминокислот и азотистых оснований.
Водородные бактерии также широко распространены в почвах. Они окисляют молекулы водорода, образующиеся в результате бескислородного окисления органических останков различными микроорганизмами:
2Н2 + О2 = 2Н2О
Железобактерии используют энергию, высвобождающуюся при окислении двухвалентного железа до трехвалентного (закисные соли до окисных).
Серобактерии обитают в болотах и «питаются» сероводородом. В результате окисления сероводорода выделяется необходимая для жизнедеятельности бактерий энергия и накапливается сера. При окислении серы до серной кислоты высвобождается еще часть энергии. Суммарный выход энергии составляет существенную величину — 666 кДж/моль. Огромное количество серобактерий обитает в Черном море. Его воды, начиная со стометровой глубины, насыщены сероводородом.
Гетеротрофный тип обмена веществ. Человек и животные не способны синтезировать необходимые для жизнедеятельности органические вещества из неорганических и вынуждены поглощать их с пищей. Такие организмы называют гетеротрофами (греч. heteros другой). К гетеротрофам относятся также большинство бактерий и грибы. Вещества, поступившие с пищей, разлагаются в организмах животных на простые углеводы, аминокислоты, нуклеотиды, из которых далее синтезируются высокомолекулярные соединения, необходимые для конкретного вида существ в конкретной фазе жизненного цикла. Часть поступивших с пищей молекул расщепляется до конечных продуктов, а высвобождающаяся энергия используется в процессах жизнедеятельности. Некоторое количество энергии рассеивается в виде теплоты, служащей для поддержания температуры тела.
Многие одноклеточные водоросли имеют миксотрофное (смешанное) питание. На свету они фотосинтезируют, а в темноте переходят к фагоцитозу, т.е. становятся гетеротрофами
Пластический обмен. Биосинтез белков. Синтез иРНК
В процессах метаболизма реализуется наследственная информация. Клетка синтезирует только те вещества, которые записаны в ее генетической программе. Каждой группе клеток присущ свой комплекс химических соединений. Среди них особенно важными для организма являются белки.
Многие функции и признаки организма определяются его набором белков. Белки-ферменты расщепляют пищу, отвечают за поглощение и выделение солей, синтезируют жиры и углеводы, производят множество других биохимических превращений. Белки определяют цвет глаз, рост — словом, внешнюю специфичность организмов. Большинство белков, выполняющих одни и те же функции, несколько различны даже у особей одного и того же вида (к примеру, белки групп крови). Но некоторые однофункциональные белки могут иметь сходное строение у далеких групп организмов (к примеру, инсулин собаки и человека).
В процессе жизнедеятельности белковые молекулы постепенно разрушаются, теряют свою структуру — денатурируют. Их активность падает, и клетки заменяют их новыми. В организмах постоянно происходит синтез необходимых белков.
Биосинтез белковых молекул — сложный ферментативный процесс, начинающийся в ядре и заканчивающийся на рибосомах. Центральную функцию в нем выполняют носители генетической информации — нуклеиновые кислоты ДНК и РНК.
Генетический код. Последовательность нуклеотидов ДНК задает последовательность аминокислот в белках — их первичную структуру. Молекулы ДНК являются матрицами для синтеза всех белков.
Отрезок ДНК, несущий информацию о первичной структуре конкретного белка, называют геном. Соответствующую последовательность нуклеотидов — генетическим кодом белка.
Идею о том, что наследственная информация записана на молекулярном уровне, а синтез белков идет по матричному принципу, впервые высказал еще в 1920-х годах русский биолог Н. К. Кольцов. В настоящее время код ДНК полностью расшифрован. В этом заслуга известных ученых: Г. Гамова (1954), а также Ф. Крика, С. Очоа, М. Ниренберга, Р. Холи и К. Хорана (1961-65). Значительную часть свойств генетического кода установил английский физик Ф. Крик, исследуя бактериофагов.
Код триплетен. Каждая аминокислота в генетическом коде задается последовательностью трех нуклеотидов — триплетом, или кодоном. Различных нуклеотидов в ДНК четыре, следовательно, теоретически возможных кодонов — 64 (43). Большинству аминокислот соответствует от 2 до 6 кодонов — код, как говорят, вырожден. Чем чаще аминокислота встречается в белках, тем, как правило, большим числом кодонов она кодируется. Оставшиеся три кодона вместе с кодоном метионина (АУГ) служат знаками препинания при считывании информации — указывают начало и конец матриц конкретных белков. Если белок имеет несколько полимерных цепей (образующих отдельные глобулы), то знаки препинания выделяют полипептидные звенья. Считывание каждого звена происходит непрерывно, без знаков препинания и пропусков — триплет за триплетом.
