Skip to content
- Главная
- Университеты
- БашГМУ
- Теория
- Гистология, теория
Башкирский государственный медицинский университет
- Лекции
- Теория для экзамена
Лекции
Теория для экзамена
Перед отправкой формы внимательно прочитайте то, что находится под каждым из них. В противном случае, ваше обращение может не дойти до нас.
Ответы на экзаменационные вопросы по гистологии
№1
Определение
и задачи гистологии
Гистология
(от греч.Histos
– ткань, logos
– учеяние) – наука о строении, развитии
и жизнедеятельности тканей животных
организмов. Гистология вместе с другими
фундаментальными медико-биологическими
науками изучает закономерности
структурной организации живой материи,
является одним из подразделений науки
о жизни – биологии. Гистологию
характеризуют 3 признака : 1- материя,
которую она изучает, 2- методы, которые
она использует, 3-законы,которые она
познает. Материалом для гистологии
являются ткани животных и человека,
клетки, из которых состоят ткани и
органы, сформированные из тканей.
Гистология исследует филогенез и
онтогенез тканей. По своему фундаментальному
содержанию гистология рассматривается
как фундаментальная наука.
Актуальными
задачами гистологии являются:
—
разработка общей теории гистологии,
отражающей эволюционную динамику тканей
и закономерности эмбрионального и
постнатального гистогенеза;
—
изучение гистогенеза как комплекса
координированных во времени и пространстве
процессов пролиферации, дифференциации,
детерминации, интеграции, адаптивной
изменчивости, программированной гибели
клеток и др.;
—
выяснение механизмов гомеостаза и
тканевой регуляции (нервной, эндокринной,
иммунной), а также возрастной динамики
тканей;
—
изучение закономерностей реактивности
и адаптивной изменчивости клеток и
тканей при действии неблагоприятных
экологических факторов и в экстремальных
условиях функционирования и развития,
а также при трансплантации;
—
разработка проблемы регенерации тканей
после повреждающих воздействий и методов
тканевой заместительной терапии;
—
раскрытие механизмов молекулярно-генетической
регуляции клеточной дифференцировки,
наследования генетического дефекта
развития систем человека, разработка
методов генной терапии и трансплантации
стволовых эмбриональных клеток;
—
выяснение процессов эмбрионального
развития человека, критических периодов
развития, воспроизводства и причин
бесплодия.
№2
Основные
периоды исторического развития
гистологии.
1
период:
Накопление
первоначальных фактов (1595-1800г), изобретение
Янсоном первого микроскопа. Микроскоп
увеличивал не более чем в 20 раз, вследствие
чего не использовался по назначению.
Для исследований использовались
одиночные линзы, увеличивающие до 200
раз. Антуан ванн Левенгук изобрел линзы,
увеличивающие в 300 раз. Именно он стал
основоположником научной микроскопии.
Левенгук в капле воды обнаружил много
живых существ. Гук, будучи физиком,
изготовил более совершенный микроскоп
и впервые ввел понятие «Клетка» в 1665
году. Грю, будучи ботаником, изучал части
растений, кору, листья, цветы. Систему
однородных элементов он назвал тканью
в 1682 году.
2
период:
В
1800г Франсуа Ксавье, не используя
микроскоп, методом мацерации (настаивания),
выделил 21 вид тканей и заложил основы
гистологии под названием «Микроскопическая
анатомия». Майер (1818) и Хойзингер (1821)
переименовывают ее в гистологию. В
1838-1839 Шванн формирует клеточную теорию.
3
период:
Период
борьбы за господство клеточной теории.
Под ее влиянием бурно развивается
гистология, и на основе ее цитология,
гематология, эндокринология, иммунология,
патологическая анатомия. Клеточная
теория оказала значительное воздействие
на общественную жизнь людей того времени.
4
период:
В
1945г. Портер и соавторы, исследуя под
электронным микроскопом фибробласты,
открывают ЭПС и хоронят идею о цитоплазме,
как о бесструктурной системе. Новые
знания о строении клеток были дополнены
информацией об их функции.
