Китообразные
1. Как это ни удивительно, являются родственниками парнокопытых. Предки китов (вероятно, мезонихии) некогда обитали на суше, способность к жизни в воде стала вторичным приобретением.
2. Передние конечности превратились в ласты, задние исчезли вовсе.
3. Волосяной покров исчез, остались лишь редкие незначительные волоски.
4. Сальные и потовые железы отсутствуют.
5. Нет ушных раковин, ноздри «съехали» с передней части головы в верхнюю и превратились в дыхала.
6. Зубы не дифференцированы или отсутствуют. У беззубых китов вместо них развился китовый ус — ороговевшие пластины на небе, являющиеся цедильным аппаратом.
7. Ключиц нет.
8. Имеется огромный раздвоенный хвостовой плавник.
9. Детеныши рождаются прямо в воде и, будучи хорошо развитыми, следуют за матерью.
10. Представители: подотряд зубатые киты (дельфин афалина, касатки, белухи, кашалоты, дельфин-белобочка), подотряд беззубые (усатые) киты (киты гренландский, южный, настоящий, горбатый, синий, серый).
Парнокопытные. Подотряд жвачные
1. Большинство имеют рога — от небольших выростов до огромных затейливых образований, как, например, у лося. У оленей рога состоят из костной ткани. Рога могут быть также из кератина (рогового вещества), как у носорога. У овцы рога растут всю жизнь, новыми не сменяются, а вот оленевые (лоси, олени) их меняют. Молодые рога, трубчатые, пронизанные кровеносными сосудами, называются пантами.
2. Имеют только два пальца, третий и четвертый, прочие редуцировались.
3. Желудок четырехкамерный, состоит из рубца, сетки, книжки, сычуга.
4. Пища долго пережевывается, превращаясь в ком жвачки — отсюда и называние подотряда.
5. Резцы растут только на нижней челюсти. Клыков нет, коренные зубы с широкой жевательной поверхностью.
6. Присуще фалангохождение. Ключица отсуствует.
7. Представители подотряда: быки и коровы, козлы, бараны, зубры, лоси, олени, жирафы, антилопы; дикий тур — предок крупного рогатого домашнего скота.
Подотряд нежвачные
1. Всеядные животные.
2. Во рту выделяются хорошо развитые острые клыки, которые растут постоянно.
3. Желудок у свиньи и кабана имеет простое строение, одну камеру. У пекари и бегемота трехкамерный.
4. У свиней и кабанов нос заканчивается пятачком, приспособленным для рытья почвы.
5. Представители подотряда: свиньи, бегемоты, пекари; дикий кабан — предок домашней свиньи.
Хочешь сдать экзамен на отлично? Жми сюда — репетитор по биологии: подготовка к ЕГЭ
Китообразные
Вторичноводные млекопитающие.
Китообразные — близкие родственники парнокопытных, в частности, гиппопотамов.

Особенности внешнего строения Китообразных

Особенности внутреннего строения Китообразных
1) сальные и потовые железы отсутствуют
2) зубы не дифференцированы или отсутствуют. У беззубых китов развился китовый ус — ороговевшие пластины на небе, являющиеся цедильным аппаратом
3) ключиц нет
4) большая ёмкость (объем) лёгких
5) наличие клапанов, закрывающих ноздри
6) высокая концентрация миоглобина в мышцах
7) высокая концентрация гемоглобина в крови
 толстый слой подкожного жира
толстый слой подкожного жира
9) задние конечности исчезли

Дыхало — дыхательное отверстие у китообразных, находящееся, как правило, на самом верху головы и представляющее собой видоизменённые и смещённые на макушку носовые отверстия (ноздри). У всех китообразных дыхало снабжено клапаном, благодаря которому плотно закрывается при погружении животного и открывается при его всплытии. Когда кит поднимается на поверхность, выдыхаемый им воздух, насыщенный водяным паром, соприкасаясь с холодным наружным, образует столб пара (так называемый фонтан). Форма фонтана различается в зависимости от вида кита.
Под водой дыхательные отверстия закрываются двумя маленькими клапанами, а так как воздушный проход не соединен со ртом, нет опасности, что вода попадет в легкие.
Детеныш рождается под водой. Сразу после появления на свет малыш всплывает и делает глубокий вдох, максимально наполняя легкие кислородом.
Примеры заданий формата ЕГЭ по биологии
Какие особенности строения скелета позвоночного животного, изображенного на рисунке, доказывают его наземное происхождение? Приведите доказательства. С какой группой современных позвоночных животных у него проявляется сходство во внешнем строении? Как называется эволюционный процесс, в результате которого сформировалось это свойство? Ответ обоснуйте.