Код однозначен. Кроме триплетности, генетический код наделен рядом других характерных свойств. Его кодоны не перекрываются, каждый кодон начинается с нового нуклеотида, и ни один нуклеотид не может прочитываться дважды. Любой кодон соответствует только одной аминокислоте.
Код универсален. Генетическому коду свойственна универсальность для всех организмов на Земле. Одинаковые аминокислоты кодируются одними и теми же триплетами нуклеотидов у бактерий и слонов, водорослей и лягушек, черепах и лошадей, птиц и даже человека. Несколько отличаются (на 1-5 кодонов) только коды митохондрий некоторых организмов, ряда дрожжей и бактерий.
Ошибка хотя бы в одном триплете приводит к серьезным нарушениям в организме. У больных серповидной анемией (их эритроциты имеют не дисковую, а серповидную форму) из 574 аминокислот белка гемоглобина одна аминокислота заменена другой в двух местах. В результате белок имеет измененную третичную и четвертичную структуру. Нарушенная геометрия активного центра, присоединяющего кислород, не позволяет гемоглобину эффективно справляться со своей задачей — связывать кислород в легких и снабжать им клетки организма.
Транскрипция. Синтез белка происходит в цитоплазме на рибосомах. Генетическую информацию от хромосом ядра к месту синтеза переносят иРНК:
ДНК — иРНК — белок
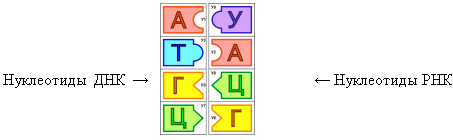
Информационная РНК синтезируется на отрезке одной из нитей ДНК как на матрице, хранящей информацию о первичной структуре конкретного белка или группы белков, выполняющих одну функцию. В основе синтеза лежит принцип комплементарности: напротив Цднк встает Грнк, напротив Гднк — Црнк, напротив Аднк — Урнк, напротив Тднк — Арнк. Затем мономерные звенья связываются в полимерную цепь. Таким образом, иРНК становится точной копией второй нити ДНК (с учетом замены Т- У). Молекула иРНК имеет одноцепочечную структуру, она в сотни раз короче ДНК.
Процесс перенесения генетической информации на синтезируемую иРНК носит название транскрипции. Перед началом каждого гена или группы однофункциональных генов расположена последовательность нуклеотидов, называемая инициатором (содержит кодон АУГ). В этой последовательности есть участок (промотор) для присоединения фермента РНК-полимеразы, осуществляющего транскрипцию. Полимераза распознает промотор благодаря химическому сродству. В конце матрицы синтеза находится стоп-кодон (один из трех в таблице), или терминатор.
В ходе транскрипции РНК-полимераза в комплексе с другими ферментами разрывает водородные связи между азотистыми основаниями двух нитей ДНК, частично раскручивает ДНК и производит синтез иРНК по принципу комплементарности. На одной ДНК «работают» сразу несколько полимераз.
Готовая молекула иРНК после небольшой перестройки связывается в комплекс со специальными белками и транспортируется ими через ядерную оболочку на рибосомы. Эти белки выполняют и другую функцию — они защищают иРНК от действия различных ферментов цитоплазмы. В прокариотической клетке ДНК не отделена от цитоплазмы, и синтез белков рибосомы начинают еще во время транскрипции.
Транспортные РНК. Необходимые для синтеза белков аминокислоты всегда имеются в составе цитоплазмы. Они образуются в процессе расщепления лизосомами белков. Транспортные РНК связывают аминокислоты, доставляют их на рибосомы и производят точную пространственную ориентацию аминокислот на рибосоме.
Рассмотрим устройство тРНК, позволяющее ей успешно выполнять свои сложные функции. В цепочке, состоящей из 70-90 звеньев, имеется 4 пары комплементарных отрезков из 4-7 нуклеотидов — А, Б, В и Г. Комплементарные участки связываются водородными связями попарно (как в молекуле ДНК). В результате нить тРНК «слипается» в четырех местах с образованием петлистой структуры, напоминающей лист клевера. В верхушке «листа» располагается триплет, код которого комплементарен кодону иРНК, соответствующему транспортируемой аминокислоте. Так, если в иРНК код аминокислоты валина ГУГ, то на вершине валиновой тРНК ему будет соответствовать триплет ЦАЦ. Комплементарный триплет в тРНК называют антикодоном.
