Фотосинтез — это создание органических веществ из неорганических с помощью световой энергии.
Другими словами, световая энергия превращается в химическую. Для этого нам нужны:
1. Пигменты (хлорофилл)
2. Мембраны
✍🏼 Друг без друга они никак, потому что хлорофилл не плавает и не висит в воздухе, он закреплен в этой мембране — тилакоиде. Тилакоиды обязательно есть у всех фотосинтезирующих организмов. Даже у цианобактерий. Однажды, такую цианобактерию фагоцитировала какая-то амебоподобная клетка и оставила себе на память. Так получились хлоропласты. То есть, у цианобактерий тилакоиды находятся в цитоплазме, и у растений как будто бы тоже в цитоплазме цианобактерий, только они теперь называются хлоропласты. Осознали?
📌 Фотосинтез делят на две стадии: световую и темновую.
 1. Световая фаза
1. Световая фаза
Идет на тилакоидах с хлорофиллом. Он улавливает свет с помощью своего электрона, который «вышибается» из молекулы хлорофилла. С помощью энергии этого электрона синтезируется АТФ. Но если электрон вылетел — в хлорофилле «дырка». Её нужно закрыть. Где взять дополнительный электрон? В молекуле воды, конечно.
Но если забрать электрон у молекулы воды, то она становится нестабильной и распадется на Н+ (протон водорода) и кислород. Кислород не нужен. Он улетает. Н+ связывается с НАДФ (Н+НАДФ = НАДФ-Н2). Он потом будет нужен.
Итак, в световой фазе образовалось два нужных продукта: АТФ и НАДФ-Н2 и один ненужный, который улетел.
2. Темновая фаза
Идёт уже не в самих тилакоидах, а в жидкости вокруг них (строма хлоропласта). Здесь происходит фиксация углекислого газа. Запомните это словосочетание❗️ ФИКСАЦИЯ УГЛЕКИСЛОГО ГАЗА.
Это означает что из него синтезируется глюкоза. Реакция: СО2 + Н2О = Глюкоза. «Цэ-о-два» и «Аж(целых)-два-о» — это простые вещества, поэтому соединить их вместе очень сложно. Нужно много энергии. Здесь мы и используем АТФ из световой фазы. А так же НАДФ-Н2.
📍 Важное замечание: темновая фаза так названа, не потому что ночью происходит, а потому что ей не нужен свет. Ей нужны только СО2, вода и АТФ. Если все это имеется, она может идти и на свету и в темноте.

Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter. Мы обязательно поправим!
Типы питания
По типу питания живые организмы делятся на автотрофы, гетеротрофы и миксотрофы. Автотрофы (греч. αὐτός — сам + τροφ — пища)
— организмы, которые самостоятельно способны синтезировать органические вещества из неорганических. Гетеротрофы (греч. ἕτερος
— иной + τροφή — пища) — организмы, использующие для питания готовые органические вещества.
Наконец, миксотрофы (греч. μῖξις — смешение + τροφή — пища) — организмы, которые могут использовать как гетеротрофный, так и
автотрофный способ питания. К примеру, эвглена зеленая на свету начинает фотосинтезировать, а в темноте питается гетеротрофно.

Фотосинтез
Фотосинтез (греч. φῶς — свет и σύνθεσις — синтез) — сложный химический процесс преобразования энергии квантов света в
энергию химических связей. В результате фотосинтеза происходит синтез органических веществ из неорганических.

Этот процесс уникален и происходит только в растительных клетках, а также у некоторых бактерий. Фотосинтез осуществляется при участии хлорофилла (греч. χλωρός — зелёный и φύλλον — лист) — зеленого пигмента, окрашивающего органы растений в
зеленый цвет. Существуют и другие вспомогательные пигменты, которые вместе с хлорофиллом выполняют светособирающую
или светозащитную функции.
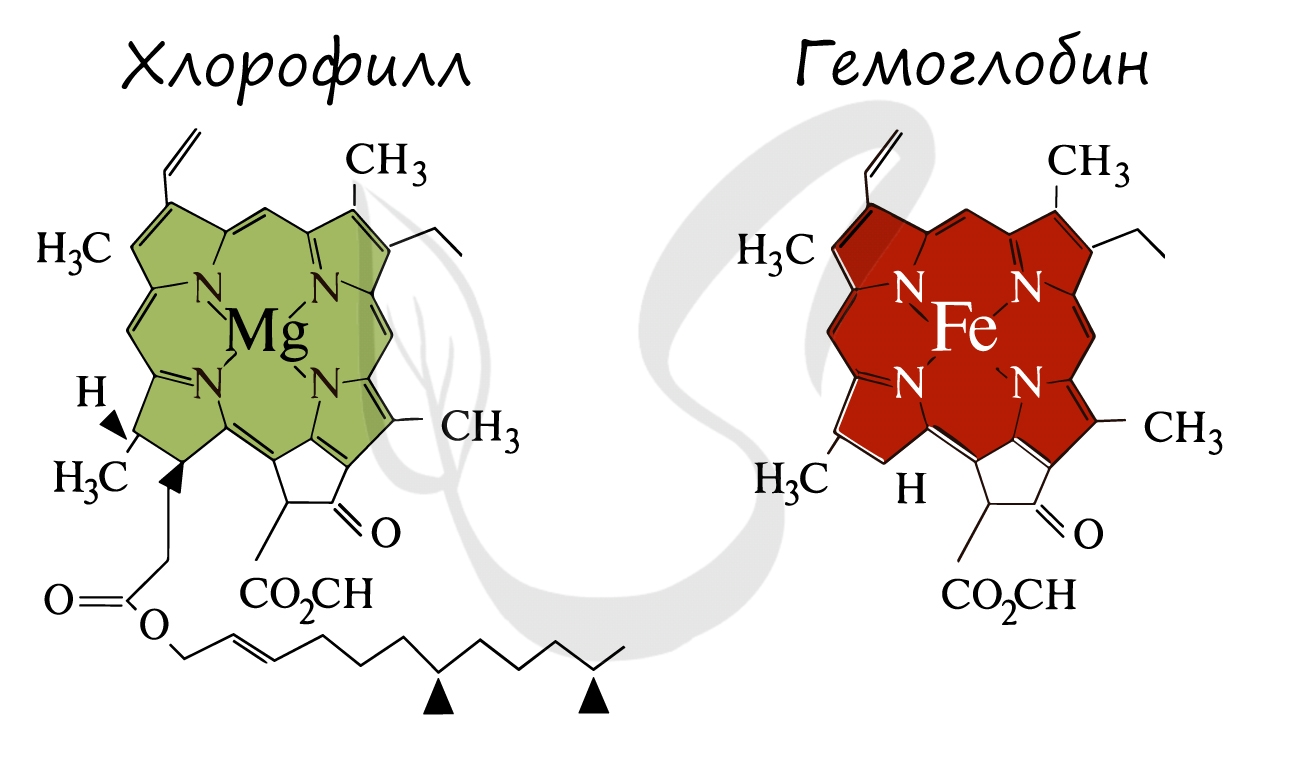
Ниже вы увидите сравнение строения хлорофилла и гемоглобина. Обратите внимание, что в центре молекулы хлорофилла находится
ион Mg.
В высшей степени гениально значение процесса фотосинтеза подчеркнул русский ученый К.А. Тимирязев: «Все органические вещества,
как бы они ни были разнообразны, где бы они ни встречались, в растении ли, в животном или человеке, прошли через лист, произошли
от веществ, выработанных листом. Вне листа или, вернее, вне хлорофиллового зерна в природе не существует лаборатории, где бы выделялось
органическое вещество. Во всех других органах и организмах оно превращается, преобразуется, только здесь оно образуется вновь
из вещества неорганического»
Более подробно мы обсудим значение фотосинтеза в завершение этой статьи. Фотосинтез состоит из двух фаз: светозависимой (световой)
и светонезависимой (темновой). Я рекомендую использовать названия светозависимая и светонезависимая, так как они способствуют
более глубокому (и правильному!) пониманию фотосинтеза.