1) грудные плавники имеют сходство в строении с конечностью наземного типа (пояс конечности и свободная конечность из трех отделов: плеча, предплечья, кисти);
2) имеются рудиментарные кости тазового пояса, свидетельствующие о наличии у предков задних конечностей;
3) сходство с рыбами по форме тела и плавников;
4) название процесса — конвергенция; это формирование сходных признаков у неродственных групп, обитающих в одинаковых условиях.
Известно, что вторичноводные млекопитающие (дельфины, киты) обладают значительными запасами жира. Объясните, в чем заключается их адаптивное значение.
1) энергетический запас на случай длительного отсутствия пищи
2) повышение плавучести
3) теплоизоляция
4) придание обтекаемости форме тела
1. Окунь — первичноводное животное, кит — вторичноводное животное. 2. У первичноводных животных есть органы боковой линии, воспринимающие давление, направление движения, скорость течения воды. 3. Передние конечности кита превратились в грудные плавники 4. Задние конечности кита видоизменены в хвостовой плавник. 5. У окуня и кита жаберное дыхание. 6. У китов к крестцовому отделу позвоночника прикреплены тазовые кости.
4 — задние конечности кита утрачены (хвостовой плавник — складка кожи).
5 — у кита легочное дыхание (а не жаберное).
6 — у китообразных крестцовый отдел позвоночника и пояс задних конечностей не развит.
Просмотров: 5266
Интерактивный урок для обучающихся
9-11 классов «Биология и эволюция китообразных»
Занятие предназначено для проведения урока по биологии
в старших классах, может быть интегрировано в следующие темы: эволюция животного
мира, разнообразие млекопитающих, идиоадаптация и др. Кроме того, можно провести
данный урок в рамках работы биологических и экологических объединений дополнительного
образования.
Сценарий урока
Материально-техническое обеспечение: компьютер
учителя, медиапроектор с экраном или интерактивная доска, компьютеры/планшеты/смартфоны
для обучающихся (по возможности), доступ в интернет.
Урок построен в виде семинарского занятия с элементами
практической работы в виде выполнения двух интерактивных заданий.
Слайд 1. Сегодня мы познакомимся с удивительным
миром китов и дельфинов. Эти животные живут в морях, океанах, а некоторые – в пресноводных
озёрах и реках. И несмотря на то, что они всю свою жизнь проводят в воде, не выходя
на сушу, имеют обтекаемую торпедообразную форму тела, напоминающую рыбу, являются
самыми настоящими теплокровными млекопитающими. Мы знаем много фактов и мифов об
этих удивительных животных! Так давайте рассмотрим особенности китообразных, их
происхождение и удивительные способности на нашем занятии.
Слайд 2. Рассмотрим таксономическое положение китообразных.
Как уже говорилось выше, все китообразные относятся к классу Млекопитающие. Как
лисы, волки, лошади, приматы, коровы и человек. Дельфины и киты выделены в отдельный
отряд – CETACEA – китообразные. Китообразные в свою очередь делятся на два подотряда
– УСАТЫЕ и ЗУБАТЫЕ киты. К усатым китам относятся гигантские млекопитающие, имеющие
особые приспособления в ротовой полости, называемые китовый ус. С помощью него киты
отфильтровывают воду и питаются мелкими рачками, рыбой. Зубатые киты — самые настоящие
хищники. Своими острыми зубами они легко ловят рыбу, головоногих моллюсков, а некоторые,
такие как плотоядые косатки — тюленей и даже китов. Представителями подотряда усатых
китов являются такие животные как — синий кит (самое большое млекопитающее на земле),
гренландский кит, финвал, серый кит, горбач и другие. Зубатые же киты делятся на
4 семейства: дельфиновые, морские свиньи, клюворылы и кашалоты. Посмотрите на картинки
и скажите по каким признакам разные зубатые киты отнесены в разные семейства?
Слайд 3 А теперь посмотрим филогенетические ряды
современных млекопитающих. На данном рисунке видно, какие животные ближе всего друг
другу. Если такую картинку рассмотреть поближе, то становится понятно, что китообразные
— родственники копытным животным, а самым близким родственником является бегемот!
При этом мы знаем, что все копытные животные являются травоядными, а вот киты и
дельфины стали хищниками.
Слайд 4. Как же это произошло? Дело в том, что
предок дельфинов и китов был обитателем суши и даже возможно травоядным. Но после
он стал вести сначала околоводный, а впоследствии и водный образ жизни. В океане
биомасса животных организмов достаточно высока, растения присутствуют только в зонах
с доступом солнечного света. Видимо поэтому предки китов изменили свои пищевые предпочтения
и стали питаться животными.
Слайд 5. А вот и первый достоверно известный предок
современных китообразных. Встречайте – ПАКИЦЕТУС. Останки этих млекопитающих с признаками
китообразных впервые появляются в отложениях начала среднего эоцена. Первым из них
был Pakicetus из семейства Protocetidae, водное животное продолговатой формы
с редуцированными задними конечностями и длинной мордой. По-видимому, он был еще
вполне способен передвигаться по суше, но значительную часть времени проводил в
воде. С китами пакицетуса роднит устройство уха: слуховая булла пакицетуса, как
и у кита, образовалась исключительно из барабанной кости. Форма ушной области пакицетуса
весьма необычна и находит аналоги только у китообразных.
Слайд 6. Следующей важной эволюционной формой китообразных
стал ПЕРЕГОЦЕТУС. Посмотрите, он уже имел перепонки на конечностях, прекрасно плавал,
вытянутыми челюстями с острыми зубами легко ловил рыбу. Но при этом для отдыха он
все же выбирался на сушу.
Слайд 7. Посмотреть, как изменялась морфология
китообразных в процессе эволюции можно на данной картинке. Обратите внимание, что
помимо изменения формы тела, конечностей, головы животные изменяли и свои размеры.
Так первый предок в сравнении с фигурой человека выглядит как большая собака, а
позже размеры стали значительно увеличиваться. Посмотрите на современную косатку
– этот зубатый хищник имеет очень впечатляющие размеры!
Слайд 8. Как видите 50 миллионов лет – это достаточный
срок для очень заметных изменений только одной ветви млекопитающих! Представьте,
что будет с нами и с китообразными еще через 50 миллионов лет!
Дети могут
пофантазировать, применяя законы эволюции, конечно, как будет выглядеть
животный мир в будущем и какова роль антропогенных факторов, влияющих на эту
эволюцию.
Слайд 9. А теперь рассмотрим особенности биологии
китообразных. Как вы уже узнали, эти животные постоянно живут в воде, не выходят
на сушу. Поэтому им пришлось преодолеть несколько трудностей, сопряженных с жизнью
в воде. Животные научились быстро двигаться в водной среде, спать, дышать, осуществлять
терморегуляцию, общаться друг с другом, спариваться, рожать и выкармливать детёнышей,
ориентироваться в пространстве, глубоко нырять.
Слайд 10. Все вы видели дельфинов в кино или в
природе. Насколько легко, стремительно и грациозно плывут эти животные! Посмотрите
видеофрагмент, на котором показано движение дельфинов-белобочек. Как можно охарактеризовать
их движение? Какие органы задействованы у дельфинов для осуществления движения?
Конечно, китообразные имеют очень хорошие приспособления для этого: широкую лопасть
хвоста и развитые мышцы, обтекаемую форму тела, гладкую, постоянно обновляющуюся
кожу. А какие приспособления может применить человек, чтобы плавать как дельфин?
(ласты, облегающий костюм для плавания, движение «дельфином»)
Слайд 11. У всех китообразных есть удивительная
способность спать под водой не так, как это делают наземные млекопитающие и человек.
Ведь когда человек или животное спит ритмы обоих полушарий мозга замедляются, мы
не контролируем многие процессы, как бы находясь без сознания. У нас есть парадоксальный
сон – мы видим сны и даже можем двигаться во сне. Китам и дельфинам нельзя отключаться
полностью, ведь процесс их дыхания должен быть контролируемым. Китообразные дышат
атмосферным воздухом и для дыхательного акта им надо всплыть на поверхность. Но
если они полностью заснут, то погибнут от асфиксии (задохнутся) или утонут. Поэтому
киты и дельфины спят только наполовину, то есть, когда одно полушарие их мозга спит
– другое бодрствует, а через какое-то время меняются – засыпает бодрствующее полушарие,
просыпается спящее. Таким образом, эти животные постоянно контролируют своё дыхание
и окружающую среду. Кроме того, у них отсутствует парадоксальный сон!
Слайд 12. В воде сложно оставаться теплым, тело
очень быстро охлаждается. А как же киты решили эту проблему? Ведь у них отсутствует
шерстный покров. Дело в том, что у китообразный очень развит слой подкожного жира.
Так, у гренландского кита толщина жирового слоя может составлять 40см! Но хорошо,
если ты живешь в высоких широтах, там в основном холодно. А как же быть животным,
обитающим в тропической зоне, или нашим черноморским дельфинам в летний период?
Из-за жира они могут перегреться. Но китообразные решили и эту проблему. В их плавниках,
особенно в спинном и хвостовом, есть особые кровеносные сосуды, называемые КОМПЛЕКСНЫЕ,
и отсутствует жировой слой, то есть они легко охлаждаются. Они-то и осуществляют
терморегуляцию. Как? Комплексный сосуд представляет из себя комплекс артерий и вен.
При этом артерия окружена сетью из венозных сосудов (см. рисунок). Венозная кровь
плавника охлаждается в незащищенном жиром плавнике и в свою очередь охлаждает артерию,
которая несет уже охлажденную кровь в тело. Нормальная температура тела кита составляет
370С (как у человека).
Слайд 13. Вода в океане не везде прозрачная, да
и в глубине мало света, а в реках и озёрах, где живут пресноводные дельфины вода
бывает чрезмерно мутной. В мутной воде и в тёмных глубинах для восприятия окружающей
среды зрение играет второстепенную роль. Первостепенное значение для
ориентации в воде имеет эхолокация, так как акустические характеристики подводной
среды очень благоприятны. Эхолокация – это прием акустических сигналов, отраженных
от различных объектов. Дельфины, посредством эхолокации способны различить довольно
тонкую проволоку, уловить движение рыбы или сородичей и даже определить внутреннюю
структуру предмета. Как настоящий УЗИ аппарат! Для дельфинов не составляет труда
обнаружить в воде очень мелкие шарики (три миллиметра в диаметре) и отличить их
материал. Для этих целей у дельфинов сложная комбинация различных органов. Звук
они издают воздушными полостями в верхних дыхательных путях, далее сигнал концентрируется
в особом жировом органе в области лба МЕЛОНЕ, получается направленный устойчивый
звук. Отраженный от объекта сигнал дельфин принимает нижней челюстью, в которой
есть жировой канал, отлично проводящий звук. Далее сигнал попадает через акустическое
окно во внутреннее ухо, где преобразованный импульс попадает в центральную нервную
систему и анализируется.
Слайд 14. В результате постоянного нахождения в
воде дельфин вынужден дышать, не выходя для этого на сушу. И да, китообразные дышат
атмосферным кислородом, органом дыхания являются лёгкие. Поэтому вся его жизнь между
дыхательными актами проходит на задержке дыхания. Китообразные контролируют акт
вдоха/выдоха и делают это сознательно. Ведь если животное случайно сделает вдох
под водой – оно утонет. Что бы не менять положение тела в пространстве во время
дыхательного акта нос дельфина в процессе эволюции переместился с морды на спину.
Дыхательное отверстие называется дыхало, а морда – рострум. Выдыхая, китообразные
освобождают от газа почти весь объём лёгких, поэтому их фонтан содержит много органических
примесей: клетки эпителия, гормоны и ферменты, микроорганизмы.
Слайд 15. Зрение у дельфинов тоже своеобразное.
Они неплохо видят в прозрачной воде и на воздухе. Но в разных средах у них меняется
кривизна глазного яблока, приспосабливаясь к воздушной или водной среде. И зрачок
у китообразных удивительный – он подковообразной формы.
Слайд 16. Еще одна важная задача, связанная с дыханием
– нырять глубоко и надолго. Рекорды современного фридайвинга – ныряние на глубину
до 130м в моноласте, а с большим грузом – 214 м, задержка дыхания на 11 минут –
являются нормой для большинства дельфинов и усатых китов. Но некоторые виды, специализирующиеся
на питании глубоководными кальмарами – рекордсмены среди китообразных. Например,
кашалот может нырять на глубины более 2.5 км и находиться под водой полтора часа.
Слайд 17. Как избегают кессонной болезни животные,
которые постоянно ныряют: тюлени, пингвины, киты? Этот вопрос давно интересовал
физиологов, и они, разумеется, нашли объяснения: пингвины ныряют ненадолго, тюлени
перед погружением выдыхают, у китов воздух на глубине выдавливается из легких в
большую несжимаемую трахею. А если в легких нет воздуха, то азот не попадает в кровь.
Еще одно объяснение отсутствия у китов кессонной болезни предложили недавно специалисты
из Университета Тромсё (University of Tromsø) и Университета Осло (University
of Oslo). По мнению ученых, китов защищает разветвленная сеть тонкостенных артерий,
снабжающая кровью головной мозг.
Эту обширную сосудистую сеть, которая занимает значительную
часть грудной клетки, пронизывает позвоночник, область шеи и основание головы китообразных,
впервые описал в 1680 году английский анатом Эдвард Тайсон в труде «Анатомия морской
свиньи, вскрытой в Грешем-колледже; с предварительным обсуждением анатомии и естественной
истории животных», и назвал ее чудесной сетью — retia mirabilia. Впоследствии
эту сеть описывали разные ученые у разных видов, в том числе у бутылконосого дельфина
Tursiops truncates, нарвала Monodon monoceros, белухи Delphinapterus
leucas и кашалота Physeter macrocephalus. Исследователи выдвигали разные
предположения о функциях чудесной сети, самая популярная заключается в том, что
она регулирует артериальное давление.
Относительно толстые артерии, образующие видимую невооруженным
глазом сеть, дробятся на множество мельчайших сосудов, которые сообщаются друг с
другом через тонкостенные синусы. Эти сосудистые структуры утоплены в жировую ткань.
Именно через эту сеть проходит кровь, поступающая в мозг.
В стенках артерий сети мало мышечных клеток, и они не
иннервируются, т. е. просвет сосудов всегда постоянен. Но исследователи отмечают,
что он и не нуждается в регуляции, поскольку мозгу необходимо постоянное количество
крови.
Общая площадь сечения всех сосудов и сосудиков так велика,
что скорость течения крови в сети падает почти до нуля, что существенно увеличивает
возможности обмена между кровью и окружающей жировой тканью через сосудистую стенку.
Исследователи предположили, что у выныривающих китообразных азот из перенасыщенной
крови диффундирует в жир, в котором он растворим в шесть раз лучше, чем в воде.
Таким образом диффузия в retia mirabilia предотвращает образование азотных
пузырьков, которые могут достичь мозга и вызвать кессонную болезнь.
Среди работ, на которые ссылаются норвежские исследователи,
есть и статья ведущего научного сотрудника Тихоокеанского океанологического института
им. В. И. Ильичева ДВО РАН Владимира Васильевича Мельникова, который в 1997 году
вскрывал кашалота.
Он пишет, что retia mirabilia у
кашалота развита сильнее, чем у других китообразных (разумеется, тех, которых анатомировали).
А ведь именно кашалот — чемпион среди китообразных по глубине и длительности погружения.
Возможно, этот факт косвенно подтверждает гипотезу норвежских ученых.
Фото из статьи: Arnoldus
Schytte Blix, Lars Walløe and Edward B. Messelt. On
how whales avoid decompression
sickness and why they sometimes strand // J. Exp Biol, 2013,
doi:10.1242/ jeb.087577.
Слайд 18. Как и все млекопитающие китообразные
вскармливают своих детёнышей молоком. Молоко у них очень жирное и питательное, по
составу сильно отличается от коровьего молока. И китята, и дельфинята на таком молоке
растут очень быстро! Для продукции молока самка имеет парные молочные железы, расположенные
в специальных складках кожи на животе. Когда детеныш прикасается к молочной железе
она открывается и молоко впрыскивается ему в рот под давлением. Акт сосания отсутствует.
Длительность вскармливания детёнышей зависит от вида китообразным.
Слайд 19. А теперь давайте познакомимся с представителями
китообразных Чёрного моря. В Черноморских водах России обитает три вида китообразных.
И все они в той или иной степени являются уязвимыми, то есть численность их заметно
снижается. Черноморская афалина — самый большой черноморский дельфин — длина
тела может достигать 3,5 м, вес 150-350 кг. Окрас серый, рострум средней длины,
серповидный (загнутый) спинной плавник, округлые формы тела. Часто встречаются в
стаях 5-10 особей и больше. Афалины относятся к виду под угрозой исчезновения, статус
в Красной книге РФ — редкий. Дельфин-белобочка средний по размеру черноморский
дельфин — длина тела может достигать 2,4 метра, вес 100-130 кг. На боках ярко выраженный
белый окрас. Длинный рострум, треугольный спинной плавник серповидной формы (слегка
загнут), более заостренные формы тела по сравнению с афалиной. Часто встречаются
в больших стаях 15-30 особей и больше. Белобочка относится к редкому виду и занесена
в Красную книгу республики Крым. Черноморская морская свинья, или азовка —
самое маленькое китообразное Черного моря — длина тела может достигать 1,8 метра,
вес 30-60 кг. Иногда азовку принимают за детёнышей более крупных дельфинов! Окрас
у них темный, нет выраженного рострума, как у других китообразных, спинной плавник
треугольный – похож на крышу домика, не загнутый, формы тела округлые. В основном
встречаются по одиночке или в небольших стаях 2-5 особей. Азовка относятся к виду
под угрозой исчезновения, статус в Красной книге РФ — редкий.
Как отличить разные виды дельфинов при наблюдении за ними
в природе? Особенно если дельфины «стесняются» и не показываются из воды? Здесь
поможет знание о форме их плавников. У азовки – треугольник в виде домика, у афалины
– серповидный сильно загнутый, у белобочки – слегка загнутый серповидный, часто
со светло-серым пятном. А еще поведение дельфинов: афалины — грациозные и неторопливые,
белобочки – быстрые как ракеты, шустрые, азовки – самые стеснительные и пугливые.
Слайд 20. А теперь давайте проверим, сможете ли
вы узнать на фото дельфинов и китов, относящихся к разным видам. Зайдите в игру
по ссылке или с помощью QR- кода. Ну как, какие получились результаты. При
необходимости педагог проходит игру на демонстрационном экране вместе со всеми
учащимися.
Слайд 21. Обобщим полученные знания и проверим
себя с помощью увлекательной викторины «Кто хочет стать миллионером». Победители
получат миллион … аплодисментов! При необходимости педагог проходит игру на демонстрационном
экране вместе со всеми учащимися.
Слайд 22. Благодарим за внимание! Спрашиваем, какие
остались у ребят вопросы и отвечаем на них.
Информация для педагога:
Вы можете помочь нам в разработке занятий.
Для этого пройдите, пожалуйста по ссылке и ответьте на вопросы:
https://forms.gle/tnWk7LcW43v95o6dA
в условии
в решении
в тексте к заданию
в атрибутах
Категория:
Атрибут:
Всего: 50 1–20 | 21–40 | 41–50
Добавить в вариант
Найдите ошибки в приведённом тексте. Укажите номера предложений, в которых сделаны ошибки.
1. Окунь — первичноводное животное, кит — вторичноводное животное. 2. У первичноводных животных есть органы боковой линии, воспринимающие давление, направление движения, скорость течения воды. 3. Передние конечности кита превратились в грудные плавники 4. Задние конечности кита видоизменены в хвостовой плавник. 5. У окуня и кита жаберное дыхание. 6. У китов к крестцовому отделу позвоночника прикреплены тазовые кости.
Источник: ЕГЭ по биологии 14.06.2016. Основная волна. Вариант 50
Миоглобин — белок, запасающий кислород в клетках поперечнополосатой и сердечной мышечной ткани. В случае временной нехватки кислорода миоглобин высвобождает его, тем самым восполняя дефицит. Миоглобин является эволюционным предшественником гемоглобина, имеет большее сродство к кислороду, то есть эффективнее его связывает и хуже высвобождает. Сравните содержание миоглобина в мышцах синего кита и в мышцах китовой акулы. Сравните содержание миоглобина в мышцах синего кита и африканского слона. Сравните содержание миоглобина в грудных мышцах полевого воробья и домашней курицы. Ответ поясните.
Щука, крокодил, тюлень, кит имеют обтекаемую форму тела, так как они
1) дышат растворённым в воде кислородом
2) населяют сходную среду обитания
3) питаются подвижной добычей
4) относятся к одному типу
Источник: ЕГЭ по биологии 30.05.2013. Основная волна. Центр, Урал. Вариант 3.
Установите соответствие между примерами гомологичных и аналогичных органов: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ПРИМЕРЫ
А) плодолистики и прицветники
Б) ласт кита и крыло птицы
В) колючки барбариса и выросты стебля у ежевики
Г) листья и тычинки цветка
Д) глаз зайца и глаз пчелы
Е) крыло летучей мыши и крыло бабочки
ОРГАНЫ
1) гомологичные органы
2) аналогичные органы
Запишите в таблицу выбранные цифры под соответствующими буквами.
| А | Б | В | Г | Д | Е |
Установите последовательность расположения таксономических названий, начиная с наименьшего. Запишите в таблицу соответствующую последовательность цифр.
1) Дельфиновые
2) Млекопитающие
3) Зубатые киты
4) Китообразные
5) Хордовые
6) Дельфин−белобочка
Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Приспособления к жизни в воде, сформировавшиеся в процессе эволюции у китов:
1) превращение передних конечностей в ласты
2) дыхание кислородом, растворённым в воде
3) дыхание кислородом воздуха
4) обтекаемая форма тела
5) развитый подкожный слой жира
6) постоянная температура тела
Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Киты, как и другие млекопитающие:
1) дышат кислородом воздуха
2) дышат кислородом, растворённым в воде
3) имеют обтекаемую форму тела
4) имеют четырёхкамерное сердце
5) передвигаются с помощью ласт и хвостового плавника
6) обладают постоянной температурой тела и интенсивным обменом веществ
Установите соответствие между примером и морфофизиологической особенностью, которой соответствует данный пример.
ПРИМЕР
А) предплечье лягушки и курицы
Б) ноги мыши и крылья летучей мыши
В) крылья воробья и крылья саранчи
Г) плавник кита и плавник рака