№3
Гистология
как учебная дисциплина, ее содержание.
Как учебная
дисциплина гистология включает
несколько разделов: 1) цитологию — учение
о клетке; 2) эмбриологию — науку о развитии
зародыша, закономерностях закладки и
образования тканей и органов; 3) общую
гистологию — учение о развитии, структуре
и функциях тканей; 4) частную гистологию,
изучающую микроскопическое строение
органов и систем органов.
Цитоло́гия (греч. κύτος —
«вместилище», здесь: «клетка» и λόγος —
«учение», «наука») — раздел биологии,
изучающий живые клетки,
их органоиды,
их строение, функционирование, процессы
клеточного размножения, старения и
смерти.
Эмбриология (от
древнегреческого ἔμβρυον, зародыш,
«эмбрион»; и -λογία, -логия)
— это наука, изучающая развитие
зародыша. Зародышемназывают
любой организм на
ранних стадиях развития до рождения или
вылупления, или, в случае растений, до
момента прорастания.
Многими учёными, в том числе
отечественными, эмбриология определяется
более широко, как синоним биологии
развития .
Гистология (от
греч. histos —
ткань, logos —
учение) — наука о строении, развитии и
жизнедеятельности тканей животных
организмов.
Частная гистология служит
основой для изучения микроскопического строения
морфофункциональных единиц органов и
органов в целом.
№4
Клеточная
теория – теоретическая фундаментальная
основа гистологии.
Клеточная
теория —
одно из общепризнанных биологических обобщений,
утверждающих единство принципа строения
и развития мира растений, животных и
остальных живых организмов с клеточным
строением,
в котором клетка рассматривается в
качестве общего структурного элемента
живых организмов.
Клеточная
теория — основополагающая для
биологии теория, сформулированная в
середине XIX
века, предоставившая
базу для понимания закономерностей
живого мира и для развития эволюционного
учения. Маттиас
Шлейден и Теодор
Шванн сформулировали клеточную
теорию, основываясь
на множестве исследований
о клетке (1838). Рудольф
Вирхов позднее
(1858)
дополнил её важнейшим положением (всякая
клетка происходит от другой клетки).
Шлейден
и Шванн, обобщив имеющиеся знания о
клетке, доказали, что клетка является
основной единицей любого организма.
Клетки животных, растений и бактерии имеют
схожее строение. Позднее эти заключения
стали основой для доказательства
единства организмов. Т. Шванн и
М. Шлейден ввели в науку основополагающее
представление о клетке: вне клеток нет
жизни. Клеточная теория дополнялась и
редактировалась с каждым разом.
Основные
положения клеточной теории:
1)
Клетка – единица строения, жизнедеятельности,
роста и развития живых организмов, вне
клетки жизни нет.
2)
Ядро – главная составная часть клетки
эукариот.
3)
Новые клетки образуются только в
результате деления исходных клеток.
4)
Клетка – единая система, состоящая из
множества закономерно связанных друг
с другом элементов, представляющих
собой определенное целостное образование.
5)
Клетки многоклеточных организмов
образуют ткани, ткани образуют органы.
Жизнь организма в целом обусловлена
взаимодействием составляющих клеток.
№5
Симпласт
и синцитий как формы организации
протоплазмы.
Симпласт
– крупное образование, состоящее из
цитоплазмы с множеством ядер. Примерами
симпластов могут быть мышечные волокна
позвоночных, наружный слой трофобласта
плаценты. Симпласты возникают вторично
за счет слияния отдельных клеток, или
же в результате деления одних ядер без
разделения цитоплазмы , без цитотомии.
Синцитий
(соклетия) – первичная надклеточная
форма организации жизни, представляющая
собой протоплазматическую решетку, в
узлах которой лежат ядра. У человека
синцитиально связанные между собой
клетки сохранились в семеннике, где эти
связи синхронизируют развитие
сперматоцитов.