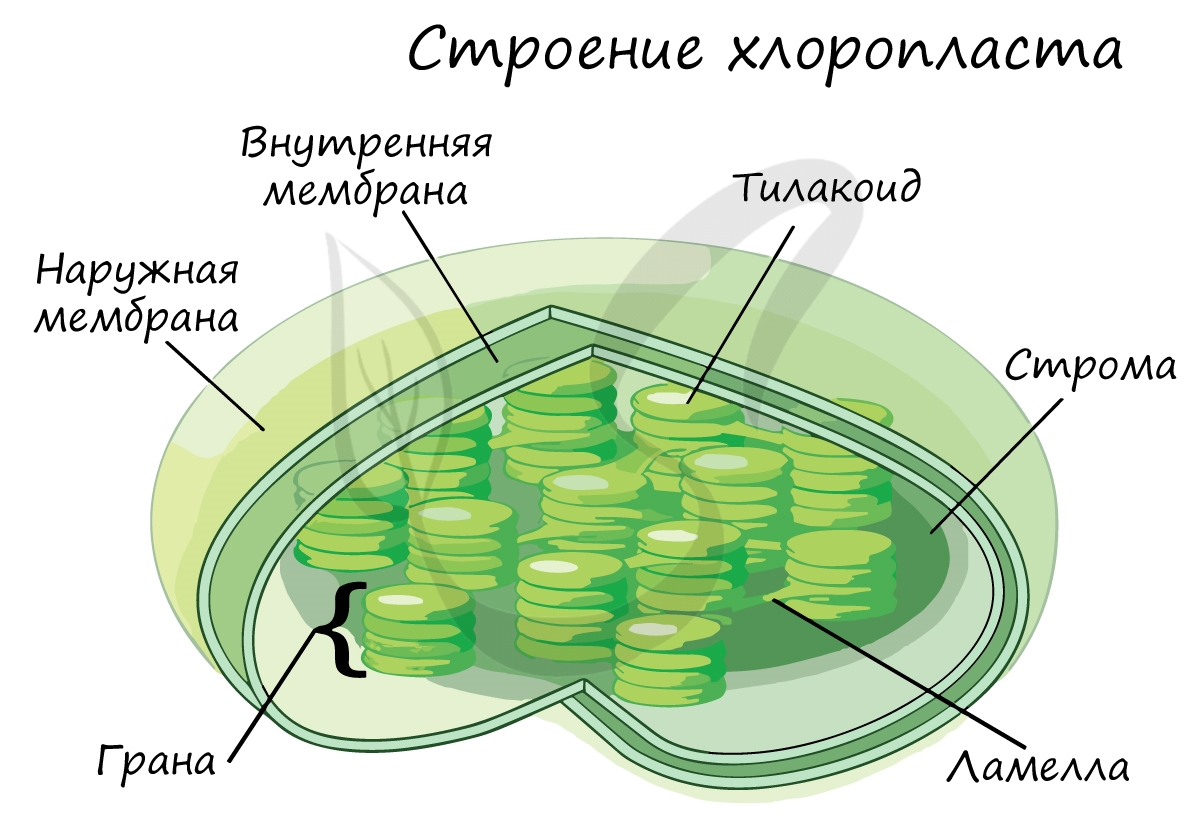
Светозависимая фаза (световая)
Эта фаза происходит только на свету на мембранах тилакоидов в хлоропластах. В ней принимают участие различные ферменты,
белки-переносчики, молекулы АТФ-синтетазы и зеленый пигмент хлорофилл.
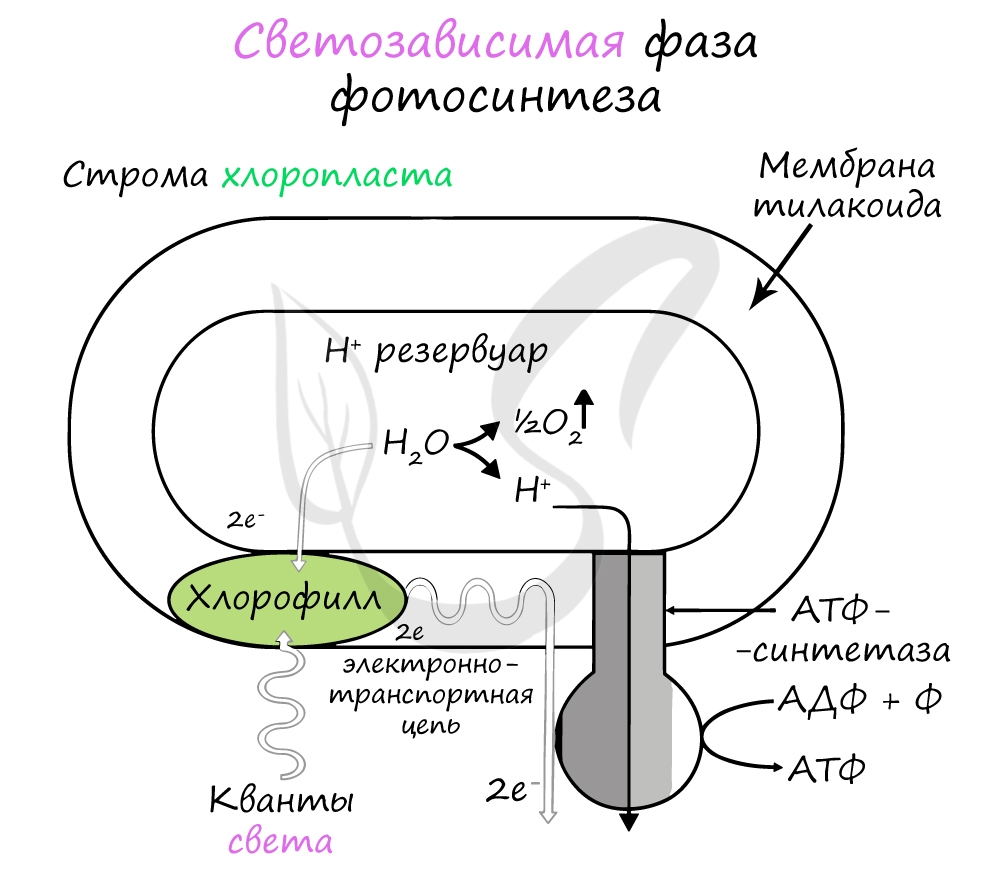
Хлорофилл выполняет две функции: поглощения и передачи энергии. При воздействии кванта света хлорофилл теряет электрон,
переходя в возбужденное состояние. С помощью переносчиков электроны скапливаются с наружной поверхности мембраны тилакоидов,
тем временем внутри тилакоида происходит фотолиз воды (разложение под действием света):
H2O —> H+ + OH—
Гидроксид-ионы отдают лишний электрон, превращаясь в реакционно способные радикалы OH, которые собираются вместе и образуют молекулу воды и свободный кислород (это побочный продукт, который в дальнейшем удаляется в ходе газообмена).
4OH —> 2H2O + O2↑
Образовавшиеся при фотолизе воды протоны (H+) скапливаются с внутренней стороны мембраны тилакоидов, а
электроны — с внешней. В результате по обе стороны мембраны накапливаются противоположные заряды.
При достижении критической разницы, часть протонов проталкивается на внешнюю сторону мембраны через канал АТФ-синтетазы.
В результате этого выделяется энергия, которая может быть использована для фосфорилирования молекул АДФ:
Протоны, попав на поверхность мембраны тилакоидов, соединяются с электронами и образуют атомарный водород, который
используется для восстановления молекулы-переносчика НАДФ (никотинамиддинуклеотидфосфат). Благодаря этому окисленная
форма — НАДФ+ превращается в восстановленную — НАДФ∗H2.
Предлагаю создать квинтэссенцию из полученных нами знаний. Итак, в результате светозависимой фазы фотосинтеза образуются:
- Свободный кислород O2 — в результате фотолиза воды
- АТФ — универсальный источник энергии
- НАДФ∗H2 — форма запасания атомов водорода
Кислород удаляется из клетки как побочный продукт фотосинтеза, он совершенно не нужен растению. АТФ и НАДФ∗H2
в дальнейшем оказываются более полезны: они транспортируются в строму хлоропласта и принимают участие в светонезависимой
фазе фотосинтеза.
Светонезависимая (темновая) фаза
Светонезависимая фаза происходит в строме (матриксе) хлоропласта постоянно: и днем, и ночью — вне зависимости от
освещения.
При участии АТФ и НАДФ∗H2 происходит восстановление CO2 до глюкозы C6H12O6.
В светонезависимой фазе происходит цикл Кальвина, в ходе которого и образуется глюкоза. Для образования одной молекулы глюкозы
требуется 6 молекул CO2, 12 НАДФ∗H2 и 18 АТФ.
Таким образом, в результате темновой (светонезависимой) фазы фотосинтеза образуется глюкоза, которая в дальнейшем может быть преобразована
в крахмал, служащий для запасания питательных веществ у растений.
Значение фотосинтеза
Значение фотосинтеза невозможно переоценить. Уверенно утверждаю: именно благодаря этому процессу жизнь на Земле приобрела такие
чудесные и изумительные формы, какие мы видим вокруг себя: удивительные растения, прекрасные цветы и самые разнообразные животные.
В разделе эволюции мы уже обсуждали, что изначально в составе атмосферы Земли не было кислорода: миллиарды лет назад его начали вырабатывать
первые фотосинтезирующие бактерии — сине-зеленые водоросли (цианобактерии). Постепенно кислород накапливался, и со временем на Земле
стало возможно аэробное (кислородное) дыхание. Возник озоновый слой, защищающий все живое на нашей планете от губительного ультрафиолета.