Д) роющие конечности крота и медведки
Е) волосы человека и шерсть собаки
ОСОБЕННОСТЬ
1) гомологичные органы
2) аналогичные органы
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| A | Б | В | Г | Д | Е |
Установите соответствие между приспособленностью организмов и эволюционным процессом, в результате которого она сформировалась.
ПРИСПОСОБЛЕННОСТЬ
А) ласты кита и роющие конечности крота
Б) крылья птицы и крылья бабочки
В) обтекаемая форма тела дельфина и акулы
Г) разные формы клюва у вьюрков
Д) крылья летучей мыши и крылья совы
ЭВОЛЮЦИОННЫЙ ПРОЦЕСС
1) дивергенция
2) конвергенция
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| A | Б | В | Г | Д |
Источник: Демонстрационная версия ЕГЭ—2015 по биологии
В результате идиоадаптаций появилась(-лись)
1) способность к смене окраски хамелеона при опасности
2) хлоропласты и фотосинтез
3) ткани растений
4) ядовитые железы змей
5) ласты кита
6) первичная и вторичная полости тела у червей
Укажите пример ароморфоза у животных
1) ласты кита
2) копыто лошади
3) конечности лягушки
4) присоски и крючки ленточных червей
Прочитайте текст:
По внешнему виду тутовый шелкопряд – невзрачная бабочка с толстым, сильно волосистым телом и белыми крыльями, достигающими в размахе 4–6 см. Несмотря на наличие крыльев, бабочки в результате одомашнивания утратили способность летать. Родина шелкопряда, по всей вероятности, Китай, откуда, вместе с тутовым деревом, он распространился на север и на юг, но в дикой природе не встречается, разводят его в Южной Европе, Средней Азии, на Кавказе и в других регионах. Гусеницы питаются только листьями тутового дерева. Поэтому распространение шелководства связано с местами произрастания тутового дерева (шелковицы).
Какие критерия вида описаны в тексте? Ответ поясните.
Источник: ЕГЭ по биологии 14.06.2016. Основная волна. Вариант 2
Выберите три верных ответа из шести и запишите в ответ цифры, под которыми они указаны. Примером общей дегенерации служит
1) редукция органов чувств у ленточных червей
2) редукция задних конечностей у кита
3) отсутствие хлорофилла у растений-паразитов
4) отсутствие конечностей у змеи
5) отсутствие пищеварительной системы у бычьего цепня
6) потеря страусом способности к полёту
Установите соответствие между примерами и типами межвидового взаимодействия: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ПРИМЕР
А) Петров крест и многолетнее растение
Б) заяц и волк
В) клевер и корова
Г) бычий цепень и человек
Д) кит и рачки
Е) трутовый гриб и берёза
ТИП
1) хищничество (выедание)
2) паразит–хозяин
Запишите в таблицу выбранные цифры под соответствующими буквами.
| А | Б | В | Г | Д | Е |
Установите соответствие между примерами и направлениями эволюции: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ПРИМЕР
А) отсутствие пищеварительной системы у бычьего цепня
Б) обтекаемая форма тела у китов
В) появление теплокровности у птиц
Г) появление плотной оболочки яйца у рептилий
Д) редукция хорды у асцидий
НАПРАВЛЕНИЕ
1) ароморфоз
2) идиоадаптация
3) общая дегенерация
Запишите в таблицу выбранные цифры под соответствующими буквами.
| А | Б | В | Г | Д |
 Проанализируйте графики «Изменение экологического следа и биоёмкости
Проанализируйте графики «Изменение экологического следа и биоёмкости
сельскохозяйственных угодий одной из провинций Китая».
Экологический след – условная величина, характеризующая размер площади,
необходимой для обеспечения одного человека пищей, теплом и т. д.
в течение года.
Биоёмкость – условная величина, характеризующая площадь биологически
продуктивной территории, которая может использоваться для
удовлетворения потребностей человека в течение года.
Выберите утверждения, сформулированные на основании анализа
полученных результатов.
1) Наблюдается истощение сельскохозяйственных ресурсов в провинции.
2) Потребление пищи и других ресурсов населением провинции растёт.
3) Забота об экологической обстановке в провинции постепенно растёт.
4) Биоёмкость среды в провинции в целом снижается.
5) Максимальным экологический след был в провинции в 1999 г.
Запишите в ответе цифры, под которыми указаны выбранные утверждения.
Источник: ЕГЭ по биологии 2018. Досрочная волна
Установите соответствие между примерами и направлениями эволюции: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ПРИМЕР
А) отсутствие хлорофилла у растения
заразиха
Б) появление кровеносной системы
у древних кольчатых червей
В) видоизменение листочков в усики
у гороха
Г) видоизменение передних конечностей
в ласты у китов
Д) редукция пищеварительной системы
у ленточных червей
Е) появление цветов и плодов у покрыто-
семенных
1) ароморфоз
2) идиоадаптация
3) общая дегенерация
Запишите в таблицу выбранные цифры под соответствующими буквами.
| А | Б | В | Г | Д | Е |
Установите соответствие между примерами и сравнительно-анатомическими доказательствами эволюции: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ПРИМЕРЫ
А) крылья птиц и крылья стрекоз
Б) шипы розы и иголки кактуса
В) щупальца осьминога и гидры
Г) ласты кита и ноги лошади
Д) усик гороха и лист ромашки
1) аналогичные органы
2) гомологичные органы
Запишите в таблицу выбранные цифры под соответствующими буквами.
| А | Б | В | Г | Д |
Установите соответствие между примерами и сравнительно-анатомическими доказательствами эволюции: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ПРИМЕРЫ
А) крыло птицы и крыло бабочки
Б) нога лошади и ласт кита
В) хелицеры паука и антенны рака
Г) присоски морской звезды и кальмара
Д) глаза рыбы и кальмара
ДОКАЗАТЕЛЬСТВА ЭВОЛЮЦИИ
1) аналогичные органы
2) гомологичные органы
Запишите в таблицу выбранные цифры под соответствующими буквами.
| А | Б | В | Г | Д |
Раздел: Основы эволюционного учения
Проанализируйте диаграмму «Изменение площади урбанизированных земель в одном из городов Китая».
Выберите утверждения, которые можно сформулировать на основании
анализа полученных результатов.
1) площадь урбанизированных земель постоянно увеличивается
2) административная площадь города постоянно увеличивается
3) с 2004 по 2009 г. скорость роста урбанизированной площади увеличилась
4) население урбанизированных территорий постоянно увеличивается
5) нагрузка урбанизированных территорий на окружающую среду постоянно растёт
Источник: ЕГЭ по биологии 2020. Досрочная волна. Вариант 2
Всего: 50 1–20 | 21–40 | 41–50
Порой вернуться куда-то сложнее, чем начать все c чистого листа. Это верно и биологически: эволюция не идет вспять, так что приспособления к новым условиям жизни не развиваются из ниоткуда, а опираются на прошлые — порой диаметрально им противоположные. Четыреста миллионов лет назад кистеперые рыбы задышали воздухом, очутились на суше и стали амфибиями, которым вода нужна в гораздо меньшей степени, затем появились те, кто умеет размножаться вовсе без воды. Но полсотни миллионов лет назад некоторые из них начали путь обратно, в воду. Их потомки — современные китообразные — не выходят на сушу даже ради размножения. Между древними и нынешними китами — пропасть физиологических проблем, которые им пришлось частично параллельно, частично последовательно решать анатомическими адаптациями.
Китообразные — потомки парнокопытных, а по каким-то классификациям и вовсе одни из них. Ближайшие живые наземные родственники китов — бегемоты. Первые киты вроде пакицета (жил на территории современного Пакистана 49–48 миллионов лет назад) внешне были похожи на головастых собачек. Кстати, тогда среди копытных встречались и хищники — мезонихии.
Что сподвигло предков китообразных подобно своим прадедам рыбам вновь окунуться в воду, точно никто не знает. Может, там пряталась желанная добыча, а может, они сами служили для кого-нибудь обедом и пытались скрыться от недругов, как сейчас это
водяной оленек. В любом случае, китам приходилось то погружаться, то всплывать.
Проблема 1: надо глубоко нырнуть, а потом всплыть
Даже если (особенно если) животное не все время проводит в воде, чтобы доставать что-то с глубины или прятаться на ней, нужно уметь нырять. Этого не выйдет, если плотность тела ниже плотности воды. Ну а если она, напротив, существенно выше, первый нырок может стать последним. Потому что всплывать тоже надо уметь.
У современных китообразных плотность костей ниже, чем, скажем, у коз или оленей. Но пришли они к этому не сразу: сначала плотность костей, наоборот, повысилась. Их микроструктура начала меняться еще до того, как передние конечности стали походить на плавники, а задние редуцировались. Древние китообразные из группы археоцетов — пакицет, амбулоцет и подобные — 50–48 миллионов лет назад при дайвинге полагались на кости и мышцы. Их скелет стал плотнее, чем у наземных млекопитающих, чтобы им проще было уходить на дно. А всплывали и выходили на сушу они, по-видимому, за счет мускулов.
Пакицет плавал, загребая лапами. Он статично регулировал плотность тела, повысив ее за счет очень тяжелых костей, в которых были малоактивны остеокласты — клетки, прорежающие живое вещество кости. Но потом эволюция китов «передумала», и они начали использовать динамические механизмы контроля: уменьшение и увеличение объема легких. У дорудонов, живших 40–36 миллионов лет назад, плотность костей уже начала снижаться, а у современных китов она ниже, чем у наземных парнокопытных.
Мы знаем об этом благодаря гистологическим исследованиям ребер современных и ископаемых китообразных, а также некоторых других водных и наземных животных (последних использовали в качестве контроля).
«Воздушное» изменение плотности тела удобнее: чтобы держаться у поверхности, можно вдохнуть поглубже и стать за счет этого «легче» воды, и выдохнуть покрепче, дабы нырнуть, став «тяжелее». Также предполагают, что менять плотность тела некоторым китообразным вроде кашалота помогает еще и спермацет — та самая жировая подушка в голове, из которой еще недавно делали свечи. На глубине холоднее, и она твердеет и становится плотнее, а ближе к поверхности, где теплее, она «разжижается». Спермацет появился около 34 миллионов лет назад с одними из первых зубатых китов.
У продвинутых китов, которые уже хорошо ныряют глубоко и надолго, появляется другая проблема: декомпрессионная (кессонная) болезнь. Она возникает главным образом из-за того, что с изменением давления меняется растворимость азота в жидкостях тела: чем давление выше, тем она больше. Азот в организме ни на что не тратится, поэтому его не становится меньше, пока не выдохнешь. Из-за этого во время быстрого всплытия кровь в буквальном смысле вспенивается — это азот образует пузырьки, способные повредить стенки сосудов и другие важные структуры.
Чтобы такого не происходило, при погружении китообразные — не сознательно, конечно — максимально сужают просвет сосудов во всех органах, кроме мозга и сердца, которые надо постоянно «кормить» кислородом, тоже присутствующем в крови. В остальные части тела в этот момент кровь практически не поступает, зато и от пузырьков азота они защищены. От гипоксии китовые органы спасаются благодаря улучшенному миоглобину, эффективно сохраняющему кислород.
Если судить по заряду поверхности молекулы миоглобина, который во многом определяет способность этого вещества связываться с кислородом, получается, что одним из первых стал нырять базилозавр 45–36 миллионов лет назад. Но умел ли он так сужать сосуды, мы пока не знаем: у ископаемых китообразных не сохранилось столько мягких тканей, чтобы это понять.
Проблема 2: ничего не видно и не слышно
И акустический импеданс, и показатель преломления у воды не такие, как у воздуха. Поэтому с ушами и глазами, заточенными на работу в воздушной среде, под водой делать нечего. В те времена, когда китообразные высматривали в воде добычу с суши и потом ныряли за ней, «наземные» органы чувств еще можно было использовать, и то с трудом, но при переходе на полностью водный образ жизни — вряд ли.
Наземное ухо усиливает колебания воздуха (звуки) за счет системы полостей и мембран. Ушная раковина собирает звуковые волны, они попадают на барабанную перепонку, подхватываются вибрациями слуховых косточек и попадают на овальное окно, площадь которого меньше, чем у барабанной перепонки. Жидкость во всей этой цепочке встречается еще дальше, она служит проводником звука только в улитке внутреннего уха. Жидкость колеблется, эти колебания воспринимают погруженные в нее волосковые клетки. Разница импеданса воздуха и жидкости способствует восприятию звуков. Извлекать из нее выгоду тоже помогают мембраны — перепонки и «окна».
У рыб есть только внутреннее ухо, и улитка его тоже заполнена жидкостью. Тем не менее, чувствительные клетки в нем одним концом прикреплены к отолитам — «камешкам» плотнее воды, а кроме того, рядом с ними есть воздушные полости. Различия в колебаниях трех сред эффективно активируют волосковые клетки — видимо, одна жидкость бы так не смогла.
В воде ушные раковины, во-первых, очень мешают эргономично перемещаться, так что у всех, кто живет в воде, они редуцированы. Во-вторых, ухо в воде уже окружено жидкостью — и та будет в него заливаться, если не закрывать наружный слуховой проход. В таких условиях разница физических свойств сред не имеет большого значения, потому что среда фактически одна.
Поэтому у китообразных наружный слуховой проход
, ушная раковина пропала, а главный уловитель и усилитель звуков — жировые подушки в основании нижней челюсти, по одной с каждой стороны, а вместо барабанной перепонки — костная барабанная пластинка.
У пакицета, который под воду еще не «переехал», ухо было еще вполне наземным, преображение началось примерно 45 миллионов лет назад с ремингтоноцетид и протоцетид, а дорудоны и базилозавры уже имели «морское» строение органа слуха.
Говорить под водой тоже не слишком удобно, ведь при этом надо либо выдыхать, либо (так умеют не все) вдыхать. Поэтому зубатым китам пришлась особенно кстати эхолокация (а усатые к ней не способны, они издают «обыкновенные» звуки): кроме того, что она заменяет зрение на глубине и обоняние в воде, она позволяет кричать, не открывая рта. Тут, как и со слухом, опять помогает жир. Полголовы у кашалота занимает линза из жира переменной плотности — мелон. Она усиливает и максимально широко распространяет звуковые волны, которые появляются в воздушных мешках под дыхалом.
Глазам в воде тоже пришлось поменяться. Помимо того, что эта среда преломляет свет не так, как воздух, она еще и поглощает довольно большую его часть, из-за чего с глубиной быстро падает освещенность.
Высмотреть что-то в таких условиях сложно, но можно. Для этого надо изменить форму глаза: сделать роговицу плоской, а хрусталик круглым (у нас наоборот). Кроме того, капризные колбочки в сетчатке, которым для работы нужна хорошая освещенность, замещаются более неприхотливыми палочками. Пусть вторые не обеспечивают такой же цветопередачи, она и не особенно нужна в мутной воде.
Воздушно-жировая машинерия для восприятия и генерации звуков в ходе эволюции все увеличивалась, носовые отверстия все отодвигались от передней части черепа к задней, и места для обонятельной системы оставалось все меньше. Ее изменения задели и тех, кто не пользуется мелоном и прочими структурами для эхолокации. У всех китообразных редуцировано и число обонятельных рецепторов, и их многообразие, и объем обонятельных луковиц.
Все это привело к тому, что усатые и зубатые киты (последние особенно) почти ничего не чуют. Но все-таки начисто они обоняния не лишились: поведенческие исследования показывают, что некоторые дельфины способны различать запах рыбы и морепродуктов, а усатые киты — зоопланктона. Учитывая, что обоняние — это в первую очередь способность воспринимать пахучие вещества, содержащиеся в воздухе, потеря не критичная: все равно искать пищу им надо в воде, а не над водой. Соответственно, и со вкусом у китообразных не густо: ощущать его особенно нечем, даже генов соответствующих рецепторов (за исключением рецепторов соленого) нет.
Проблема 3: морской водой не напьешься
Полсотни миллионов лет назад древнейшие киты вроде пакицета плавали в пресной воде, но уже через пять миллионов лет перебрались в море. Это вычислили по содержанию изотопа кислорода-18 в зубах ископаемых китообразных: чем больше его в объекте, тем чаще он контактировал с морской водой. В зубах ремингтоноцетов содержится столько изотопа кислорода-18, что становится ясно: их чаще использовали в морской воде, чем на суше или в воде пресной. Современные китообразные почти все морские — за исключением речных дельфинов.
«Переезд» из пресной воды в соленую — непростое приключение для почек. Они выводят соединения азота и некоторые минеральные ионы, но не в твердом виде, а в составе водного раствора. Почкам пресноводных и обычных наземных зверей можно производить мочу с концентрацией хлорида натрия 0,9 процентов по массе. Хотя при выведении такого раствора потеряется довольно много H2O, нехватку последней можно быстро восполнить водой из реки, ручья или подобного источника.
В «усредненной» морской воде NaCl в четыре раза больше — 3,5 процента, то есть она заметно более концентрированная, чем моча. Поэтому, если пить только ее, избытки соли обычные почки вывести не смогут. Ее концентрация в жидкостях тела начнет повышаться, и это приведет к обезвоживанию.
Выход — делать такую мочу, чтобы концентрация солей в ней была выше, чем в морской воде. Это как раз умеют делать почки китообразных (а кроме них еще дюгоней, тюленей и каланов). По всей видимости, процессу способствует особая структура почек: они у китов не цельные, а скорее похожи на массивные грозди винограда, и это, как полагают, помогает увеличить общую длину и объем фильтрационных канальцев. Помогают, видимо, и гормоны вроде вазопрессина, но как именно, еще не очень ясно.
Тем не менее многие китообразные вообще избегают пить морскую воду либо пьют ее совсем понемногу. Они довольствуются той H2O, которая содержится в пище или получается в разных реакциях в клетках. Более того, китам приходится постоянно от нее избавляться, ведь при каждой попытке поесть она сама заливается в рот.
Современные усатые киты, заглатывая очередную порцию планктона, отфильтровывают воду через серии роговых пластин — собственно, «усы» (кстати, они уникальны и не гомологичны ни зубам, ни другим составляющим головы китов) — и выбрасывают ее в районе задней части нижней губы. Китовый ус существует уже примерно 34 миллиона лет и был у одного из древнейших усатых китов — этиоцета (но и зубы тот еще сохранил).
Зубатые киты, чья добыча нередко крупнее, могут всосать ее и потом как бы отгородить языком от воды, а затем выбросить воду через полуприоткрытые челюсти (не через дыхало!). Конечно, это удобнее делать в воздух: так новые порции не затекут обратно. Правда, для этого надо подняться к поверхности.
В любом случае, сначала животные избавляются от излишков воды насколько могут, а потом уже глотают добычу. Но даже то небольшое количество жидкости, от которого не удается избавиться, нельзя допускать в дыхательные пути. У косаток для этой цели
мышцы гортани — не дают жидкости заливаться в трахею.
Проблема 4: никогда нельзя расслабляться
Любому организму постоянно кто-то или что-то угрожает: хищники, бескормица, холод или жара. Но настолько враждебной среды, как та, в которой находятся китообразные, редко встретишь. Она не поддерживает один из важнейших процессов жизнедеятельности — нормальный газообмен. Кислорода легкие из нее не способны добыть: для такого нужны жабры. Поэтому приходится периодически всплывать на поверхность.
Да, время между вдохами можно увеличить, но задерживать дыхание бесконечно невозможно. Так что в спячку китообразным впадать нельзя (кстати, это, в свою очередь, заставляет постоянно поддерживать высокую интенсивность метаболизма). Спать без задних ног тоже рискованно: в состоянии, когда все чувства приглушены, можно пропустить момент, когда уже понадобится подняться за очередной порцией кислорода.
Китообразные, а также ластоногие, нашли выход — изменили структуру сна. Стадия быстрых движений глаз (REM-сон) у них на электроэнцефалограммах (ЭЭГ) никогда не проявляется, хотя ее поведенческие проявления, такие как эрекция, подергивания мышц и век, имеются.
Остался медленноволновой сон. Его регистрируют то в одном полушарии, то в другом. Получается, что мозг китов никогда не спит полностью. Это позволяет им сохранять постоянный контроль над дыхалом: по каким-то причинам китообразные, видимо, не могут подобно людям дышать «бессознательно». Интересно, что при однополушарном сне животное не обязательно закрывает глаз, которым руководит покоящаяся половина мозга.
Если учесть, что мы видим сны в основном во время REM-сна, получается, что китам ничего не снится. Еще есть предположения, что эта стадия необходима для повторения и укрепления воспоминаний (их перевода в долговременную память). Но китообразным ее отсутствие будто бы не мешает запоминать информацию: известно, что афалины узнают друг друга по голосам даже после двадцати лет разлуки.
Что сподвигло китообразных покинуть сушу, мы точно не знаем — но понятно, что процесс это был непростой и энергозатратный. Кроме перечисленных проблем, китам пришлось еще как минимум решать задачи терморегуляции и экономичного движения в воде.
В этом они, кстати, не уникальны. Кроме китов к полностью водному образу жизни перешли другие млекопитающие — сирены. У них приспособления немного иные хотя бы потому, что они гораздо чаще китообразных встречаются в пресной воде.
В преображении китов навряд ли стоит усматривать какой-то умысел. Маловероятно, что первые представители этой группы, еще четвероногие, собирались преодолевать колоссальные трудности и кардинально меняться. Был ли у китообразных выбор не уходить в воду, мы не знаем. Скорее всего, все шло своим чередом, адаптации появлялись как появляются — и в какой-то момент (не сразу, правда, а через десятки миллионов лет) на смену сухопутным «оленькам» и «мини-бегемотам» пришли гигантские океанические пожиратели криля и ныряющие на сотни метров борцы с кальмарами. В любом случае, приспособления китов показывают: кем бы ты ни был, при должном количестве времени можно кардинально измениться для новой жизни.
Светлана Ястребова