№6
Характеристика
межклеточного вещества.
Межклеточное
вещество – продукт жизнедеятельности
определенных групп клеток.Составная
часть соединительной ткани позвоночных
и многих беспозвоночных животных,
включающая соединительнотканные волокна
и аморфное основное вещество, выполняющая
механическую, опорную, защитную и
трофическую функции.
Межклеточное
вещество образуется у зародыша из
белков, углеводов, липидов, продуцируемых
клетками эмбриональной соединительной
ткани, начиная со стадии гаструлы.
Гистогенез межклеточного вещества
продолжается и в постэмбриональном
периоде. Наибольшая роль в образовании
межклеточного вещества принадлежит
фибробластам, хондробластам, остеобластам.
Полагают, что в образовании межклеточного
вещества волокнистой соединительной
ткани могут участвовать гистиоциты,
лаброциты (тучные клетки) и другие.
Соединительнотканные
волокна межклеточного вещества могут
быть представлены коллагеновыми,
эластическими, ретикулярными, или
ретикулиновыми (аргирофильными), и
другими волокнами, от чего зависит
прочность, эластичность и в определенной
степени архитектоника соединительной
ткани органов (дерма различных участков
кожи, сухожилия, строма кроветворных
органов и так далее).
Аморфное
основное вещество, окружающее
соединительнотканные волокна и клетки
соединительной ткани, состоит из
высокополимерных соединений, от
концентрации и состава которых в
различных видах соединительной ткани
зависят физические, химические и
биологические свойства межклеточного
вещества (вязкость, гидрофильность,
интенсивность метаболических процессов,
тургор и другие).
Состав
волокон и аморфного вещества неодинаков
в различных видах соединительной ткани,
в различных ее топографических участках
межклеточное вещество может быть
минерализованным. При этом кристаллы
минералов (фосфорнокислый кальций,
углекислый кальций и другие) импрегнируют
органическую основу межклеточного
вещества твердых скелетных тканей
(дентин, кость). С возрастом межклеточное
вещество претерпевает инволюционные
изменения: меняется соотношение основного
вещества и волокон — масса волокнистых
структур коллагена и плотность его
«упаковки» возрастают, а масса основного
вещества уменьшается, происходят
конденсация эластических волокон,
глубокие физико-химические изменения
межклеточного вещества.
В
эксперименте на животных выявлено, что
недостаточное питание задерживает
развитие возрастных изменений коллагена,
а «атерогенная» диета вызывает его
постарение.
Характером
строения межклеточного вещества в
значительной мере определяются
функциональными особенности тех или
иных видов соединительной ткани. Чем
плотнее межклеточное вещество, тем
сильнее выражена механическая, опорная
функция, которая достигает наибольшего
развития в костной ткани. Трофическая
функция, напротив, лучше обеспечивается
полужидким по консистенции межклеточным
веществом (интерстициальная соединительная
ткань, окружающая кровеносные сосуды).
Коллагеновые
и эластические волокна, входящие в
состав межклеточного вещества, построены
из склеропротеинов — коллагена и
эластина. Из коллагена состоят и
ретикулиновые волокна отличающиеся
повышенным содержанием углеводов и
наличием липидов. В эластических волокнах
имеется микрофибриллярный компонент,
отличный от эластина по аминокислотному
составу. Этот же компонент образует
особую разновидность немногочисленных,
сходных с эластическими волокнами
межклеточного веществ (окситалановых),
волокон резистентных к действию эластазы.
Свойства основного вещества определяются
преимущественно углеводно-белковыми
биополимерами — гликозаминогликанами
и гликопротеидами. Наличие гликозаминогликанов
придает основному веществу межклеточного
вещества выраженную базофильность.
Качественные и количественные соотношения
этих биополимеров, отличающихся
интенсивным метаболизмом, различны в
разных видах соединительной ткани.
№7
Клетка
– главная форма организации протоплазмы.