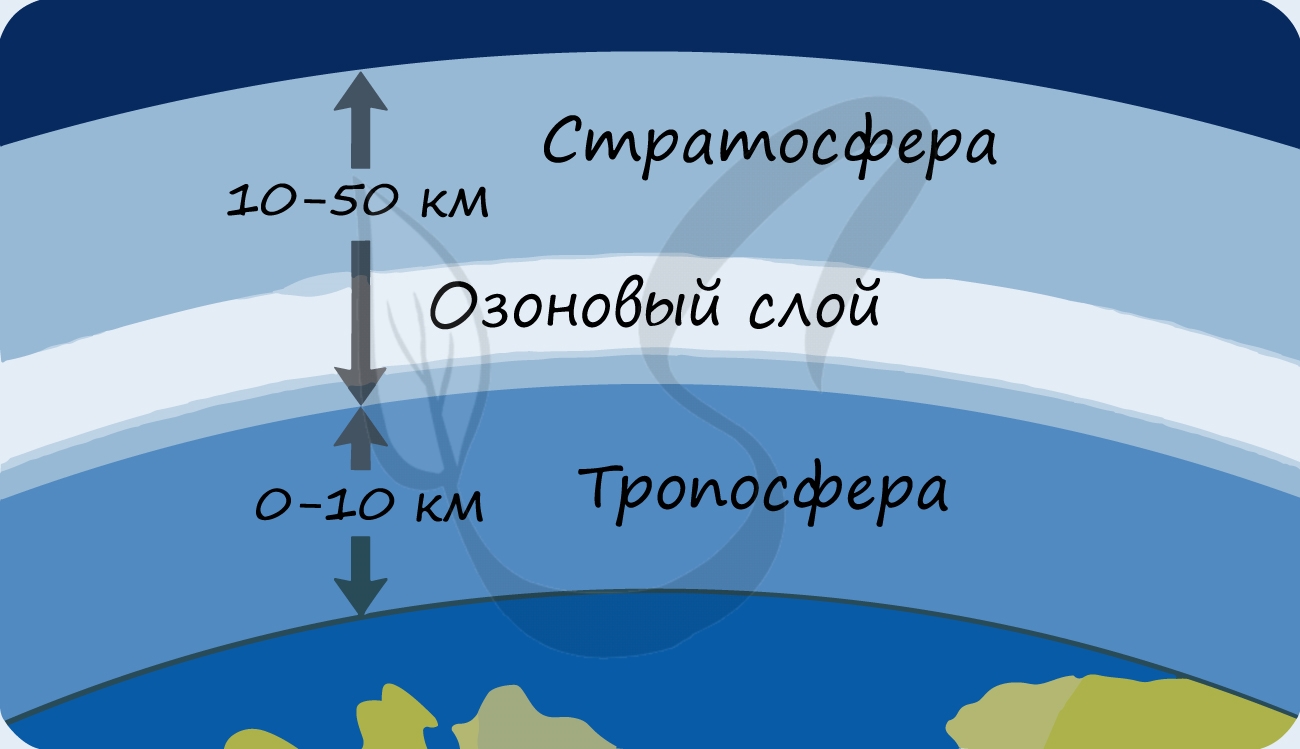
Говоря о роли фотосинтеза, выделим следующие функции, объединяющиеся в так называемую космическую роль растений. Итак, растения за счет фотосинтеза:
- Синтезируют органические вещества, являющиеся пищей для всего живого на планете
- Преобразуют энергию света в энергию химических связей, создают органическую массу
- Растения поддерживают определенный процент содержания O2 в атмосфере, очищают ее от избытка CO2
- Способствуют образованию защитного озонового экрана, поглощающего губительное для жизни ультрафиолетовое излучение

Хемосинтез (греч. chemeia – химия + synthesis — синтез)
Хемосинтез — автотрофный тип питания, который характерен для некоторых микроорганизмов, способных создавать органические
вещества из неорганических. Это осуществляется за счет энергии, получаемой при окислении других неорганических соединений
(железо- , азото-, серосодержащих веществ).
Хемосинтез был открыт русским микробиологом С.Н. Виноградским в 1888 году. Большинство хемосинтезирующих бактерий относится
к аэробам, для жизни им необходим кислород.
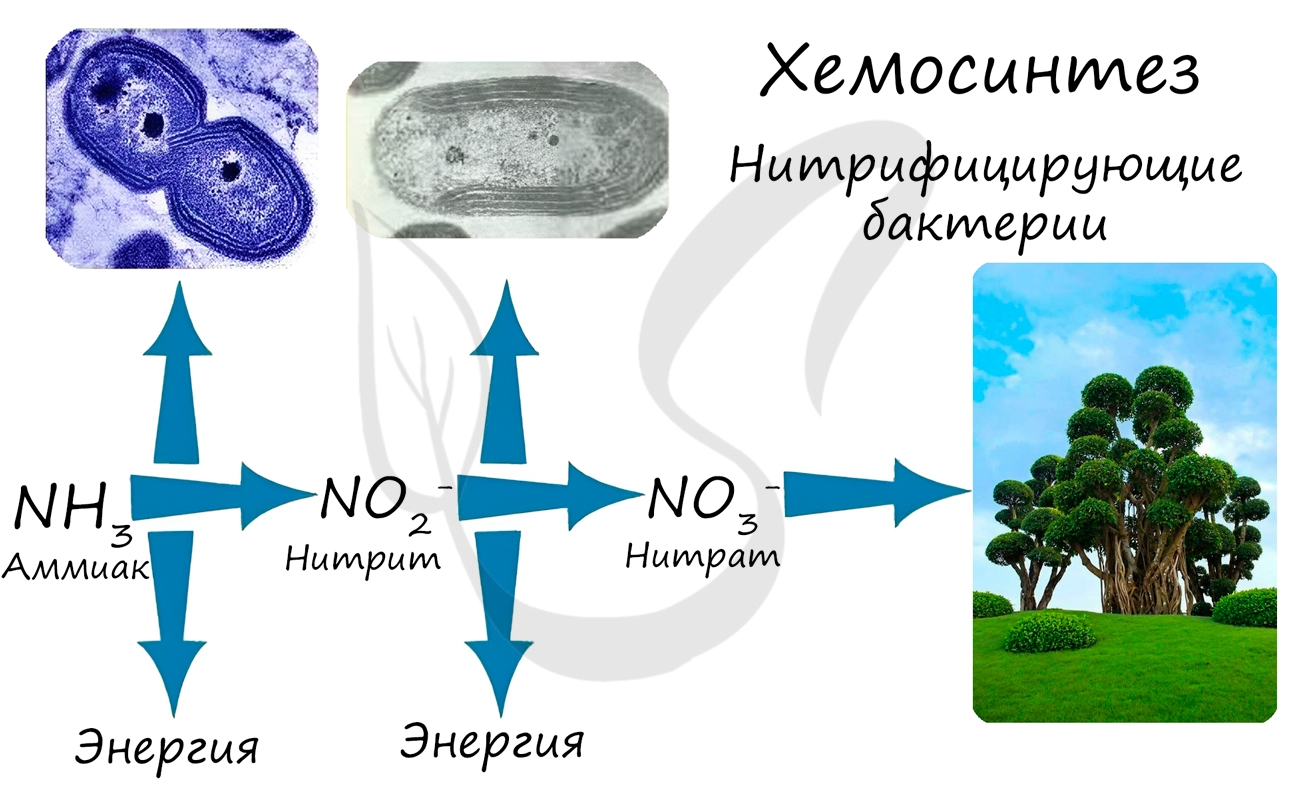
При окислении неорганических веществ выделяется энергия, которую организмы запасают в виде энергии химических связей.
Так нитрифицирующие бактерии последовательно окисляют аммиак до нитрита, а затем — нитрата. Нитраты могут быть усвоены
растениями и служат удобрением.
Помимо нитрифицирующих бактерий, встречаются:
- Серобактерии — окисляют H2S —> S 0 —> (S+4O3)2- —> (S+6O4)2-
- Железобактерии — окисляют Fe+2 —>Fe+3
- Водородные бактерии — окисляют H2 —> H+12O
- Карбоксидобактерии — окисляют CO до CO2
Значение хемосинтеза
Хемосинтезирующие бактерии являются неотъемлемым звеном круговорота в природе таких элементов как: азот, сера, железо.
Нитрифицирующие бактерии обеспечивают переработку (нейтрализацию) ядовитого вещества — аммиака. Они также обогащают
почву нитратами, которые очень важны для нормального роста и развития растений.
Усвоение нитратов происходит за счет клубеньковых бактерий на корнях бобовых
растений, однако важно помнить, что клубеньковые (азотфиксирующие) бактерии, в отличие от нитрифицирующих бактерий, питаются гетеротрофно.
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Фотосинтез
Автор статьи — Л.В. Окольнова.

Определение довольно простое, уравнение тоже суммарное. оно не описывает сам процесс — сложный и многоступенчатый.
В этой статье мы не будем разбирать все стадии, мы разберем только две основные фазы фотосинтеза — световую и темновую, а также основные процессы, которые происходят в это время в организме растения.
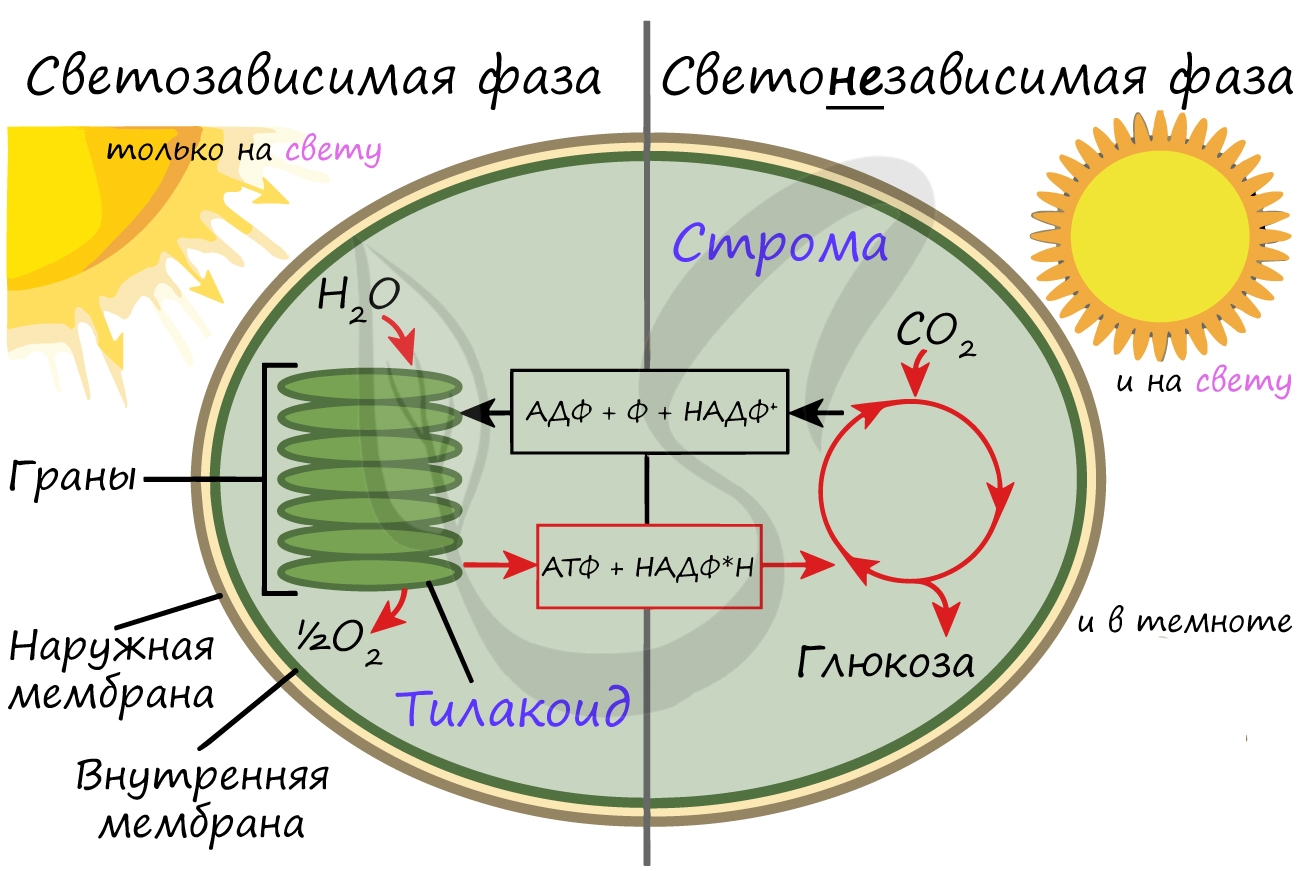
Световая фаза фотосинтеза.