![]()
Современные виды отряда Китообразные.
Китообразные (киты, дельфины, морские свиньи) происходят от сухопутных предков, о чём свидетельствуют многие их признаки:
- дыхание атмосферным воздухом с помощью легких,
- кости плавников китообразных напоминают кости конечностей наземных млекопитающих,
- движение хвоста и изгибание позвоночника в вертикальной плоскости более характерно для бегущего млекопитающего, чем для плывущей рыбы, изгибающейся обычно в горизонтальной плоскости.
Вопрос о том, как наземные млекопитающие эволюционировали в морских, долгое время оставался загадкой из-за отсутствия ископаемых останков переходных видов. Однако благодаря открытию пакицетуса в 1992 году стало возможным прояснить ряд вопросов и увидеть стадии перехода наземных животных в морских.
Содержание
- 1 Предки китообразных
- 2 Самые ранние животные из семейства китовых: Pakicetids или Indohyus?
- 3 Амбулоцетиды и ремингтоноцетиды
- 4 Протоцетиды
- 5 Базилозавриды и дорудонтиды: полностью морские китообразные
- 6 Появление эхолокации
- 7 Ранние усатые киты
- 8 Ранние дельфины
- 9 Скелетное развитие
- 10 Примечания
- 11 Ссылки
Предки китообразных
![]()
Эволюционное древо китообразных (также включены копытные)
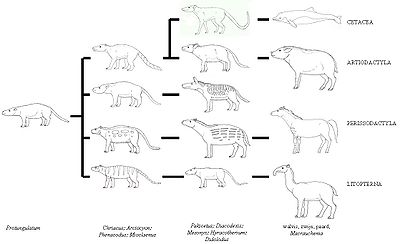
Традиционные взгляды на эволюцию китообразных состояли в том, что их ближайшими родственниками и, вероятно, предками являлись мезонихии — вымерший отряд хищных копытных, которые напоминали волков с копытцами вместо когтей и были сестринской группой парнокопытных. Эти животные имели зубы необычной конической формы, похожие на зубы китообразных. В частности из-за этого учёные долгое время считали, что китообразные произошли от некой предковой мезонихии. Однако новые молекулярно-генетические данные свидетельствуют о том, что китообразные — близкие родственники парнокопытных, в частности, гиппопотамов[1]. На основе этих данных предлагается даже включать китообразных в состав отряда парнокопытные и предлагается название Cetartiodactyla для монофилетического таксона, включающего две эти группы. Однако наибольший возраст известных ископаемых остатков антракотериев, предков бегемотов, на несколько миллионов лет меньше, чем возраст Pakicetus, наиболее древнего из известных предков китов.
Недавнее открытие рода Pakicetus, наиболее древнего из известных «прото-китоообразных», подтверждает молекулярные данные. Строение скелета Pakicetus показывает, что киты не являются прямыми потомками мезонихид. Напротив, предки китов отделились от парнокопытных и перешли к водному образу жизни уже после того, как сами парнокопытные отделились от общих с мезонихидами предков. Таким образом, прото-китообразные были ранними формами парнокопытных, которые сохранили некоторые характерные для мезонихид признаки (коническую форму зубов), утраченные современными парнокопытными. Интересно, что самые ранние предки всех копытных млекопитающих были, вероятно, частично плотоядными или «мусорщиками».
Самые ранние животные из семейства китовых: Pakicetids или Indohyus?