Клетка
– ограниченная активной мембраной ,
упорядоченная система биополимеров,
образующих ядро и цитоплазму, участвующих
в единой совокупности метаболических
и энергетических процессов, осуществляющих
поддержание и воспроизведение всей
системы в целом. В своем историческом
развитии протоплазма приобретает
разнообразные формы, среди которых
различают первичные и вторичные.
Первичной формой протоплазмы является
эукариотическая клетка многоклеточных.
Это главная, исторически сложившаяся
форма организации живой материи,
обладающая всеми основными свойствами
жизни, имеющая ядро, цитоплазму и
цитоплазматические органеллы. Вторичные
формы – весь многоклеточный мир.
№
8
Величина
и форма клеток. Факторы их обуславливающие.
Величина
клетки определяется
ядерно-цитоплазматическими отношениями
и отношением площади поверхности к
объему цитоплазмы, которые должны быть
постоянными. Контактное торможение,
которое определяет положение и
пространство, занимаемое клеткой.
Смещение константы ведет либо к делению
клетки, либо к ее гибели.
Форма
клетки:
Веретеновидная
форма
– клетка имеет утолщенную среднюю
часть, брюшко, где лежит ядро и основные
органеллы и 2 конца, фиксирующие к
субстрату. (миоциты внутренних полых
органов). Клетка
призматической формы
находится на поверхности органов , несут
защитную функцию, приближены друг к
другу (эпителиальные клетки). Клетки
шаровидной
формы – расположены не плотно и находятся
в жидкости (лейкоциты). Клетки
звездчатой формы
(нейроны).
Между
формой и содержанием, структурой и
функцией имеется диалектическое
взаимодействие.
Основными
структурными компонентами клетки
являются:
1)
клеточная поверхность (надмембранный
комплекс, плазматическая мембрана,
подмембранный комплекс);
2)
цитоплазма (гиалоплазма, органеллы и
включения);
3)
ядро (кариолемма, ядрышко, хроматин,
кариолимфа).
№9
Классификация
цитоплазматических органелл.
|
2.1.2. |
|
А. Мембранные органеллы
1.
а) эндоплазматическая
б) комплекс (аппарат) Гольджи
в) эндосомы и лизосомы
г) пероксисомы —
2.
Б. Немембранные
Их
1.
2.
I. сократительные структуры
II.
а) микрофиламенты (МФ)
б) промежуточные
в) микротрубочки (МТ)
3.
а)
б)
I. II. |
№10
Клеточная
поверхность и ее функции
Клеточная
поверхность выполняет
следующие функции: разграничительная,
барьерно-защитная, рецепторная,
транспортная, контактная, опорно-механическая,
двигательная. Ее основными химическими
компонентами являются: липиды (40%), белки
(50%) и углеводы (10%). Соотношение этих
веществ может варьировать в зависимости
от функциональной активности клетки.
Надмембранный
комплекс – это гликокаликс клетки,
образован молекулами олигосахаридов,
связанных с интегральными и покровными
белками плазмолеммы. В состав гликокаликса
входят: мальтоза, глюкоза, галактоза.
Они образуют над плазмолеммой
гетерополиморфные ветвящиеся цепочки.
Гликокаликс осуществляет сортировку
макромолекул из межклеточной среды, их
удержание.
Плазматическая
мембрана – образована слоем гидрофильных
и гидрофобных липидов. Между липидами
вставлены молекулы интегральных и
полуинтегральных белков. Снаружи
липидного комплекса лежат покровные
белки. Плазматическая мембрана имеет
каналы для натрия, калия, кальция, хлора
и рецепторы к медиаторам, гормонам и
цитокинам.
Подмембранный
комплекс – слой глобулярных белков
цитозоля и фиксированных к плазмолемме
концов микротрубочек и филаментов
клеточного скелета.
Производные
клеточной поверхности: микроворсинки,
реснички и жгутики.
Транспорт:
1)Простая
диффузия – перемещение веществ по
градиенту концентрации.