Днем растения работают как солнечные батарейки — аккумулируют энергию света солнца:
● на мембранах тилакойдов хлоропластов молекулы хлорофилла поглощают (аккумулируют) свет,

● происходит синтез АТФ,

● образуется НАДФ — кофермент.
Кофермент (коэнзим) — это биологический катализатор, но ферментом его назвать нельзя, т.к. у него не белковая природа, который ускоряет и направляет протекание окислительно-восстановительных процессов. Он понадобится на следующей — темновой фазе процесса .
●происходит расщепление (фотолиз) воды: 2H20 = 4H+ + 4e- + O2.
растение выделяет кислород .
Темновая фаза фотосинтеза.
Это уже фаза синтеза. Энергия, полученная в ходе световой фазы, идет на восстановление CO2 до молекулы глюкозы.
Этот процесс происходит уже в строме.
Общая схема фотосинтеза:

Спасибо за то, что пользуйтесь нашими статьями.
Информация на странице «Фотосинтез» подготовлена нашими редакторами специально, чтобы помочь вам в освоении предмета и подготовке к экзаменам.
Чтобы успешно сдать нужные и поступить в ВУЗ или колледж нужно использовать все инструменты: учеба, контрольные, олимпиады, онлайн-лекции, видеоуроки, сборники заданий.
Также вы можете воспользоваться другими материалами из разделов нашего сайта.
Публикация обновлена:
08.03.2023
Чем растения отличаются от других царств живой природы? Несмотря на то, что отличий масса, скорее всего, в первую очередь вы подумаете о фотосинтезе. Так что именно о фотосинтезе на ЕГЭ и ОГЭ мы сейчас и поговорим.

Что такое фотосинтез?
Почему растения фотосинтезируют? Стандартный ответ: «Потому что они зеленые».
На самом деле, растения получили способность к фотосинтезу благодаря наличию симбиотических органоидов — хлоропластов, в которых и происходят темновая и световая фазы, а в хлоропластах содержится пигмент хлорофилл, именно он окрашивает растения в зеленый цвет.
Фотосинтез — одна из реакций обмена веществ. Как любая реакция метаболизма, он идет поэтапно (световая и темновая фазы) и с участием ферментов. Фотосинтез относится к реакциям пластического обмена. Особенность пластического обмена в том, что органические вещества синтезируются, а энергия на это тратится.
Фотосинтез — это синтез органических веществ из неорганических веществ с использованием энергии солнечного света.
Далее разберем подробно обе фазы и процессы, происходящие в них.
Как идет процесс фотосинтеза?
Световая фаза фотосинтеза для ЕГЭ и ОГЭ
Световая фаза проходит в хлоропластах на тилакоидах. Там хранится пигмент хлорофилл, с которого все начинается — именно из-за него растения имеют зеленую окраску. Квант света попадает на тилакоид и возбуждает молекулу хлорофилла. В этот момент инициируется процесс фотосинтеза. При этом выделяется энергия АТФ.
Самые внимательные из вас могут заметить некоторую несостыковку. Почему выделяется? Это же реакция пластического обмена, а не энергетического, значит, энергия должна тратиться. Да, действительно при фотосинтезе выделяется АТФ, но она не накапливается и не тратится на другие реакции, как при энергетическом обмене, а вся уходит на фотосинтез. Поэтому это реакция анаболизма, хоть и с выделением АТФ.
Параллельно идет фотолиз воды.
Название процесса говорит само за себя: «фото» — свет, «лизис» — расщепление. Буквально переводится как расщепление воды на свету. Легко запомнить, что проходит фотолиз в световую фазу.
На что же может распасться молекула воды? На свободный кислород и водород. У каждого из этих элементов свой путь.
Кислород — это сильный окислитель, буквально смерть для любой неспециализированной клетки, поэтому растения быстро от него избавляются, выделяя в атмосферу как побочный продукт. А уже из атмосферы аэробные организмы (в том числе, растения) поглощают его и используют для дыхания. Так что нам повезло! Не было бы процесса фотосинтеза, не было бы кислорода и что было бы с жизнью на нашей планете представить сложно.
Но помимо кислорода, выделяется еще водород, если бы он был человеком, мы бы сказали, что он растерян и нуждается в помощи. На помощь к нему приходит молекула-переносчик НАДФ (полное ее название —никотинамиддинуклеотидфосфат, но мы ласково зовем ее НАДФ). Она использует водород для восстановления до НАДФ*Н2. Задача этой молекулы переносить водород из тилакоидов в строму, поэтому мы называем ее молекула-переносчик. На этом световая фаза заканчивается.
Резюмируем
- Квант света возбуждает молекулу хлорофилла
- Инициируется процесс фотосинтеза
- Выделяется АТФ
- Фотолиз воды
- Кислород выходит в окружающую среду как побочный продукт фотосинтеза
- Водород соединяется с молекулой переносчиком НАДФ*
Темновая фаза фотосинтеза для ЕГЭ и ОГЭ
В некоторых источниках эту фазу еще называют светонезависимой фазой. Действительно, название «темновая стадия» часто вызывает затруднения. Кажется, что световая проходит на свету, а темновая тогда в темноте, но это не так. Для темновой фазы действительно не нужен свет, соответственно, у нее есть варианты — может проходить и на свету, и в темноте. Она идет практически параллельно со световой и в ней используются продукты, образовавшиеся в световой фазе.
Для того чтобы фазы друг другу не мешали, они проходят в разных частях хлоропласта. Световая, как мы уже выяснили, идет на тилакоидах, а темновая в строме — это внутренняя полужидкая среда хлоропласта.
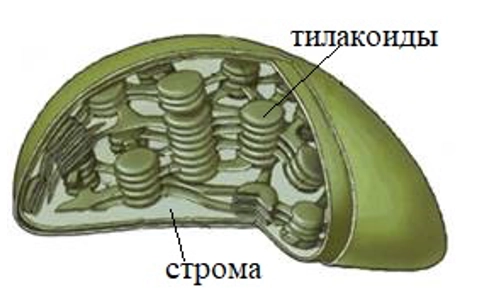
В строму приходят АТФ, молекула-переносчик приносит водород. Но из водорода и энергии ничего органического создать не получится, нужны еще элементы. Растения нашли гениальный выход, они используют вещество, которого достаточно в атмосфере, следовательно, за него нет конкуренции. Это вещество — углекислый газ.
Дальше начинается очень сложный циклический процесс, который называется цикл Кальвина. Мы не будем слишком подробно его рассматривать, это не пригодится для государственных экзаменов, но именно в нем активно работают ферменты, и на него тратится энергия АТФ, полученная в световой фазе. В результате цикла Кальвина образуется шестиуглеродный сахар-глюкоза. Далее эта глюкоза может быть переработана в крахмал и откладываться растением как запасной углевод.
Резюмируем
- Фиксация СО2
- Цикл Кальвина
- Синтез глюкозы
- Образование крахмала
Значение фотосинтеза
На Земле, пожалуй, практически не существует процессов, которые повлияли на эволюцию планеты так же сильно, как фотосинтез. Давайте разберем основные значения фотосинтеза:
- Сформировалась атмосфера с высоким содержанием кислорода, пригодная для дыхания. Аэробные организмы, включая человека, проводят энергетический обмен с использованием кислорода и получают энергию для жизнедеятельности.
- Возникновение озонового слоя. Вследствие фотосинтеза в атмосфере накопился кислород, что привело к появлению озонового экрана. Жизнь, которая до этого вынуждена была развиваться под водой, боясь ультрафиолета, смогла выйти на сушу и освоить ее.
- Синтез органических веществ. Растения — автотрофные организмы, сами производят органические вещества, которые затем используют гетеротрофы. Вещества, которые образуют растения в процессе фотосинтеза, являются первичным источником веществ и энергии практически для всех живых организмов.