Пакицетиды были копытными животными, иногда их классифицируют как ранних китов[2][3]. Они жили на территории современного Пакистана (откуда и название — «кит из пакистана») в раннем эоцене, около 50 млн лет назад. Это было животное, внешне напоминавшее собаку, однако с копытами на пальцах и с длинным тонким хвостом. С китами пакицетуса роднит устройство уха: слуховая булла пакицетуса, как и у кита, образовалась исключительно из барабанной кости. Форма ушной области пакицетуса весьма необычна и находит аналоги только у китообразных. Первоначально предполагалось, что ухо пакицетуса было приспособлено для жизни под водой, однако, как показали дальнейшие исследования, уши пакицетуса подходят только для воздушной среды, и если пакицетус действительно предок китов, то возможность слышать под водой была позднейшей адаптацией уже имевшегося слухового аппарата[4]. Как утверждает Тевиссен, зубы пакицетуса также напоминают зубы ископаемых китов[5].

Тевиссеном было также обнаружено, что схожее устройство уха наблюдалось в окаменелостях небольшого, похожего на оленя животного индохиуса. Индохиус обитал около 48 млн лет назад в Кашмире[3]. Это небольшое — размером с домашнюю кошку — травоядное животное обладало некоторыми чертами, сближающими его с китами и свидетельствующими об адаптации к водной среде. В их числе — толстая и тяжёлая костная оболочка, напоминающая костную оболочку некоторых современных полуводных животных, таких как гиппопотамы[6][1], что способствует уменьшению плавучести и как следствие, позволяет оставаться под водой. Это позволяет предположить, что индохиус, подобно современному водяному оленьку, нырял под воду, чтобы укрыться от хищника[7][8][9].
Амбулоцетиды и ремингтоноцетиды


![]()
Ambulocetus natans. Реконструкция
Наиболее примечательным из древних китов является Амбулоцетус, известный из эоцена Пакистана. Внешне это млекопитающее походило на трёхметрового крокодила. Амбулоцетус был полуводным животным: его задние лапы лучше приспособлены для плавания, нежели для ходьбы по суше. Вероятно, он плавал, изгибая тело в вертикальной плоскости, как современные выдры, тюлени и киты. Предполагается, что амбулоцетиды охотились подобно современным крокодилам, подстерегая в засаде рыб и пришедших на водопой животных.
Близкими родственниками амбулоцетуса были ремингтоноцетиды. Представители этого семейства были мельче по размеру, имели более удлинённую морду и были лучше приспособлены к подводной жизни. Предполагается, что образом жизни они напоминали современных выдр, охотясь из засады на рыб.
У представителей обеих групп ноздри располагались на конце морды, как у наземных млекопитающих.
Протоцетиды

![]()
Протоцетус. Реконструкция

![]()
Родоцетус. Реконструкция

Протоцетиды образуют крупную и разнообразную группу, известную по находкам в Азии, Европе, Африке, и Северной Америке. Данное семейство включает в себя большое количество родов, некоторые из них довольно хорошо изучены (например, родоцетус, известный из третичных отложений Белуджистана). Все известные протоцетиды обладали хорошо развитыми передними и задними конечностями, которые могли поддерживать тело на земле; вероятно, они вели амфибиотический образ жизни, обитая как в водной среде, так и на суше. Пока не ясно, имелся ли у протоцетид хвостовой плавник, как у современных китообразных, однако очевидно, что они были неплохо приспособлены к водному образу жизни. Например, крестец — часть позвоночника, к которой крепится таз — у родоцетуса состоял из пяти раздельных позвонков, в то время как позвонки в крестце наземных млекопитающих слиты. Носовые отверстия сдвинулись у протокетид вверх по рылу — это является первым шагом к расположенным на макушке ноздрям нынешних китообразных. Версия об амфибийной природе протоцетид подкреплена находкой беременной самки майацетуса[10] с окаменевшим плодом, повёрнутым головой к выходному отверстию. Это заставляет предположить, что роды у майацетуса проходили на суше — в противном случае детёныш имел шансы захлебнуться.
О происхождении ранних китов от копытных говорят такие особенности, как, например, наличие копыт на концах пальцев у родоцетуса.
Базилозавриды и дорудонтиды: полностью морские китообразные

![]()
Базилозавр. Реконструкция
Базилозавр (обнаруженный в 1840 г. и первоначально принятый за рептилию, чем объясняется «рептильное» имя) и дорудон жили приблизительно 38 миллионов лет назад и представляли собой чисто морских животных. Базилозавр был столь же велик, как крупные современные киты, достигая порой 18 метров в длину. Дорудонтиды были несколько меньше, до 5 метров.