2)Облегченная
диффузия — пассивный транспорт воды из
клетки в клетку через специальные
каналы.
3)Активный
транспорт – перенос веществ с затратой
энергии.
4)Экзо-
и эндоцитоз.
№11
Основные
функции клетки.
1)Деление
клетки –вид размножения клеток. Наиболее
распространенным способом деления
соматической клетки является митоз. Во
время митоза клетка проходит ряд
последовательных стадий, в результате
которых каждая дочерняя клетка получает
такой же набор хромосом, как и у материнской
клетки.
2)Обмен
веществ. Основная функция клетки. Из
межклеточного вещества в клетку постоянно
поступают питательные вещества и
кислород, и выделяются продукты распада.
Обмен веществ выполняет 2 функции:
обеспечение клетки строительным
материалом. Из веществ, поступающих в
клетку, непрерывно происходит биосинтез
белков, углеводов, липидов, из которых
впоследствии формируются органеллы.
Вторая функция – это обеспечение клетки
энергией. (Энергетический и пластический
обмен).
3)Раздражимость
– реакция на физические и химические
воздействия окружающей среды. В
возбужденном состоянии разные клетки
выполняют свойственные им функции.
№12
Синтетический
аппарат клетки.
Синтетический
аппарат клетки обеспечивает
синтезы различных веществ и включает
ЭПС, кГ и рибосомы.
Рибосомы –
округлые мелкие, немембранные органеллы,
состоящие из двух округлых субъединиц
– малой и большой. Каждая субъединица
образованы рибосомальной РНК и сложным
набором белков. Синтез рРНК и сборка
субъединиц происходит в ядрышке, а их
объединение – уже в цитоплазме. Рибосомы
обеспечивают процесс трансляции белка.
Малая субъединица связывается с иРНК,
а большая катализирует образование
пептидных связей между аминокислотами.
Одиночные
рибосомы неактивны и для белкового
синтеза они объединяются в цепочки,
нанизываясь на молекулу иРНК. Так
образуются полисомы. Свободные
полисомы синтезируют белки, которые
диффузно распределяются в гиалоплазме.
Синтез
мембранных белков, лизосомальных белков
и секреторных белков, которые будут
выведены за пределы клетки, осуществляют
полисомы, прикрепленные к ЭПС.
При
синтезе секреторных и лизосомальныз
белков используется особый механизм,
который позволяет полипептидной цепи
проникать внутрь полости ЭПС. После
завершения синтеза молекула приобретает
вторичную и третичную структуру и выйти
наружу уже не может. Синтез таких белков
начинается с особого участка – сигнального
пептида. Благодаря ему рибосома
соединяется с рибофорином – белком,
который встроен в мембрану ЭПС. В
присоединении участвует еще и специальный
мембранный рецептор ЭПС. После
присоединения рибосомы рибофорин
приобретает форму канала, через который
проходит синтезируемая полипептидная
цепь. Когда белковая молекула готова,
сигнальный участок отсоединяется.
Если
синтезируется мембранный белок, то в
полипептидной цепи оказывается еще
один участок, который заякоривает
белковую цепь в мембране.
ЭПС
представляет собой сложную систему
мембранных полостей. Обычно в форме
плоских цистерн, распределенных по всей
клетке.
Есть
два типа ЭПС
– гранулярная и агранулярная.
К поверхности грЭПС прикрепляются
полисомы.
Итак, главные
функции грЭПС:
синтез, химическая модификация, накопление
и транспортировка белков.
Агранулярная
ЭПС является
продолжением грЭПС, но лишена я
ЭПС является
продолжением грЭПС, но лишена рибосом
и имеет иной набор белков-ферментов.
аЭПС – трубчатыми каналами. У нее
множество функций:
-
Синтез
липидов и холестерина, поэтому ее много
в клетках, синтезирующих стероидные
гормоны и жиры. -
Синтез
гликогена (клетки печени) -
Детоксикация
вредных веществ (лекарственные препараты,
алкоголь, токсины) -
Накопление
Са2+,
необходимого для сокращения мышечных
клеток.