Примеры заданий на фотосинтез в ЕГЭ и ОГЭ по биологии
Вопросы по фотосинтезу встречаются как в ЕГЭ, так и в ОГЭ. Причем, если для 9 класса достаточно знать что это такое и основные этапы, то для ЕГЭ необходимо понимание последовательности процессов. Кстати, актуальна эта тема для решения новых заданий по экспериментам (2 и 22 линии в ЕГЭ 2022).
Задание на фотосинтез в ОГЭ по биологии

Решение. Типичный вопрос для первой части ОГЭ из открытого банка ФИПИ. Какие из этих процессов происходят во время фотосинтеза? Возбуждение молекул хлорофилла квантом света, расщепление (фотолиз) воды и образование глюкозы.
Во время фотосинтеза, наоборот, выделяется кислород, как побочный продукт, и поглощается углекислый газ. А синтез белка вообще проходит на рибосомах.
Ответ. 123
Задание на фотосинтез в ЕГЭ по биологии

Решение. Это задание из открытого варианта 2021 года (в 2021 эти варианты заменяли варианты досрочного ЕГЭ). Необходимо соотнести процессы и фазы. В световой фазе происходит возбуждение молекулы хлорофилла, фотолиз воды и образование энергии. В темновую фазу фиксируется углекислый газ и восстановление углерода водородом для синтеза глюкозы.
Ответ. 12212
Конечно, процесс фотосинтеза значительно сложнее, чем мы с вами разобрали. Да и на ОГЭ и ЕГЭ проверяют знание многих других тем. Чтобы сдать экзамен на высокий балл, надо знать анатомию, зоологию, генетику, микробиологию и даже психологию. При этом недостаточно только хорошо разбираться в основных темах. Надо уметь избегать ловушек экзаменаторов, вчитываться в формулировки заданий и оформлять ответы в четком соответствии с критериями. Поэтому необходимо готовиться к ОГЭ и ЕГЭ по биологии системно.
Экзамен по биологии — не шутка. Если вы хотите сдать его на 90+, записывайтесь на мои курсы подготовки к ОГЭ или ЕГЭ. Мы разберемся со всеми темами, которые спрашивают в 9 или 11 классе, научимся решать задания быстро и правильно, а также разберем основные лайфхаки, которые помогут вам не стрессовать. Я также проведу с вами пробный экзамен в формате реального ОГЭ или ЕГЭ, чтобы вы были готовы к любым неожиданностям. После мы разберем все ошибки и поймем, как избежать их в будущем. Приходите на мои занятия, и я помогу вам сдать ОГЭ или ЕГЭ на самый высокий балл!
Шпаргалка: Фотосинтез
Содержание
1. МЕХАНИЧЕСКИЕ ТКАНИ… 2
2. АНАТОМИЧЕСКОЕ СТРОЕНИЕ ОСЕВЫХ ОРГАНОВ… 6
2.1. Анатомическое строение стебля двудольного растенияпучкового строения 6
3. ФОТОСИНТЕЗ. 11
4. КОМПЛЕКСЫ ТКАНЕЙ… 17
4.1. Понятие о флоэме. 17
4.2. Понятие о ксилеме. 19
5. ХАРАКТЕРИСТИКА СЕМЕЙСТВ: РОЗОЦВЕТНЫЕ И ЯСНОТКОВЫЕ 22
СПИСОК ИСПОЛЬЗУЕМОЙ ЛИТЕРАТУРЫ. 23
1. МЕХАНИЧЕСКИЕ ТКАНИ
Механическиеткани – это опорные ткани, придающие прочность органам растений. Ониобеспечивают сопротивление статическим и динамическим нагрузкам.
В самыхмолодых участках растущих органов механических тканей нет, так как живые клеткив состоянии высокого тургора обусловливают их форму благодаря своим упругимстенкам. По мере увеличения размеров организма и развития органов в нихпоявляются специализированные механические ткани. Сочетаясь с другими тканями,они образуют как бы арматуру органа, поэтому их называют арматурными. Иногдавсю систему механических тканей называют стереомом, а составляющие ее клетки –стереидами.
Степеньразвития механических тканей во многом зависит от условий обитания. Онаневелика у растений гидрофитов и значительна у растений засушливыхместообитаний – склерофитов.
Механическиеткани наиболее развиты в осевой части побега – стебле. Здесь они чащерасполагаются по его периферии: либо отдельными участками в гранях стебля, либосплошными кольцами.
Напротив, вкорне, который выдерживает главным образом сопротивление на разрыв,механическая ткань сосредоточена обычно в центре. В листьях механические тканирасполагаются в соответствии с принципом устройства двутавровой балки.Механические ткани могут формироваться как из первичных, так и из вторичныхмеристем.
Наиболеезаметная особенность клеток механических тканей – их значительно утолщенныеоболочки, которые продолжают выполнять опорную функцию даже после отмирания ихживого содержимого. Различают три основных типа механических тканей:
Колленхима –это простая первичная опорная ткань, состоящая из более или менее вытянутыхвдоль оси органа клеток с утолщенными слоистыми неодревесневшими первичнымиоболочками. В зависимости от характера утолщений стенок и соединения клетокмежду собой различают:
1.1. Уголковуюколленхиму – на поперечном срезе утолщенные части оболочек соседних клетокзрительно сливаются между собой, образуя трех-, четырех — или пятиугольники.
1.2 Пластинчатуюколленхиму – клеточная оболочка утолщена равномерно.
1.3. Рыхлуюколленхиму – имеются видимые межклетники.
Колленхимаформируется из основной меристемы и обычно располагается непосредственно подэпидермой либо на расстоянии одного или нескольких слоев клеток от нее. Вмолодых стеблях она часто образует сплошной цилиндр по периферии. Иногдаколленхима встречается в форме продольных тяжей в выступающих ребрах стеблейтравянистых и тех частей древесных растений, которые еще не вступили в стадиювторого роста. Обычно колленхима в черешках и по обеим сторонам крупных жилок.Корни содержат колленхиму редко. Клетки колленхимы, будучи живыми снеодревесневшими стенками, способны к росту в длину и не препятствуют ростуорганов, в которых они расположены. Иногда колленхима содержит хлоропласты.
Функцииарматурной ткани колленхима может выполнять только в состоянии тургора.Эволюционно колленхима возникла из паренхимы основной ткани и близка к ней.
2.Склеренхима – механическая ткань, состоящая из прозенхимных клеток содревесневшими, или реже неодревесневающими и равномерно утолщеннымиоболочками. Оболочки склеренхимных клеток обладают прочностью, близкой кпрочности стали. Оболочки их толсты, а полость клетки мала и узка. Отложениелигнина повышает прочность склеренхимы. Поры в оболочках склеренхимынемногочисленные, простые.
По сравнениюс колленхимой склеренхимные отличаются большей упругостью, равной 15-20 кг/мм2,тогда как у колленхимы она составляет не более 10-12 кг/мм2. Наличиесклеренхимы дает возможность осевым органам растения противостоять нагрузкам наизгиб и удерживать кроны самих растений. Различают:
2.1. Первичнуюсклеренхиму – возникает из клеток основной меристемы, прокамбия или перицикла.
2.2. Вторичнуюсклеренхиму — возникает из клеток камбия.
Сами волокна– сильно вытянутые прозенхимные клетки с заостренными концами, в исключительныхслучаях достигают нескольких десятков сантиметров длины. Волокна, входящие всостав флоэмы (луба), носят название лубяных. Помимо луба, они встречаютсятакже в листовых черешках и пластинках, в цветоножках, плодоножках, реже вплодах. Волокна ксилемы (древесины) называются древесинными, или волокнамилибриформа. Они короче лубяных, и их стенки всегда одревесневают. Эволюционноволокна либриформа образовались из трахеид. У многих растений, обычно уоднодольных, волокна составляют механическую обкладку проводящих пучков.
В стебляхдвудольных волокна часто располагаются на месте перицикла и в первичной флоэме.В стеблях и листьях однодольных они образуют субэпидермальные тяжи, а в корняхсосредоточены главным образом в центральной части.
3. Склереиды– структурные элементы механической ткани, обычно возникают из клеток основнойпаренхимы в результате утолщения и лигнификации их оболочек. Склереиды могутвстречаться в виде скоплений либо располагаются поодиночке. По происхождениюони чаще первичные, т.е. произходят из различных первичных меристем. Клеткитипа склереид находятся в стеблях (хинное дерево), плодах (груша), семенах(многие бобовые). Считается, что функция склереид – противостоять сдавливанию,но иногда они защищают части растений от поедания животными.
Промышленноезначение имеют главным образом лубяные волокна стеблей двудольных и листовыеволокна крупных однодольных. Лубяные волокна некоторых двудольных в техникеназываются мягкими и используются преимущественно для изготовления различныхтканей (волокна льна, рами, кенафа), реже веревочно-канатных изделий (пенька,получаемая из конопли), а твердые волокна однодольных – почти исключительно дляизготовления веревок и канатов (новозеландский лен, волокна сизаля и др.).
2. АНАТОМИЧЕСКОЕ СТРОЕНИЕ ОСЕВЫХ ОРГАНОВ2.1 Анатомическое строение стебля двудольногорастения пучкового строения
Стебельпредставляет собой ось побега. Наряду с листом он является основной структурнойчастью побега. Основные функции – опорная и проводящая. Через стебельосуществляется связь между корнями и листьями и обмен продуктами воздушного иминерального питания. Кроме того, в стебле нередко откладываются запасныепитательные вещества.
Стебель, каки весь побег в целом, представляет собой «открытую» систему роста,т.е. он длительное время нарастает, и на нем возникают новые листья.
Стебель какчасть побега имеет систему меристем, поддерживающих нарастание тканей в длину итолщину. Рост в длину осуществляется за счет верхушечной и вставочных меристем,а в толщину у двудольных за счет боковых вторичных меристем – камбия ифеллогена.