![]()
Дорудон. Реконструкция
При всём сходстве с современными китами, у базилозаврид и дорудонтид отсутствовал лобно-жировой выступ, так называемая «дыня», позволяющая ныне существующим китообразным эффективно использовать эхолокацию. Мозг базилозаврид был сравнительно небольшого размера, из чего можно предположить, что они вели одиночный образ жизни и не имели такой сложной социальной структуры, как у некоторых современных китообразных. В связи с переходом к чисто водному образу жизни у базилозаврид наблюдается деградация задних конечностей — они, хотя и хорошо сформированы, но невелики и уже не могут использоваться для передвижения. Впрочем, возможно, они играли вспомогательную роль при спаривании. Тазовые кости базилозаврид уже не связаны с позвоночником, как это было у протоцетид.
Появление эхолокации

![]()
Реконструкция Squalodon’а
Зубатые киты (Odontocetes) осуществляют эхолокацию, создавая серию щелчков на различных частотах. Звуковые импульсы излучаются посредством лобной дыни, отражаются от объекта и регистрируются с помощью нижней челюсти. Изучение черепов Squalodon позволяет предположить первичное возникновение эхолокации именно у этого вида. Squalodon жил с начала среднего олигоцена до середины миоцена, около 33—14 миллионов лет назад, и имел ряд признаков, сходных с современными зубатыми китами. Так, например, сильно сплюснутый череп и вынесенные челюстные дуги наиболее характерны для современных Odontoceti. Несмотря на это, возможность происхождения современных дельфинов от Squalodon считается маловероятной.
Ранние усатые киты

![]()
Реконструкция Cetotherium’a
Ранние дельфины

![]()
Скелет Xiphiacetus-а.
См. также «Дельфины, Развитие и анатомия (англ.)»
Скелетное развитие
Примечания
- ↑ 1 2 University Of California, Berkeley (2005, February 7) UC Berkeley, French Scientists Find Missing Link Between The Whale And Its Closest Relative, The Hippo. ScienceDaily. Архивировано из первоисточника 24 августа 2011. Проверено 1 февраля 2010.
- ↑ Philip D. Gingerich, D. E. Russell (1981). «Pakicetus inachus, a new archaeocete (Mammalia, Cetacea) from the early-middle Eocene Kuldana Formation of Kohat (Pakistan)». Univ. Mich. Contr. Mus. Paleont 25: 235–246.
- ↑ 1 2 Northeastern Ohio Universities Colleges of Medicine and Pharmacy (2007, December 21) Whales Descended From Tiny Deer-like Ancestors. ScienceDaily. Архивировано из первоисточника 24 августа 2011. Проверено 21 декабря 2007.
- ↑ J. G. M. Thewissen, E. M. Williams, L. J. Roe and S. T. Hussain (2001). «Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls». Nature 413: 277–281. DOI:10.1038/35095005.
- ↑ Whale Origins
- ↑ University Of Michigan (2001, September 20) New Fossils Suggest Whales And Hippos Are Close Kin. ScienceDaily. Архивировано из первоисточника 4 апреля 2012. Проверено 21 декабря 2007.
- ↑ Carl Zimmer The Loom : Whales: From So Humble A Beginning…. ScienceBlogs (19 декабря 2007). Архивировано из первоисточника 4 апреля 2012. Проверено 21 декабря 2007.
- ↑ Ian Sample Whales may be descended from a small deer-like animal — Science — Guardian Unlimited. Guardian Unlimited (19 декабря 2007). Архивировано из первоисточника 4 апреля 2012. Проверено 21 декабря 2007.
- ↑ PZ Myers Pharyngula: Indohyus. Pharyngula. ScienceBlogs (19 декабря 2007). Архивировано из первоисточника 4 апреля 2012. Проверено 21 декабря 2007.
- ↑ Gingerich PD, ul-Haq M, von Koenigswald W, Sanders WJ, Smith BH, et al. New Protocetid Whale from the Middle Eocene of Pakistan: Birth on Land, Precocial Development, and Sexual Dimorphism. PLoS one. Архивировано из первоисточника 4 апреля 2012. Проверено 4 февраля 2009.
Ссылки
- Cetacean Paleobiology — University of Bristol
- BBC: Whale’s evolution
- Hooking Leviathan by Its Past by Stephen Jay Gould
- Whale Origins, Thewissen Lab, Northeastern Ohio Universities College of Medicine
- Digital Library of Dolphin Development, Thewissen Lab
- Research on the Origin and Early Evolution of Whales (Cetacea), Gingerich, P.D., University of Michigan
- Evolution of Whales Adapted from National Geographic, November 2001, Revised 2006 Dr. J.G.M. Thewissen
- Pakicetus inachus, a new archaeocete (Mammalia, Cetacea) from the early-middle Eocene Kuldana Formation of Kohat (Pakistan). Gingerich, P.D., 1981, Museum of Paleontology, The University of Michigan
- Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls, Nature 413, 277—281 (20 September 2001), J. G. M. Thewissen, E. M. Williams, L. J. Roe and S. T. Hussain
- Evolution of Whales segment from the Whales Tohorā Exhibition Minisite of the Museum of New Zealand Te Papa Tongarewa
- журнал membrana: Первые киты с ногами определили стиль плавания всех китов
Алексей Лопатин
«Природа» №5, 2018
Об авторе

Алексей Владимирович Лопатин — академик РАН, доктор биологических наук, директор Палеонтологического института им. А. А. Борисяка РАН, профессор кафедры палеонтологии геологического факультета Московского государственного университета имени М. В. Ломоносова. Область научных интересов — морфология, филогения и эволюция позвоночных животных, биостратиграфия, палеобиогеография, палеоэкология.
Покорив 375 млн лет назад сушу, позвоночные неоднократно возвращались в море, осваивая неисчерпаемые запасы рыбы и других морских организмов. Некоторые из них, например морские черепахи и ластоногие, сохранили связь с сушей, другие же полностью перешли к водному образу жизни. Хорошо известны мезозойские морские ящеры — плезиозавры, плиозавры и мозазавры. Самые большие из них достигали почти 20 м в длину. По-видимому, они охотились на своих более мелких собратьев. Хищниками были и мезозойские морские крокодилы — искусные пловцы, имевшие плавники. Но самыми успешными подводными охотниками стали ихтиозавры и китообразные, которые независимо друг от друга в совершенстве приспособились к обитанию в открытых морских водах, уподобившись рыбам формой тела и научившись размножаться без выхода на сушу.
Переход к жизни в воде сопровождался сходными преобразованиями в строении тела. Его форма становилась либо обтекаемой, либо змеевидной, уплощенной с боков. Если основным органом движения становился хвост, он приобретал вид лопасти, а задние конечности сильно уменьшались или исчезали. Передние конечности превращались в ласты или плавники, служащие веслами, рулями глубины, средствами поворота и торможения. Зубные ряды составляли двойной острый частокол, способный удерживать и раздирать скользкую добычу…
После вымирания гигантских водных рептилий мезозоя ниша морских суперхищников долго оставалась свободной и лишь частично осваивалась большими акулами, крокодилами и морскими змеями. Но около 55 млн лет назад появилась группа млекопитающих, со временем достигшая господства в морях кайнозойской эры, — китообразные…
Родственные узы
Китообразные — потомки наземных млекопитающих, и наследие их сухопутного прошлого проявляется в дыхании атмосферным воздухом, наличии плаценты, вскармливании детенышей молоком, а также в деталях строения скелета. В отличие от рыб, у которых при плавании позвоночник изгибается в горизонтальной плоскости, у китообразных он двигается в плоскости вертикальной. Современные киты и дельфины не могут жить на суше и выглядят очень необычно по сравнению с наземными млекопитающими. Вопрос о том, каким образом наземные звери превратились в морских чудовищ, долгое время оставался без ответа, так как в палеонтологической летописи отсутствовали соответствующие связующие звенья. В настоящее время картина в значительной мере прояснилась.
Предками китообразных до 2000-х годов считались мезонихии, родственные парнокопытным вымершие хищники, обликом отдаленно напоминавшие массивных собак с копытцами вместо когтей. Мезонихии были величиной от ласки до медведя, среди них известны и полуводные рыбоядные формы, такие как гапалодекты (Hapalodectes). Однако большинство мезонихий были наземными хищниками и падальщиками. Строение черепа и зубов этих древних млекопитающих (они жили 63–33 млн лет назад) долгое время давало основание полагать, что китообразные произошли непосредственно от мезонихий.
С этой гипотезой вступили в противоречие молекулярно-генетические данные, показывающие, что китообразные должны рассматриваться в пределах группы парнокопытных в качестве ближайших родственников бегемотов. На этом основании парнокопытные и китообразные иногда объединяются в группу «китопарнокопытных» (Cetartiodactyla) [1].

Открытие полного скелета наземного китообразного пакицета (Pakicetus attocki) поддержало молекулярно-генетические данные, показав, что китообразные отделились от древних парнокопытных уже после того, как те разошлись с мезонихиями [2]. Иными словами, предками китов были примитивные парнокопытные, сохранившие признаки их общего с мезонихиями происхождения (например, в строении зубов), которые современные парнокопытные давно утратили. Это были всеядные и плотоядные животные. Парнокопытные позже перешли к питанию растительным кормом, но некоторые из них (например, свиньи) и ныне в значительном объеме потребляют животную пищу. Киты — водные теплокровные, нуждающиеся в высококалорийной пище, поэтому они сохранили и развили животноядный характер питания своих предков.
В пользу этой гипотезы может свидетельствовать строение скелета индохиуса (Indohyus), который был найден в Кашмире (Индия) в отложениях возрастом около 48 млн лет [3]. Это животное относится к отряду парнокопытных, к вымершему семейству раоэллид. Телосложением и величиной оно напоминало вовсе не китов, а современных оленьков семейства трагулид, живущих в Западной Африке и Юго-Восточной Азии в густых лесах, кустарниковых и мангровых зарослях, умеющих хорошо плавать и нырять. Но по строению слуховой области черепа (это очень важный признак для выяснения родства среди млекопитающих) индохиус сближается лишь с одними древнейшими китами семейства пакицетид. Индохиус жил на несколько миллионов лет позже пакицетид и не может быть их предком — но он вполне может быть потомком одной из линий, родственной предкам династии морских великанов.
Основатели династии
Древние китообразные эволюционировали быстро, и переход от полуназемных форм к постоянноводным занял приблизительно 8 млн лет. Примитивных эоценовых китообразных, называемых археоцетами (древними китами), разделяют на пять семейств — это пакицетиды, амбулоцетиды, ремингтоноцетиды, протоцетиды и базилозавриды. В 2001 г. стали известны первые анатомически полные скелеты археоцетов, позволившие в деталях выяснить процесс перехода китообразных к жизни в море [2].
Пакицетиды жили в раннем эоцене (около 52 млн лет назад) на северном побережье субтропического внутриконтинентального моря Тетис, в те времена пересекавшего Евразию в широтном направлении. Это были большеголовые и длинномордые звери с копытцами на пальцах и длинными хвостами, пакицет и налацет (Nalacetus ratimitus) размерами соответствовали волку, а ихтиолест (Ichthyolestes pinfoldi) — лисице. Судя по строению скелета, пакицетиды хорошо плавали, используя волнообразные движения тела и гребки конечностями. По строению слуховой области черепа они сближаются с китами. Однако уши пакицетид были приспособлены к восприятию звуков в воздушной, а не в водной среде. Следовательно, эти животные жили на суше, добывая пищу в мелководных пресных водоемах и поблизости от них, подобно современным тапирам. Скелеты пакицетид были найдены в Пакистане и Северо-Западной Индии, в речных отложениях. Видимо, именно эти места были центром происхождения китообразных.
Данные по эволюции системы полукружных каналов внутреннего уха, участвующих в регуляции равновесия и положения тела в пространстве и ответственных за контроль локомоции, свидетельствуют, что пакицетиды в этом отношении еще были близки наземным млекопитающим [4].
Киты-амфибии
Одной из самых значимых находок древних китообразных, «недостающим звеном» в их эволюции, стал амбулоцет (Ambulocetus natans). Его название, означающее «плавающий ходячий кит», говорит само за себя. Почти полный скелет этого животного найден на севере Пакистана в отложениях возрастом около 49 млн лет назад [5]. Обликом трехметровый амбулоцет напоминал большеголового крокодила. При плавании его тело изгибалось в вертикальной плоскости, как у современных китов, тюленей и выдр. Предполагают, что амбулоцеты охотились подобно крокодилам, подстерегая своих жертв на мелководье (активно передвигаться по суше они не могли). Амбулоцеты имели мощные челюсти и зубы и были способны умертвить достаточно крупную добычу. Строение носа позволяло «киту-амфибии» глотать пищу прямо в воде. Глаза давали только боковой обзор. Ушные раковины амбулоцет, судя по всему, утратил, но хорошо слышал в воде. Движение своей добычи по суше он отслеживал, прижимая голову к грунту и улавливая колебания субстрата. Химический анализ зубов показал, что зверь мог охотиться как в солоноводных, так и в пресных водоемах. К семейству амбулоцетид относятся еще два рода: гандаказия (Gandakasia) и гималайяцет (Himalayacetus). Последний примечателен своей древностью, наибольшей среди китообразных — 53,5 млн лет.