От
ЭПС отшнуровываются транспортные
пузырьки, содержащие синтезированные
вещества, перемещаются в сторону
комплекса Гольджи и сливаются с ним.
Комплекс
Гольджи –
мембранная органелла, представленная
диктиосомами (стопка из 3-10 плоских
цистерн). Диктиосома имеет незрелую
поверхность, обращенную к ЭПС (цис-) и
зрелую, обращенную к плазмолемме
(транс-). С цис-поверхностью сливаются
транспортные мембранные пузырьки,
содержащие продукты синтеза, которые
отшнуровываются от ЭПС. Вещества,
попавшие в полости кГ, направляются в
различные части диктиосомы, где
подвергаются процессингу. Это химические
превращения молекул – к ним могут
присоединяются сахара, сульфатные и
фосфатные группы, белковые молекулы
могут частично расщепляться и т.д.
От
боковых участков кГ отшнуровываются
гидролазные пузырьки, заполненные
гидролитическими ферментами. Из них
формируются лизосомы.
Функции
кГ:
-
Синтез
полисахаридов и гликопротеинов (слизь,
гликокаликс). -
Процессинг
молекул -
Накопление
продуктов синтеза, их упаковка и
транспортировка. -
Формирование
лизосом.
,
№13
ГЭРЛ-система
и поток мембран в клетке.
ГЭРЛ
(Г — Гольджи, Р- эндоплазм.ретикулом,
Л -лизосомы) — —
система и поток мембран в клетке:
синтетический и транспортный компармент
клетки, связанный с потоком мембран
клеток. Чем активней клетка, тем активней
поток. Мембранный транспорт, или
мембранный поток («membrane flow»), является
необходимым для обмена веществ между
различными клеточными органеллами. В
настоящее время для описания этого
процесса принята следующая модель:
1)
Внутриклеточный транспорт является
направленным; транспорт от эндоплазматического
ретикулума через аппарат
Гольджи к
лизосомам, эндосомам или клеточной
поверхности называется антероградным;
транспорт в обратном направлении —
ретроградным .
2)
Транспортный путь проходит через ряд
отдельных компартментов, перенос
вещества между которыми осуществляется
преимущественно с помощью окаймленных
мембранных пузырьков . В клетке существует
несколько типов окаймляющих белков и,
соответственно, несколько типов
окаймленных пузырьков, курсирующих
между определенными компартментами.
3)
Не все органеллы способны к прямому
обмену пузырьками, например, транспорт
между эндоплазматическим ретикулумом
и эндосомами, лизосомами и плазматической
мембраной может быть осуществлен только
через аппарат Гольджи .
4)
Процесс, в результате которого нужные
вещества направляются в соответствующие
компартменты, называют сортингом
(«sorting»).
№14
Митохондрии
и их энергетические функции.
Митохондрии
– органеллы синтеза АТФ. Их основная
функция связана с окислением органических
соединений и использованием освобождающейся
при распаде этих соединений энергии
для синтеза молекул АТФ.
Форма
и размеры митохондрий митохондрий
животных клеток разнообразны и их
количество в клетках варьирует от единиц
до сотен.
Основная
функция митохондрий – синтез АТФ,
происходящий в результате процессов
окисления органических субстратов и
формирование АДФ. Начальные этапы этих
сложных процессов совершаются в
гиалоплазме. Здесь происходит первичное
окисление субстратов (например сахаров
до ПВК с одновременным синтезом небольшого
количества АТФ. Эти процессы совершаются
при отсутствии кислорода. Все последующие
выработки энергии (дыхание) – анаэробное
окисление и синтез основной массы АТФ
осуществляется с потребление кислорода
и локализ. внутри митохондрий. При этом
происходит дальнейшее окисление пирувата
и других субстратов энергетического
обмена с выделение углекислого газа и
переносом протонов на их акцепторы. Эти
реакции осуществляются с помощтю ряда
ферментов ЦТК, которые локализированы
в матриксе митохондрий.