Первичнаяструктура стебля складывается по мере дифференциации клеток верхушечноймеристемы побега. Верхушечная меристема побега двудольных довольно ранодифференцируется на несколько групп клеток, различающихся по особенностямделения и степени меристематической активности. Наружные ее слои преобразуютсяв протодерму, клетки которой позднее формируют первичную покровную ткань –эпидерму.
На уровнеоснований первых листовых примордиев клетки верхушечной меристемы,расположенные к периферии и в центре апекса, перестают активно делиться,увеличиваются в размерах и вакуолизируются. Из этих клеток формируютсяпервичная кора и сердцевина.
У многихдвудольных в инициальном кольце дифференцируется круг изолированных друг отдруга прокамбиальных тяжей. Клетки инициального кольца, расположенные междуэтими тяжами, дифференцируются позднее в паренхимные элементы. На поперечныхсрезах эти лучи имеют вид радиальных полос, соединяющих сердцевину с первичнойкорой.
Прокамбий уряда двудольных может закладываться также и в виде сплошного кольца. Он можетразвиваться по всей толще инициального кольца или формироваться из его части.Прокамбий является предшественником первичных проводящих тканей: первичнойфлоэмы и первичной ксилемы. Флоэма начинает формироваться раньше изакладывается в наружных частях прокамбиальных тяжей или прокамбиальногокольца. Флоэма развивается центростремительно, т.е. самые первые элементызанимают наружное положение, а самые поздние – внутреннее. Ксилемазакладывается во внутренних участках прокамбия и развивается центробежно. Такимобразом, флоэма и ксилема формируются навстречу друг другу. Первые элементыксилемы – проксилемы, узкие, сравнительно тонкостенные сосуды или трахеиды соспиральными или кольчатыми вторичными утолщениями. Метаксилема образуетсянесколько позднее протоксилемы и состоит из лестничных и пористых сосудов.Основным фактором, контролирующим дифференцировку как флоэмы, так и ксилемы,является фитогормон ауксин, вырабатываемый листовыми примордиями иперемещающийся по прокамбию от верхушки к основанию. Образовавшиеся изпрокамбия первичные флоэма и ксилема составляют основу осевого цилиндра, илистелы.
Стела,занимающая центральную часть стебля, состоит из проводящих тканей, сердцевины,перицикла и тех постоянных тканей, которые из него возникают. К наружи отперецикла располагается первичная кора, которая состоит из из паренхимы, нередкоколленхимы и иногда секреторных элементов.
Сердцевинаобычно состоит из относительно тонкостенных паренхимных клеток. В сердцевинечасто откладываются запасные питательные вещества. Здесь же нередко встречаютсяидиобласты, т.е. отдельные клетки, заполненные таннидами, кристаллами, слизью идр. Иногда часть сердцевины разрушается и образуется полость. Периферическаячасть сердцевины, примыкающая к ксилеме, называется перимедуллярной зоной.
Вторичноеутолщение стебля у большинства двудольных возникает довольно рано, что приводитк формированию вторичного тела растения. Эти изменения связаны главным образомс активностью боковой вторичной меристемы – камбия и отчасти другой вторичноймеристемы – феллогеном. За счет появления вторичных тканей осуществляется рострастений двудольных в толщину.
Вторичныеизменения в центральном цилиндре начинаются с заложения камбия. Камбийвозникает из остатков прокамбия, на границе первичных ксилемы и флоэмы.
Клеткикамбия сильно вакуолизированы и удлинены в вертикальном или в горизонтальномнаправлении. Первые – веретеновидные инициали дают проводящие элементыпроводящих тканей, вторые – лучевые инициали – образуют горизонтальноориентированные лучевые клетки сердцевинных лучей. Инициальные клетки камбияспособны к двум типам деления – периклинальному и антиклинальному. В первомслучае клеточная пластинка закладывается параллельно поверхности стебля, вовтором – перпендикулярно. В результате возникает непрерывный ряд производныхклеток, тянущихся от камбия радиально кнаружи и вовнутрь. Клетки,откладывающиеся в сторону наружной поверхности и стебля, постепеннодифференцируются во вторичную флоэму, в сторону сердцевины – во вторичнуюксилему.
Деятельностькамбия активизируется фитогормонами гиббериллином и ауксином, поступающими изпочек и молодых листьев.
В конечномитоге в структуре стебля однолетнего побега двудольного растения возможновыделить видоизмененный центральный цилиндр, включающий постоянные ткани,возникшие из перицикла, остатки первичной и вторичную флоэму, камбий, вторичнуюи остатки первичной ксилемы и сердцевину. Видоизмененный центральный цилиндрокружен остатками первичной коры.
2.2 Корень вторичного строения
Корень –основной орган высшего растения. Он осуществляет функцию минерального и водногопитания. Другая важная функция – закрепление, «заякоривание» растенияв почве.
Через кореньрастения поглощают из почвы воду и растворенные в ней ионы минеральных солей. Вкорнях осуществляется также биосинтез ряда вторичных метаболитов, в частностиалколоидов.
Кореньспособен к метаморфозам. Наиболее часто он является местом хранения запасныхпитательных веществ. Иногда выполняет роль дыхательного органа, может служитьорганом вегетативного размножения.
Образованиевторичной структуры корня связано прежде всего с деятельностью камбия, которыйобеспечивает рост корня в толщину. Камбий вначале возникает из тонкостенныхпаренхимных клеток в виде разобщенных участков с внутренней стороны тяжейфлоэмы между лучами первичной ксилемы. Камбиальную активность вскоре приобретаюти некоторые участки перицикла, располагающиеся кнаружи от лучей первичнойксилемы. В результате образуется непрерывный камбиальный слой.
К центрукамбий откладывает клетки вторичной ксилемы, а к периферии – клетки вторичнойфлоэмы.
Клетки камбия,заложившегося в перицикле, образуют широкие радиальные светлые лучи паренхимы,располагающиеся между тяжами вторичной проводящей ткани. Эти лучи, иногданазываемые первичными сердцевинными лучами, обеспечивают физиологическую связьцентральной части корня с первичной корой. Позднее могут закладываться ивторичные сердцевинные лучи, «связывающие» вторичную ксилему ифлоэму.
В результатедеятельности камбия первичная флоэма оттесняется кнаружи, а «звезда»первичной ксилемы остается в центре корня. Ее «лучи» сохраняютсядолго, иногда до конца жизни корня.
Помимовторичных изменений, происходящих в центральном цилиндре, существенные переменыпроисходят и в первичной коре. Вследствие быстрого нарастания изнутри вторичныхтканей, обусловливающего сильное утолщение корня, первичная кора нередкоразрывается. К этому времени клетки перицикла, делясь по всей окружностиосевого цилиндра, образуют широкую зону паренхимных клеток, во внешней частикоторой закладывается феллоген, откладывающий наружи пробку, а внутрьфеллодерму. Пробка изолирует первичную кору от проводящих тканей, кора отмираети сбрасывается. Клетки феллодермы и паренхима, сформированная за счетперицикла, в дальнейшем разрастаются и составляют паренхимную зону, окружающуюпроводящие ткани. Иногда эту зону называют вторичной корой. Снаружи корнидвудольных, имеющие вторичное строение, покрыты перидермой. Корка образуетсяредко, лишь на старых корнях деревьев.
3. ФОТОСИНТЕЗ
Фотосинтез –процесс образования органических веществ при участии энергии света – свойственлишь клеткам, содержащим специальные фотосинтезирующие пигменты, главнейшими изкоторых являются хлорофиллы. Это единственный процесс в биосфере, ведущий кзапасанию энергии за счет ее внешнего источника.
Ежегодно врезультате фотосинтеза на Земле образуется, как полагают, 150 млрд торганического вещества (первичная продукция) и выделяется около 200 млн тсвободного кислорода. Круговорот кислорода, углерода и других элементов,вовлекаемых в фотосинтез, поддерживает современный состав атмосферы,необходимый для существования на Земле современных форм жизни. Помимо«подпитки» атмосферы кислородом, фотосинтез препятствует увеличениюконцентрации CO2, предотвращая перегрев Земли вследствие так называемого парниковогоэффекта. Фотосинтез – главнейшее звено биохимических циклов на Земле и основавсех цепей питания. Запасенная в продуктах фотосинтеза энергия – основнойисточник энергии для человечества.
Существуютдва принципиально различных типа фотосинтеза:
1. Анаэробныйфотосинтез – свойствен немногим фотосинтезирующим бактериям. Фотосинтезирующимипигментами у них будут главным образом бактериохлорофиллы, в основе которых,как и хлорофиллов, лежит порфириновый скелет. Кислород в ходе анаэробногофотосинтеза не выделяется. Это обусловлено отсутствием фотосистемы II и тем,что донором электронов выступает не вода, а сера, сероводород или некоторыедругие органические соединения.
2. Аэробныйфотосинтез – важнейший для современных условий жизни на Земле. Он характерен длявсех оксифотобактерий, фотосинтезирующих протоктист и растений. Происходит онтолько в клетках, содержащих хлорофиллы. Чисто внешнее проявление этогопроцесса – выделение кислорода, поскольку донором электронов выступает вода.
Фотосинтез врастениях и фотосинтезирующих протоктистах осуществляется в хлоропластах. Уоксифотобактерий хлоропластов нет. Отдельная клетка у этой группы организмов визвестной мере соответствует отдельному хлоропласту. В такого рода организмахфотосистемы включены в соответствующие мембраны.