Специальный фермент распознает антикодон тРНК, присоединяет к «черенку листа» определенную аминокислоту (в нашем примере — валин), и затем тРНК перемещает ее к рибосоме. Каждая тРНК транспортирует только свою аминокислоту.
Синтез белковой цепи на рибосоме
Матричные процессы составляют основу способности живых организмов к воспроизведению. В клеточном ядре происходит удвоение ДНК. Новая молекула воспроизводится на матрице старой и представляет собой ее точную копию. Информационная РНК синтезируется на матрице ДНК в виде точной копии одного из участков ДНК. Далее на матрице иРНК происходит синтез белков.
Трансляция. Перевод последовательности нуклеотидов иРНК в последовательность аминокислот синтезируемых белков называют трансляцией.
В активном центре рибосомы размещаются два триплета иРНК и соответственно две тРНК. Рибосома перемещается по иРНК не плавно, а прерывисто, триплет за триплетом. На каждом шаге присоединяется новая аминокислота. Транспортные РНК перемещаются со своей аминокислотой к рибосоме и «примеряют» свой антикодон к очередному кодону иРНК, находящемуся в активном центре (комплементарные нуклеотиды хорошо подходят друг другу, между ними возникают водородные связи). Если антикодон оказывается не комплементарным, то тРНК удаляется в цитоплазму к другим рибосомам. Если же он оказывается комплементарным, то тРНК присоединяется к кодону водородными связями.
Далее особый фермент рибосомы подсоединяет синтезируемую полипептидную цепь к «прибывшей»аминокислоте. Ее транспортная РНК продолжает удерживать всю формирующуюся белковую цепь на кодоне иРНК до прихода следующей тРНК. Освободившаяся тРНК перемещается в цитоплазму «на поиск» аминокислот. Рибосома перескакивает на следующий триплет иРНК, и процесс повторяется. Интервал между перескакиваниями продолжается не более 1/5 — 1/6 с, а вся трансляция среднего белка — 1-2 минуты.
На одной нити иРНК «трудятся» сразу несколько рибосом. На иРНК, содержащей информацию о белке гемоглобине, размещается до 5 рибосом, на некоторых других иРНК — до 20 рибосом. Когда в активном центре рибосомы оказывается один из трех триплетов, кодирующих знаки препинания между генами, синтез белка завершается.
Освободившаяся рибосома отправляется на другую иРНК. Рибосомы универсальны и могут синтезировать полипептиды по любой матрице иРНК. Субъединицы рибосом соединяются только для синтеза белка, после окончания синтеза они вновь разъединяются.
Сворачивание в спираль и приобретение третичной структуры происходит по мере синтеза белковой цепи, поэтому зачастую невозможно восстановление денатурированных белков со сложной пространственной структурой, даже если их первичная структура сохранена. Сворачивание белковой цепи целой молекулы происходит по-иному, и белок оказывается неактивным.
Многие белки — например, пищеварительные ферменты — очень активны и способны переварить саму клетку, поэтому синтезируемые молекулы белков сразу попадают в эндоплазматическую сеть (к мембране которой прикреплены синтезирующие белки рибосомы) и по ее каналам перемещаются к комплексу Гольджи, а от него в тот участок клетки или в ту часть организма, где требуется этот вид белков. Синтез небелковых соединений клетка осуществляет в два этапа. Сначала рибосомы производят трансляцию специфического белка-фермента. Затем при его участии образуется молекула необходимого соединения — углевода, жиров и т.д. Сходным образом синтезируются и другие соединения: витамины, небелковые гормоны и пр.
Процесс синтеза белков требует затрат большого количества специфической энергии АТФ, которую нельзя заменить иным источником. Только на присоединение каждой аминокислоты к тРНК расходуется энергия одной молекулы АТФ, а в среднем белке 400-500 аминокислот. Общее количество затрачиваемой энергии существенно превышает суммарную химическую энергию образующихся пептидных связей между молекулами аминокислот. По этой причине белки редко используются клеткой в качестве топлива: слишком велики затраты на их изготовление.