В
мембранах крист митохондрий располагаются
системы дальнейшего переноса электронов
и сопряженного с ним фосфорилирования
АДФ. При этом происходит перенос
электронов с одного белка-акцептора
электронов к другому и,наконец, связывание
их с кислородом, вследствие чего
образуется вода. Одновременно с этим
часть энергии, выделяемой при таком
окислении в цепи переноса электронов
запасается в виде макроэргической связи
при фосфорилировании АДФ, что приводит
к образованию большого количества
молекул АТФ. Именно на мембранах крист
митохондрий происходит процесс
окислительного фосыорилирования с
помощью здесь расположенных белков
цепи окисления и ферментов фосфорилирования
АДФ, АТФ-синтетазы.
№15
Пищеварительный
аппарат клетки – лизосомы.
Класс
шаровидных структур, ограниченных
одиночной мембраной. Характерным
признаком является наличие в них
гидролитических ферментов – гидролаз
(протеинкиназы, липазы), расщепляющих
различные биополимеры. Можно выделить
3 типа лизосом: первичные, вторичные и
остаточные тельца.
Первичные
лизосомы – мелкие мембранные пузырьки,
заполненные бесструктурным веществом,
содержащим гидролазы, в т.ч. и фосфатазу,
которая является маркерным лизосомным
ферментом. Местом синтеза фосфатазы
является гранулярная ЭПС, затем этот
фермент появляется в проксимальных
участках диктиосом, а затем в мелких
везикулах по периферии диктиосом и,
наконец, в первичных лизосомах. Вторичные
лизосомы – вторичные пищеварительные
вакуоли, формируются при слиянии
первичных лизосом с фагосомами
(фагоцитарными вакуолями) или пиноцитозными
вакуолями, образуя гетерофагосомы.
Вещества, попавшие в состав вторичной
лизосомы, расщепляются гидролазами до
мономеров, которые транспортируются
через мембрану лизосом в гиалоплазму
где они включаются в обменные процссы.
Расщепление, переваривание биогенных
молекул внутри лизосом может идти в
ряде клеток не до конца. В этом случае
в полостях накапливаются непереваренные
продукты. Такая лизосома называется
остаточным тельцем.
№16
Регуляция
синтеза белка в клетке
Биосинтез
белка – многостадийный процесс синтеза
полипептидной цепи из а/к остатков,
происходящий на рибосомах клеток живых
организмов с участием молекул мРНК и
тРНК.
Биосинтез
можно разделить на 3 стадии: транскрипция,
процессинг и трансляция.
Во
время транскрипции
происходит считывание генетической
информации, зашифрованной в молекулах
ДНК, и запись этой информации в молекулы
и РНК. В ходе ряда последовательных
стадий процессинга из мРНК удаляются
некоторые фрагменты, ненужные в
последующих стадиях, и происходит
редактирование нуклеотидных
последовательностей. После транспортировки
когда из ядра к рибосомам происходит
собственно синтез белковых молекул
путем присоединения отдельных а/к
остатков к растущей полипептидной цепи.
Процессинг
РНК: Между
транскрипцией и трансляцией молекула
мРНК претерпевает ряд изменений, которые
обеспечивают созревание функционирование
матрицы для синтеза полипептидной
цепочки. К 5’ концу присоединяется кэп
(модифицированный гуаниновый нуклеотид,
который добавляется на 5’ (передний)
конец незрелой матричной рибонуклеиновой
кислоты), а к 3’ концу поли – А- хвост,
который увеличивает длительность жизни
иРНК. С появлением процессинга в
эукариотической клетке стало возможно
комбинирование экзонов гена для получения
большего разнообразия белков, кодируемых
единой последовательностью нуклеотидов
ДНК.
Трансляция:
заключается
в синтезе полипептидной цепи в соответствии
с информацией, закодированной в мРНК.