Фотосинтезвключает два главнейших этапа, последовательно связанных между собой. Этаппоглощения и преобразования энергии (явление, получившее название световогопроцесса) и этап превращения веществ (темновой процесс).
Световойпроцесс осуществляется в тилакоидах хлоропластов, темновой – главным образом вих строме.
Пигментырастений, участвующие в фотосинтезе, «упакованы» в тилакоидыхлоропластов в виде функциональных фотосинтетических единиц, называемыхфотосистемами. Имеется два типа фотосистем – I и II. Каждая фотосистемасодержит 250-400 молекул пигментов. Все пигменты фотосистемы могут поглощатьчастицы световой энергии, называемые фотонами или квантами света, но толькоодна молекула хлорофилла данной фотосистемы может использовать поглощеннуюэнергию в фотохимических реакциях. Эта молекула называется реакционным центромфотосистемы, а другие молекулы пигментов называются антенными, поскольку ониулавливают энергию света, подобно антеннам, для последующей передачиреакционному центру.
Вфотосистеме I реакционный центр образован особой молекулой хлорофилла a,обозначаемой как P700, где 700 – оптимум поглощения в нм. Реакционный центрфотосистемы II также образован молекулой хлорофилла a и обозначается индексомP680, поскольку оптимум поглощения лежит в районе 680 нм.
ФотосистемыI и II работают обычно синхронно и непрерывно, но фотосистема I можетфункционировать отдельно.
Все молекулыпигментов в фотосистемах способны улавливать энергию солнечного света. В случаеантенных пигментов свет, поглощенный молекулами, поднимает их электроны наболее высокий энергетический уровень, в конечном итоге высокоэнергетическиеэлектроны достигают реакционного центра. В случае фотосистемы II энергия светаутилизируется реакционным центром P680. Возбужденные энергизированные электроныцентра P680 парами переносятся на молекулу соединения, очевидно, относящегося кклассу хинонов и называемого акцептором. От акцептора начинается электронныйпоток, в котором электроны спускаются по электронотранспортной цепи кфотосистеме I. Компонентами этой цепи являются цитохромы – белки содержащиежелезо и серу, хиноны и белок пластоцианин, содержащий медь.Электронотранспортная цепь между фотосистемами I и II устроена так, что АТФможет образовываться из АДФ и Ф, причем этот процесс аналогичен окислительномуфосфорилированию, происходящему в митохондриях. В хлоропластах он связан сэнергией света и поэтому получил название фотофосфорилирование.
МолекулаP680, потерявшая свои электроны, заменяет их электронами донора. Как известно,таковым в аэробном фотосинтезе является вода. Когда электроны молекулы водыпоступают к P680, молекула диссоциирует на протоны и кислород. Этоокислительное расщепление молекул воды осуществляется под влиянием энергиисолнечного света – фотолиз. Ферменты, ответственные за фотолиз воды,располагаются на внутренней стороне мембраны тилакоидов. Таким образом, фотолизводы участвует в создании градиента протонов через мембрану, где высокаяконцентрация протонов оказывается во внутреннем пространстве тилакоидов.Протонный градиент способствует синтезу АТФ из АДФ и фосфата в ходефотофосфорилирования.
Вфотосистеме I энергия света «уловленная» антенными пигментамифотосистемы, поступает в реакционный центр P700. От P700 электроны передаютсяна электронный акцептор P430, который представляет белок, содержащий железо исеру. P430 передает свой электроны на другой железосеросодержащий белок –ферредоксин, а последний – на кофермент НАД, который восстанавливается до НАД*Н2. Молекула P700 в ходе процесса окисляется, но затем восстанавливаетпотерянные электроны за счет электронов, поступающих по электронотранспортнойцепи от фотосистемы II. Таким образом, на свету электроны перемещаются от водык фотосистемам II и I, а затем к НАД. Этот однонаправленный поток называетсянециклическим потоком электронов, а образование АТФ, которое при этомпроисходит, — нециклическим фотофосфорилированием.
ФотосистемаI может работать независимо от фотосистемы II. Этот процесс называютциклическим потоком электронов.
В ходе процессане происходит фотолиза воды, выделение O2 и образование НАД*Н2, однакообразуется АТФ.
Уэукариотических клеток циклическое фотофосфорилирование осуществляетсядостаточно редко, в тех случаях, когда клетка с избытком снабжаетсявосстановителем в форме НАД*Н2 извне, из других клеток или из другихкомпартментов клетки.
На второй(темновой) стадии фотосинтеза химическая энергия, запасенная в ходе световойреакции, используется для восстановления углерода. Углерод доступен дляфотосинтезирующих клеток в виде диоксида углерода, причем водоросли, багрянки иоксифотобактерии усваивают CO2, растворенный в воде. У растений диоксидуглерода поступает к фотосинтезирующим клеткам через устьица.
Восстановлениеуглерода происходит у эукариот в строме хлоропластов в цикле реакций – циклКальвина.
Исходноесоединение цикла Кальвина – пятиуглеродный сахар, фосфорилированный двумяфосфатными остатками – рибулозо-1,5-биофосфат (РБФ). CO2 входит в цикл ификсируется на РБФ. Образуемое при этом шестиуглеродное соединение затемрасщепляется на две молекулы 3-глицерофосфата, или 3-фосфоглицерата. Каждаямолекула 3-глицерофосфата содержит 3 атома углерода, в силу чего другоеназвание цикла Кальвина – C3-путь. Катализирует эти ключевые реакции ферментрибулозобисфосфаткарбоксилаза. Он располагается на поверхности тилакоидов. Втечение каждого оборота цикла одна молекула CO2 восстанавливается, а молекулаРБФ регенерируется и вновь может участвовать в следующем аналогичном цикле.
Шестиуглеродныйсахар глюкоза в конечном итоге образуется в результате шести оборотов цикла,которые ведут к «поглощению» 6 молекул CO2.
ЦиклКальвина — не единственный путь фиксации углерода в темновых реакциях. Унекоторых растений первый продукт фиксации CO2 – не трехуглеродная молекула3-глицерофосфата, а четырехуглеродное соединение – оксалоацетат. Отсюда этотпуть фотосинтеза получил название C4-путь. Оксалоацетат затем быстропревращается либо в малат, либо в аспартат, которые переносят CO2 к РБФ циклаКальвина. У C4-растений начальные этапы фотосинтеза осуществляются попреимуществу в клетках обкладок проводящих пучков, а C3-путь – в клеткахмезофилла. C4-растения используют оба пути фотосинтеза, но они в пределаходного растения пространственно разделены. C4-растения более экономно утилизируютCO2, чем C3-растения. C4-растения обладают способностью поглощать CO2 сминимальной потерей воды. Кроме того, у C4-растений практически отсутствуетфотодыхание – процесс выделения CO2 и поглощение O2 на свету.
Помимо C3 — и C4-путей, известен еще так называемый метаболизм органических кислот по типутолстянковых (САМ-метаболизм). Растения, фотосинтезирующие по САМ-типу, могутфиксировать CO2 в темноте с помощью фермента фосфоенолпируваткарбоксилазы,образуя яблочную кислоту, которая запасается в вакуолях. В течение последующегосветового периода яблочная кислота декарбоксилируется и CO2 присоединяется кРБФ цикла Кальвина в пределах той же клетки. САМ-метаболизм обнаружен у многихсуккулентных пустынных растений, у которых устьица открыты в ночное время изакрыты днем.
Каким быпутем ни осуществлялся фотосинтез, в конечном итоге он завершается накоплениемэнергетически богатых запасных веществ, составляющих основу для поддержанияжизнедеятельности клетки, а в итоге – всего многоклеточного организма. Эти веществаявляются продуктами первичного метаболизма. Помимо главнейшей своей функции,первичные метаболиты – основа для биосинтеза соединений, которые принятоназывать продуктами вторичного метаболизма. Следует заметить, что синтезвторичных метаболитов осуществляется за счет энергии, освобождающейся вмитохондриях в процессе клеточного дыхания.
4. КОМПЛЕКСЫ ТКАНЕЙ4.1 Понятие о флоэме
Термин«флоэма» ввел К.В. Негели в 1858 г. Флоэма – сложная проводящаяткань, по которой осуществляется транспорт продуктов фотосинтеза от листьев кместам их использования или отложения.
Первичнаяфлоэма дифференцируется из прокамбия, вторичная (луб) – производное камбия. Встеблях флоэма располагается обычно снаружи от ксилемы, а в листьях обращена книжней стороне пластинки. Первичная и вторичная флоэмы, помимо различноймощности ситовидных элементов, отличаются тем, что у первой отсутствуютсердцевинные лучи.
В составфлоэмы входят ситовидные элементы, паренхимные клетки, элементы сердцевинныхлучей и механические элементы. Большинство клеток нормально функционирующейфлоэмы живые. Отмирает лишь часть механических элементов. Собственно проводящуюфункцию осуществляют ситовидные элементы. Различают два их типа: ситовидныеклетки и ситовидные трубки. Стенки ситовидных элементов содержат многочисленныемелкие сквозные канальцы, собранные группами в так называемые ситовидные поля.У ситовидных клеток, вытянутых в длину и имеющих заостренные концы, ситовидныеполя располагаются главным образом на боковых стенках. Ситовидные клетки –основной проводящий элемент флоэмы у всех групп растений, исключаяпокрытосемянные. Клеток-спутниц у ситовидных клеток нет.
Ситовидныетрубки покрытосемянных более совершенны. Они состоят из отдельных клеток –члеников, располагающихся один над другим. Длина отдельных члеников ситовидныхтрубок колеблется в пределах 150-300 мкм. Поперечник – составляет 20-30 мкм.