Более мелкими родичами амбулоцетид были ремингтоноцетиды, известные из Индии и Пакистана, из отложений возрастом 49–43 млн лет назад. Эти животные обладали сильно вытянутыми челюстями и были несколько лучше приспособлены к подводному плаванию, чем амбулоцетиды. Они тоже имели хорошо развитые конечности, но отличались маленькими глазами, тонкими челюстями и расширенным основанием черепа. Если сравнивать «земноводных китов» по общему облику с крокодилами, то среди них амбулоцетиды больше напоминали аллигаторов, а ремингтоноцетиды — гавиалов. Строение слуховой области в сочетании с маленькими глазами показывает, что для обнаружения добычи ремингтоноцетиды пользовались в основном слухом. Известно шесть родов ремингтоноцетид. Их размеры были невелики: например, качхицет (Kutchicetus minimus) не превосходил величиной речную выдру. Для эндрюсифия (Andrewsiphius sloani) установлено наличие мощного уплощенного хвоста, служившего основным органом движения в водной среде [6]. Райанист (Rayanistes afer), найденный в среднем эоцене Египта [7], демонстрирует, что ремингтоноцетиды достигли южного побережья Тетиса.
Киты дальнего плавания
Первыми с мелководья на настоящую глубину отправились протоцетиды, жившие 49–37 млн лет назад. Их главным эволюционным приобретением был удлиненный мощный хвост (поначалу без хвостовой лопасти), который обеспечивал быстрое плавание [8]. При этом протоцетиды сохраняли развитые задние конечности. Вероятнее всего, по образу жизни их можно сопоставить с современными крупными ластоногими. У некоторых протоцетид зубы были уменьшены, это свидетельствует, что добычу — мелких морских животных — они заглатывали целиком. Такими были первые китообразные, которые сумели распространиться за пределы моря Тетис — их остатки найдены в мелководно-морских отложениях в Африке, Европе и Северной Америке. Самые крупные из них достигали длины более 3 м и весили до 400–500 кг. Вероятно, от протоцетид произошли все более поздние китообразные, включая современные группы. Описано около 20 родов протоцетид, из них наиболее известны протоцет (Protocetus) и родхоцет (Rodhocetus). Своеобразно выглядел макарацет (Makaracetus bidens), у которого (судя по строению передней части черепа) был короткий хоботок [9] или мясистая верхняя губа, которые могли служить для сбора со дна моллюсков и других животных.
Важные сведения о биологии протоцетид были получены на основе изучения майяцета (Maiacetus inuus). В Пакистане, в отложениях возрастом 47,5 млн лет, был обнаружен скелет самки, внутри которого сохранились череп и часть скелета эмбриона [10]. Его положение (вперед головой, а не хвостом, т.е. так, как у наземных млекопитающих) свидетельствует, что протоцетиды рождали детенышей на суше, а не в воде. Самцы майяцетов были крупнее самок и имели более мощные клыки. Такой половой диморфизм характерен для многих современных ластоногих, в частности моржей и морских львов. Возможно, он отражает сходный, «гаремный», тип социальной организации у древних китообразных.
Властелины древних морей

Первые действительно гигантские киты — базилозавриды — жили в позднем эоцене, приблизительно 41–35 млн лет назад. Их ископаемые остатки в основном происходят с востока США и из Египта, но, скорее всего, эти животные были распространены по всему свету [11]. Известно около 15 родов этих археоцетов. Представители подсемейства базилозаврин были огромны: их змеевидное тело массой до 6 т достигало в длину 15–20 м. У них имелся хвостовой плавник, но был ли он их основным органом движения — неизвестно. Как и для прочих древних китов, для базилозаврин были характерны зазубренные коренные зубы и конические предкоренные. Дорудонтины внешне походили на дельфинов (хотя имели заметную шею), при весе в 1,5 т достигали в длину до 6 м. Все базилозавриды имели вполне оформленные задние конечности с подвижным коленным суставом и несколькими пальцами. Однако эти конечности были совсем маленькими и явно не функционировали как органы движения. Возможно, самцы использовали их при спаривании для захватывания самок, как делают своими рудиментарными задними конечностями удавы. Несмотря на внешнее сходство с современными китообразными, базилозавриды не были способны к ультразвуковой эхолокации и акустической коммуникации. Относительно маленький мозг указывает на отсутствие у них сложного социального поведения, характерного для современных китов и дельфинов. В то же время явно выраженная асимметрия черепа базилозаврид была частью комплекса приспособлений, связанного с развитием у них направленного слуха для различения высокочастотных звуков, которые производили рыбы.
Самый крупный археоцет (и самое крупное млекопитающее своего времени) — это базилозавр (Basilosaurus cetoides). Он был описан еще в 1839 г. из Алабамы (США). Другие виды найдены в Египте и Пакистане. Строение позвоночника базилозавра позволяет предполагать, что при плавании он мог змеевидно извиваться. Скорее всего, базилозавр не был способен к длительному плаванию и глубокому нырянию. Не мог и выходить на сушу. Вероятно, он охотился на крупную добычу (например, акул, а также дорудонтин) у водной поверхности недалеко от побережья. В 2015 г. в Египте нашли полный скелет 18-метрового базилозавра с остатками другого кита внутри.

Между зубом и усом
У современных китов задние конечности рудиментарны — они скрыты внутри тела и служат для крепления мышц половых органов и лишь поэтому не исчезли полностью. Иногда рождаются киты с атавистическими миниатюрными задними конечностями. Вместо ноздрей современные киты имеют дыхало специфичного строения, смещенное на макушку. Строение ушной системы обеспечивает способность к акустической коммуникации. Современные группы китообразных — зубатые (Odontoceti) и усатые (Mysticeti) киты — появилсь в позднем эоцене, около 40 млн лет назад. К раннему олигоцену они стали обычны, а в позднем олигоцене (27–23 млн лет назад) дали мощную вспышку разнообразия, которая привела к возникновению большинства современных семейств. Резкое увеличение головного мозга у зубатых китов отмечается дважды: первоначально у самых ранних форм 40 млн лет назад и затем у дельфинов 15 млн лет назад [12].
Подотряд зубатых китов включает дельфинов, нарвалов, морских свиней и кашалотов. Большинство из них живут в океане (хотя есть речные дельфины), известны прибрежные и океанические виды, полярные и тропические. Все они активные хищники, добывают рыбу и кальмаров или, как косатки, охотятся на других китообразных, а также на пингвинов и ластоногих. В отличие от современных усатых китов, зубатые имеют зубы и способны к эхолокации. Каждое из современных семейств подотряда обладает собственной специализацией. Не менее разнообразны были и вымершие группы.
Современные усатые киты утратили зубы, но зато приобрели китовый ус, служащий им для фильтрации больших объемов мелкой добычи. Среди усатых китов — крупнейшее создание мира животных за всю его историю — синий кит.
Некоторые необычные представители как зубатых, так и усатых китов, в прошлом населявшие моря нашей планеты, заслуживают более близкого знакомства.
Мечерылые, акулозубые, моржеголовые…
Самые ранние черепные признаки, связанные с обладанием эхолокацией, обнаружены у олигоценового зубатого кита котилокары (Cotylocara macei) из вымершего семейства ксенорофид, который жил у побережья Южной Каролины (США) 28 млн лет назад [13]. Для позднего олигоцена и миоцена характерно высокое разнообразие архаичных групп зубатых китов, имевших пильчатые зубы, скорее похожие на археоцетовые, чем на более простые конические одонтоцетовые. У эвринодельфиса (Eurhinodelphis) из семейства эвринодельфинид беззубый конец верхней челюсти превратился в длинный отросток, служивший смертоносным оружием, как у меч-рыбы. Этот «мечерылый дельфин» был небольшим (около 2 м), но родственный ему макродельфин (Macrodelphinus kelloggi) достигал размеров косатки (6–8 м). Представители рода сквалодонов (Squalodon) из семейства акулозубых дельфинов (Squalodontidae) имели длинные узкие челюсти. Многочисленные виды семейства кентриодонтид (около 20 родов) внешним видом весьма напоминали современных дельфинов (Delphinidae), появившихся около 15 млн лет назад.

Симоцет (Simocetus rayi) из верхнего олигоцена Орегона (США), выделенный в самостоятельное семейство симоцетид, получил прозвание «курносый дельфин». Его короткие челюсти были заметно загнуты вниз, а редко расположенные зубы служили скорее для просеивания пищи, чем для ее разрывания. Вероятно, симоцет добывал донных беспозвоночных, процеживая осадок [14].
У двухметровой морской свиньи семиростра (Semirostrum ceruttii), жившей 3 млн лет назад у берегов южной Калифорнии (США), нижняя челюсть была вдвое длиннее верхней. Эта часть челюсти формировала покрытый кожей выступ-подбородок, который использовался как специальный чувствительный орган для поиска пищи «на ощупь» в мутной воде у самого дна [15].
История группы речных дельфинов (Platanistoidea) прослеживается с позднего олигоцена, когда ее представители еще обитали в морских бассейнах. При этом оказывается, что они вовсе не близкородственны друг другу. Слепой гангский дельфин (Platanista) принадлежит линии архаичных зубатых китов, родственной сквалодонтидам, тогда как другие — амазонский (Inia), лаплатский (Pontoporia) и озерный (Lipotes) — скорее всего, происходят от различных вымерших групп [16].
Самый необычный облик среди вымерших зубатых китов имел одобеноцетопс из плиоцена Перу (возраст 4–3 млн лет назад), названный так за внешнее сходство с современным моржом (Odobenocetops можно перевести как ‘моржеголовый кит’). Это небольшой дельфин, выделенный в собственное семейство одобеноцетопсид, родственник нарвалов и морских свиней. В противоположность большинству дельфинов, у одобеноцетопса не было удлиненного рыла, ноздри сместились вперед, а глаза — к передне-верхней стороне черепа, обеспечивая бинокулярное зрение. В верхней челюсти имелись два бивня, у самок — очень маленькие, а у самцов разные — левый почти как у самок (длина 5 см), а правый длинный, копьевидный, выступал наружу больше чем на 1 м (при том, что тело животного было чуть более 2 м). При плавании голова животного опускалась вниз, а бивень располагался почти параллельно телу и своим концом был направлен назад. Предполагается, что, подобно моржу, одобеноцетопс питался донными беспозвоночными, собирая их большой мясистой верхней губой с чувствительными вибриссами. Бивни, скорее всего, не использовались для добывания пищи, а служили самцам брачным оружием [17].