Регуляция транскрипции и трансляции. Клетки различных тканей живых организмов сильно отличаются друг от друга по строению и функциям, но все они произошли от единственной зиготы (оплодотворенной яйцеклетки) в результате многократного деления.
Перед каждым актом деления ДНК реплицируется в две точные копии. Ученым удалось экспериментально доказать, что дифференцированные клетки организма имеют одинаковый набор ДНК, не утрачивая ни одной из его частей в процессе делений.
В 1958 г. Ф. Стюарт впервые вырастил из единственной клетки взрослой моркови целое растение. В 1968 г. Дж. Гордону удалось пересадить ядра из клеток кишечника головастика в яйцеклетки лягушки, лишенные собственных ядер, и вырастить из них нормальных лягушек. Следовательно, клетки всех тканей организма имеют один и тот же набор генов, одну и ту же информацию о строении белков.
Однако клетки каждой ткани производят свои белки, свои ферменты. Гемоглобин образуется только в эритроцитах, белковый гормон роста синтезируется только в клетках гипофиза, зрительный белок опсин — в клетках сетчатки глаза, а инсулин — в клетках поджелудочной железы.
Такое разнообразие происходит по причине того, что клетки каждой ткани реализуют только свою часть информации ДНК. В разных клетках происходит транскрипция разных участков ДНК, синтезируются разные иРНК, по которым воспроизводятся разные белки. Более того, специфичные для клеток белки производятся не все сразу, а по мере необходимости. В организмах имеются удивительно точные механизмы«включения» и «выключения» генов на разных этапах жизненного цикла клетки от ее появления до деления.
Геном эукариот устроен намного сложнее генома прокариот. Это связано с увеличением сложности регуляции генетических процессов, а не с увеличением количества структурных белков и ферментов. В клетках высокоразвитых организмов только 10% всех генов ответственны за синтез ферментов и структурных белков, остальные 90% составляют «административный аппарат» клетки.
Кроме того, существуют системы, регулирующие синтез веществ в организме как едином целом. В клетках желез внутренней секреции вырабатываются гормоны, которые разносятся с кровью по всему телу. Эти гормоны регулируют процессы синтеза иРНК и трансляцию именно в тех клетках, для которых они предназначены. На поверхности мембраны клеток есть рецепторы для «своих» гормонов. Связываясь с рецепторами, гормоны управляют активностью различных систем клетки, регулирующих обмен. В результате может изменяться как транскрипция конкретных генов, так и синтез белков на рибосомах.
Даже синтезированные иРНК могут долгое время не транслироваться, если нет «команды» от гормонов. Каждый гормон через определенные системы клеток активирует свои гены. Так, адреналин дает сигнал на синтез ферментов, расщепляющих гликоген до глюкозы, интенсивно потребляемой мышцами при физической нагрузке. Инсулин выполняет обратную функцию, он участвует в синтезе гликогена из глюкозы в клетках печени.
Как транскрипция, так и трансляция могут подавляться различными химическими веществами, относящимися к классу антибиотиков (греч.anti против + bios жизнь). Например, эритромицин и стрептомицин подавляют синтез белка на рибосомах инфекционных бактерий, а бледная поганка содержит вещество, подавляющее функционирование РНК-полимеразы человека. Прекращение полимеразой синтеза иРНК приводит к тяжелейшим последствиям.
Подведем итоги. В воспроизведении белков в живых существах участвует ДНК, информационная РНК, транспортные РНК для 20 аминокислот (всего 61, по количеству кодонов), рибосомы, состоящие из нескольких рибосомных РНК (3 у прокариот и 4 у эукариот) и десятков различных молекул белка, а также целый комплекс ферментов. Для осуществления синтеза одной белковой молекулы необходим тонкий, специфический подвод энергии посредством более чем 1000 молекул АТФ. Практически вся клетка участвует в синтезе, нарушение строения хотя бы одного из компонентов нарушает процесс воспроизведения белковых молекул.
Для современных ученых удивителен сам факт функционирования этой сложной системы в организме. Доказана абсолютная невозможность самообразования молекул РНК и даже ее более простой составляющей — рибозы. Возможность же самосборки белков в гипотетическом первобытном океане при возникновении в нем жизни квалифицированные биохимики совершенно исключают. Так познание внутриклеточных процессов приводит к мысли о богосотворенности мира и приближает нас к очевидности Божия бытия.