Ситовидныеполя этих члеников находятся главным образом на их концах. Ситовидные поля двухрасположенных один над другим члеников образуют ситовидную пластинку.Ситовидные поля представляют собой участки клеточной стенки, пронизанныемногочисленными мелкими отверстиями, через которые с помощью цитоплазматическихтяжей сообщаются протопласты соседних ситовидных элементов.
Членикиситовидных трубок формируются из вытянутых клеток прокамбия или камбия. Приэтом материнская клетка меристемы делится в продольном направлении и производитдве клетки. Одна из них превращается в членик, другая – в клетку-спутницу.Наблюдается и поперечное деление клетки-спутницы с последующим образованиемдвух-трех подобных клеток, расположенных продольно одна над другой рядом счлеником. В клетках-спутницах вырабатываются различные ферменты, которыепередаются в ситовидные трубки. При своем формировании членик ситовидной трубкиимеет постенную цитоплазму, ядро и вакуоль. С началом функциональнойдеятельности он заметно вытягивается. На поперечных стенках появляетсямножество мелких отверстий-перфораций, образующих канальцы диаметром внесколько микрометров, через которые из членика в членик проходятцитоплазматические тяжи. На стенках канальцев откладывается полисахарид –каллоза, сужающий их просвет, но не прерывающий цитоплазматические тяжи.
По мереразвития членика ситовидной трубки в протопласте образуются слизевые тельца.Ядро и лейкопласты, как правило, растворяются. Граница между цитоплазмой ивакуолью – тонопласт – исчезает, и все живое содержимое сливается в единуюмассу. При этом цитоплазма теряет полупроницаемость и становится вполнепроницаемой для растворов органических и неорганических веществ. Слизевыетельца также теряют очертания, сливаются, образуя слизевый тяж и скопленияоколо ситовидных пластинок. На этом формирование членика ситовидной трубки завершается.Длительность функционирования ситовидных трубок невелика. У кустарников идеревьев она продолжается не более 3-4 лет. По мере старения ситовидные трубкизакупориваются каллозой и затем отмирают.
Основнымивеществами флоэмного тока являются сахара, главным образом сахароза. Крометого, обнаружены азотсодержащие вещества (аминокислоты), органические кислоты ифитогормоны.
Скоростьпередвижения ассимилятов во флоэме относительно невелика – 50-100 см/ч.
Паренхимныеэлементы флоэмы (лубяная паренхима) состоят из тонкостенных клеток. В нихоткладываются запасные питательные вещества, и отчасти по ним осуществляетсяближний транспорт ассимилятов.
Сердцевинныелучи флоэмы также состоят из тонкостенных паренхимных клеток. Они предназначеныдля осуществления ближнего транспорта ассимилятов.
4.2. Понятие о ксилеме
Термин«ксилема» ввел немецкий ботаник К.В. Негели. По ксилеме от корня клистьям передвигаются вода и растворенные в ней минеральные вещества. Первичнаяи вторичная ксилемы содержат клетки одних и тех же типов. Однако первичнаяксилема не формирует сердцевинных лучей. Первичная ксилема формируется изпрокамбия, вторичная – из камбия.
В составксилемы входят морфологически различные элементы, осуществляющие функции какпроведения, так и хранения запасных веществ, а также чисто опорные функции.Данный транспорт осуществляется по трахеальным элементам ксилемы – трахеидам исосудам, ближний в основном по паренхимным элементам. Дополнительные – опорную,а иногда и запасающую – функции выполняют трахеальные элементы и волокнамеханической ткани либриформа, также входящие в состав ксилемы.
Трахеиды взрелом состоянии – мертвые прозенхимные клетки, суженные на концах и лишенныепротопласта. Длина трахеид в среднем 1-4 мм, поперечник же не превышает десятыхи даже сотых долей миллиметра. Стенки трахеид одревесневают, утолщаются и несутпростые или окаймленные поры, через которые происходит фильтрация растворов.Боковые стенки трахеид в определенной степени водопроницаемы, что способствуетосуществлению ближнего транспорта. Большая часть окаймленных пор находитсяоколо окончаний клеток, т.е. там, где растворы «просачиваются» изодной трахеиды в другую. Трахеиды есть у спорофитов всех растений, а убольшинства хвощевидных, плауновидных, папоротниковидных и голосемянных ониявляются единственными проводящими элементами ксилемы.
Сосуды –полые трубки, состоящие из отдельных члеников, располагающихся друг над другом.
Междурасположенными один над другим члениками одного и того же сосуда имеютсяразного типа сквозные отверстия – перфорации. Благодаря перфорации вдоль всегососуда свободно осуществляется ток жидкости. Сосуды могут состоять из весьмазначительного числа члеников различной длины и диаметра. Общая же длина сосудовдостигает иногда нескольких метров. Диаметр же варьирует от 0,2 мм до 1 мм.Последнее зависит от вида растения, а у деревьев, растущих в сезонном климате,также и от того, в какой части ксилемы – «весенней» или«осенней» — сосуд расположен.
Помимопервичной оболочки, сосуды, как и многие трахеиды, в большинстве случаев имеютвторичные утолщения. В самых ранних трахеальных элементах вторичная оболочкаможет иметь форму колец, не связанных друг с другом. Затем следуют сосуды итрахеиды с утолщениями, которые могут быть охарактеризованы как спирали, виткикоторых связаны между собой. В конечном итоге вторичная оболочка сливается вболее или менее сплошной цилиндр, формирующийся внутрь от первичной оболочки.Сосуды и трахеиды с относительно небольшими округлыми участками первичнойклеточной оболочки, не прикрытыми изнутри вторичной оболочкой, нередко называютпористыми. В тех случаях, когда поры во вторичной оболочке образуют подобиесетки или лестницы, говорят о сетчатых или лестничных трахеальных элементах.
Вторичная, аиногда и первичная оболочки, как правило, лигнифицируется, т.е. пропитываютсялигнином, это придает дополнительную прочность, но ограничивает возможностьдальнейшего их роста в длину.
Трахеальныеэлементы, т.е. трахеиды и сосуды, распределяются в ксилеме различным образом.Иногда на поперечном срезе они образуют хорошо выраженные кольца. В другихслучаях сосуды рассеяны более или менее равномерно по всей массе ксилемы.
Помимотрахеальных элементов, ксилема включает лучевые элементы, т.е. клетки, образующиесердцевинные лучи, сформированные чаще всего тонкостенными клетками. Реже,например, в лучах хвойных, встречаются лучевые трахеиды. По сердцевинным лучамосуществляется ближний транспорт веществ в горизонтальном направлении.
В ксилемепокрытосемянных, помимо проводящих элементов, содержатся также тонкостенныенеодревесневшие живые паренхимные клетки – древесинная паренхима. По нимотчасти наряду с сердцевинными лучами осуществляется ближний транспорт. Крометого, древесинная паренхима служит местом хранения запасных веществ. Элементысердцевинных лучей и древесинной паренхимы, подобно трахеальным элементам,возникают из камбия, но из паренхимных инициалей.
Клеткипаренхимы, примыкающие к сосудам, могут образовывать выросты в полость сосудачерез поры, так называемые тилы. Иногда тилы заполняют всю полость сосуда, и вэтом случае проводящая функция нарушается. Тилообразование усиливаетмеханическую прочность центральной части стволов деревьев. Кроме того, тилыиграют особую роль в процессе формирования ядра древесины.
5. ХАРАКТЕРИСТИКА СЕМЕЙСТВ: РОЗОЦВЕТНЫЕ И ЯСНОТКОВЫЕ
Розоцветные
Rosaceae
Яснотковые
Lamiaceae
Состав Жирное масло, цианогенные гликозиды, Тритерпеновые сапонины, танниды, терпеноиды, полисахариды, редко алколоиды, кумарины. Эфирные масла, ди — и тритерпеноиды, сапонины, полифенолы и танниды, иридоиды, хиноны, кумарины, гормоны линьки насекомых, редко алколоиды. Листья Очередные, редко супротивные, простые или сложные, с прилистниками. Супротивные Соцветия Цимоидные или ботриодные(в виде кисти) Цветки Правильные, обоеполые, нередко довольно крупные Правильные, обоеполые Околоцветник Двойной Двойной Чашечка Из 5, редко 4 сросшихся чашелистиков, иногда снабженная подчашием Пятизубчатая, двугубая, правильная или неправильная Лепестки В числе 5 или 4, свободные, прикрепленные, подобно тычинкам Андроцей Из большого числа расположенных в несколько рядов тычинок, иногда число тычинок уменьшается до 4-8-9 Тычинок 4, прикрепленных к трубке венчика Гинецей Апокарпный или ценокарпный Ценокарпный Число плодолистиков Число может быть различно от большого и неопределенного числа до 1 – два семязачатка 2 плодолистика, делящиеся, пополам продольной перегородкой Завязь Верхняя или нижняя Четырехгнездная, четырехлопостная Плоды Многолистовка, многокостянка и многоорешек Ценобий, распадающийся на 4 доли Семена Без эндосперма, зародыш прямой Без эндосперма
СПИСОК ИСПОЛЬЗУЕМОЙ ЛИТЕРАТУРЫ
1. Яковлев Г.П., Челомбитько В.А. Ботаника.М., 1990
2. Бавтуто Г.А., Еремин В.М. Ботаника: Морфология и анатомия растений.Минск, 1997
3. Рейвн П., Эверт Р., Айкхорн С. Современная ботаника.М., 1990
4. Чебышев Н.В., Гринева Г.Г., Кобзарь М.В., Гулянков С.И. Биология.М., 2000
5. Эсау К. Анатомия семенных растений.М., 1980