Кашалоты-убийцы и сам Левиафан
В раннем миоцене, около 20 млн лет назад, появились кашалоты (Physeteroidea). Ныне это специализированные охотники на кальмаров. Однако некоторые неогеновые кашалоты были китами-убийцами своего времени, занимая экологическую нишу косаток до их появления около 3 млн лет назад. Наименование «кашалота-убийцы» в научной литературе получил зигофизетер (Zygophyseter varolai) из позднего миоцена Италии, возрастом 10–7 млн лет назад [18]. Этот кит был длиной 6–7 м, с мощным черепом и многочисленными большими зубами в верхней и нижней челюстях. Грозными хищниками были и другие кашалоты-убийцы: семиметровый бригмофизетер (Brygmophyseter shigensis) из Японии (возраст 15–14 млн лет назад) и представители рода акрофизетеров (Acrophyseter) из Перу (9–6 млн лет), достигавшие в длину лишь 4 м, но также занимавшие нишу активных морских охотников [19].
Остатки намного более крупного морского хищника были также обнаружены в Перу, их возраст 10–9 млн лет назад [19, 20]. В длину гигантский кашалот-убийца достигал 18 м, а весил он около 40 т (примерно как самцы современного кашалота). Длина его черепа около 3 м, а зубы имеют диаметр около 12 см и длину до 36 см. Пожалуй, это самые крупные зубы в животном мире, не считая гипертрофированных клыков и бивней. Колосс получил название «мелвиллов левиафан» (Livyatan melvillei), в честь библейского чудовища Левиафана и американского писателя Германа Мелвилла, автора романа «Моби Дик, или Белый кит». Можно предполагать, что появление таких крупных хищников было связано с развитием гигантизма среди их потенциальных жертв — усатых китов.
Киты-леопарды и беззубые великаны
Усатые киты произошли от археоцетов, по-видимому, в высоких широтах. Известны олигоценовые группы, которые имели широко расставленные зубы, а в строении черепа совмещали черты, свойственные археоцетам и усатым китам. Очень необычен зубастый янджуцет (Janjucetus hunderi) из позднего олигоцена Австралии, длиной около 3,5 м, имевший исключительно большие глаза и укороченную морду. Предполагается, что он был активным хищником, охотившимся на крупную рыбу, включая акул, и экологически напоминал морского леопарда — современного хищного тюленя [21]. Янджуцет относится к семейству маммалодонтид. Другие представители этой вымершей группы, маммалодоны (Mammalodon) из олигоцена Австралии и Новой Зеландии, имели редко посаженные зубы. Вероятно, они питались донными беспозвоночными, процеживая ил [22]. Их открытие подтвердило догадку Чарлза Дарвина, который предположил в «Происхождении видов», что фильтрующие усатые киты произошли именно от таких форм — Дарвин называл их «просеивателями».

В середине олигоцена, около 30 млн лет назад, появились фильтрующие усатые киты. У представителей вымершего семейства эомистицетид (Eomysticetus, Waharoa и др.), сравнительно некрупных (до 8 м) китов стройного сложения, китовый ус занимал только заднюю часть неба, на переднем конце челюсти имелось несколько зубов, а дыхало располагалось на значительном расстоянии спереди от глаз. Древнейшие находки гладких китов (Balaenidae) имеют возраст 23 млн лет. Полосатиковые (Balaenopteridae), к которым относится гигантский синий кит, появились в Северной Атлантике около 12 млн лет назад. Судя по ископаемым остаткам, современному синему киту по размерам не уступали некоторые киты-полосатики плиоцена и плейстоцена, например Balaenoptera sibbaldina из раннего плиоцена Бельгии [23]. Однако самыми широко распространенными усатыми китами неогена были цетотерии (Cetotheriidae) — очень разнообразная (более 50 видов и около 20 родов) группа сравнительно небольших форм (около 3–4 м) без признаков специализации современных семейств. Судя по всему, в основном на них и охотились кашалоты-убийцы, а также гигантские акулы мегалодоны. Остатки цетотериев известны из Европы, Северной и Южной Америки, Японии, Австралии и Новой Зеландии.

***
Итак, появившись почти 55 млн лет назад в прибрежных водах Индо-Пакистанской области, китообразные постепенно освоили Мировой океан, породив огромное множество форм весьма разнообразного облика и величины.
Благодаря палеонтологическим открытиям последнего времени китообразные стали одной из немногих групп млекопитающих с хорошо реконструированной эволюционной историей. Очень важно, что палеонтологические данные хорошо согласуются с молекулярно-генетическими*, дополняя друг друга и независимо фиксируя последовательность и время возникновения соответствующих ключевых анатомических, физиологических и биохимических адаптаций и магистральных эволюционных событий при переходе от наземных форм к полуводным, от полуводных к прибрежно-морским и от прибрежно-морских — к океаническим. Несомненно, новые исследования ископаемых и современных видов в ближайшее время существенно дополнят и детализируют складывающуюся картину.

Ископаемая летопись китообразных постоянно пополняется яркими находками, сулящими дальнейшие открытия. В России остатки вымерших китов встречаются довольно редко, в основном в отложениях миоцена (Ставрополье, Дагестан, Таманский п-ов, Сахалин). В Ростовской обл. обнаружены фрагментарные остатки среднеэоценовых археоцетов [24]. На Северном Кавказе находят цетотериев. Недавно оттуда были описаны несколько новых родов этих китов. Важную находку передал в Палеонтологический институт имени А. А. Борисяка РАН житель Чеченской республики Х. А. Абдулмежидов, раскопавший в Ножай-Юртовском р-не почти полный скелет кита. Этот новый кит из группы цетотериев получил название «вампал» (Vampalus sayasanicus), по имени великана из чеченской мифологии [25].
Литература
1. Grauer D., Higgins D. Molecular evidence for the inclusion of cetaceans within the order Artiodactyla // Molecular Biology and Evolution. 1994; 11(3): 357–364.
2. Thewissen J. G. M., Williams E. M., Roe L. J., Hussain S. T. Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls // Nature. 2001; 413: 277–281.
3. Thewissen J. G. M., Copper L. N., Clementz M. T. et al. Whales originated from aquatic artiodactyls in the Eocene epoch of India // Nature. 2007; 450(7173): 1190–1194.
4. Spoor F., Bajpai S., Hussain S. T. et al. Vestibular evidence for the evolution of aquatic behaviour in early cetaceans // Nature. 2002; 417(6885): 163–166.
5. Williams E. M. Synopsis of the earliest cetaceans: Pakicetidae, Ambulocetidae, Remingtonocetidae, and Protocetidae // The emergence of whales. Evolutionary patterns in the origin of Cetacea. N.Y., 1998: 1–28.
6. Thewissen J. G. M., Bajpai S. New skeletal material of Andrewsiphius and Kutchicetus, two Eocene cetaceans from India // Journal of Paleontology. 2009; 83(3): 635–663.
7. Bebej R. M., Zalmout I. S., Abed El-Aziz A. A. et al. First remingtonocetid archaeocete (Mammalia, Cetacea) from the middle Eocene of Egypt with implications for biogeography and locomotion in early cetacean evolution // Journal of Paleontology. 2016; 89(5): 882–893.
8. Bajpai S., Thewissen J. G. M., Sahni A. The origin and early evolution of whales: macroevolution documented on the Indian Subcontinent // Journal of Biosciences. 2009; 34(5): 673–686.
9. Gingerich P. D., Zalmout I. S., Ul-Haq M., Bhatti M. A. Makaracetus bidens, a new protocetid archaeocete (Mammalia, Cetacea) from the Early Middle Eocene of Balochistan (Pakistan) // Contributions from the Museum of Paleontology, University of Michigan. 2005; 31(9): 197–210.
10. Gingerich P. D., Ul-Haq M., Koenigswald W. von et al. New protocetid whale from the Middle Eocene of Pakistan: birth on land, precocial development, and sexual dimorphism // PLoS One. 2009; 4(2): e4366: 1–20.
11. Thewissen J. G. M., Williams E. M. The early radiation of Cetacea (Mammalia): evolutionary pattern and developmental correlation // Annual Review of Ecology and Systematics. 2002; 33: 73–90.
12. Marino L., McShea D. W., Uhen M. D. Origin and evolution of large brains in toothed whales // Anatomical Record. Pt A. 2004; 281A: 1247–1255.
13. Geisler J. H., Colbert M. W., Carew J. L. A new fossil species supports an early origin for toothed whale echolocation // Nature. 2014; 508(7496): 383–386.
14. Fordyce R. E. Simocetus rayi (Odontoceti: Simocetidae) (new species, new genus, new family), a bizarre archaic Oligocene dolphin from the eastern North Pacific // Smithsonian Contributions to Paleobiology. 2002; 93: 185–222.
15. Racicot R. A., Deméré T. A., Beatty B. L., Boessenecker R. W. Unique feeding morphology in a new prognathous extinct porpoise from the Pliocene of California // Current Biology. 2014; 24(7): 774–779.
16. Hamilton H., Caballero S., Collins A. G., Brownell R. L. Evolution of river dolphins // Proceedings of the Royal Society of London. Ser. B. 2001; 268: 549–556.
17. Muizon C. de. Walrus-like feeding adaptation in a new cetacean from the Pliocene of Peru // Nature. 1993; 365(6448): 745–748.
18. Bianucci G., Landini W. Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy // Zoological Journal of the Linnean Society. 2006; 148: 103–131.
19. Lambert O., Bianucci G., Muizon C. de. Macroraptorial sperm whales (Cetacea, Odontoceti, Physeteroidea) from the Miocene of Peru // Zoological Journal of the Linnean Society. 2017; 179: 404–474.
20. Lambert O., Bianucci G., Post K. et al. The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru // Nature. 2010; 466(7302): 105–108.
21. Fitzgerald E. M. G. A bizarre new toothed mysticete (Cetacea) from Australia and the early evolution of baleen whales // Proceedings of the Royal Society of London. Ser. B. 2006. 273(1604): 2955–2963.
22. Fordyce R. E., Marx F. G. Mysticetes baring their teeth: a new fossil whale, Mammalodon hakataramea, from the Southwest Pacific // Memoirs of Museum Victoria. 2016; 74: 107–116.
23. Deméré T. A., Berta A., McGowen M. R. The taxonomic and evolutionary history of fossil and modern balaenopteroid mysticetes // Journal of Mammalian Evolution. 2005; 12(1/2): 99–143.
24. Тесаков А. С., Александрова Г. Н., Тарасенко К. К. и др. Палинологическая характеристика палеогеновых отложений и находка древнего кита на юго-западном побережье Цимлянского водохранилища (Ростовская область) // Бюллетень Региональной межведомственной стратиграфической комиссии по центру и югу Русской платформы. Вып. 5. М., 2012: 138–143.
25. Тарасенко К. К., Лопатин А. В. Новые роды усатых китов (Cetacea, Mammalia) из миоцена Северного Кавказа и Предкавказья. 2: Vampalus gen. nov. (средний — поздний миоцен, Чечня и Краснодарский край) // Палеонтол. журн. 2012; 6: 72–81.
* См. подробный обзор: Гельфанд М. С. Молекулярная эволюция: как киты уходили под воду // Природа. 2016. № 10. С. 39–50.