А/к последовательность выстраивается
при помощи тРНК, которые образуют
комплексы с а/к. Каждой а/к соответствует
своя тРНК, имеющая соответствующий
антикодон, «подходящий» к кодону мРНК.
Во время трансляции рибосома движется
вдоль мРНК, по мере этого наращивается
полипептидная цепь. Энергией биосинтез
белка обеспечивается за счет АТФ. Готовая
белковая молекула затем отщепляется
от рибосомы и транспортируется в нужное
место клетки. Для достижения своего
активного состояния некоторые белки
требуют дополнительной посттрансляционной
модификации.
№17
Строение
и функции ядра
Ядро
– система генетической детерминации
и регуляции белкового синтеза.
Функции
ядра:Хранение
и поддержание наследственной информации
в виде неизменной структуры ДНК
. В ядре происходит воспроизведение и
редупликация молекул ДНК, что дает
возможность при митозе двум дочерним
клеткам получить совершенно одинаковые
в качественном и количественном отношении
объемы генетической информации. Вторая
функция – создание аппарата белкового
синтеза. Это
не только синтез, транскрипция
на молекулах ДНК разных иРНК, но и
транскрипция
всех
видов тРНК и рРНК. В ядре происходит
образование субъединиц рибосом.
Структура
ядра:Ядрышко
– имеет величину от 1-5 мкм, очень богаты
РНК, базофильны, не является самостоятельной
структурой или орагнеллой. Ядрышко
является местом образования рРНК и
рибосом, на которых происходит синтез
полипептидных цепей в цитоплазме.
Образование ядрышек и их число связаны
с активностью и числом определенных
участков хромосом – ядрышковых
организаторов, которые расположены
большей частью в зонах вторичных
перетяжек. Вокруг ядрышек есть зона
конденсированного хроматина. Схема
участия ядрышек в синтезе цитоплазматических
белков: на ДНК ядрышкового организатора
образуется предшественник рРНК, который
в зоне ядрышка одевается белком, здесь
происходит сборка рибонуклеопротеидных
частиц – субъединиц рибосом; субъединицы,
выходя из ядрышка, участвуют в синтезе
белка. Ядерная
оболочка – состоит
из внешней ядерной мембраны и внутренней
мембраны оболочки, разделенных
перинуклеарным пространством или
цистерной ядерной оболочки. Ядерная
оболочка содержит ядерные поры. Внешняя
мембрана ядерной оболочки непосредственно
контактирующая с цитоплазмой клетки,
имеет ряд структурных особенностей,
позволяющих отнести ее к собственно
мембранной системе ЭПС: на ней со стороны
гиалоплазмы расположены многочисленные
полирибосомы , а сама внешняя ядерная
мембрана может прямо переходить в
мембраны ЭПС. Внутренняя мембрана
связана с хромосомным материалом ядра.
Кариоплазма(от
карио… и плазма), кариолимфа, ядерный
сок, содержимое клеточного ядра, в к-рое
погружены хроматин, ядрышки, а также
различные внутриядерные гранулы. После
экстракции хроматина химич. агентами
в К. сохраняется т. н. внутриядерный
матрикс, состоящий из белковых фибрилл
толщиной 2-3 нм, к-рые образуют в ядре
каркас, соединяющий ядрышки, хроматин,
поровые комплексы ядерной оболочки и
др. структуры.
№18
Способы
репродукции протоплазмы
Можно
выделить несколько источников и способов
обновления живой материи. Главным
является клеточный камбий, а в его
составе стволовые клетки. Большинство
стволовых клеток находится в клеточном
цикле. Каждая стволовая клетка способна
совершить от 50 до 65 циклов. Всякий раз
из нее образуются 2 клетки. Одна из них
вступает в фазу G0,
получает
программу развития, дифференцируется
и функционирует в составе органа, другая
сохраняя свойства стволовой клетки,
превращается в GI.
Другим
источником репродукции являются
дифференцированные клетки органов, не
имеющие стволовых. Здесь каждая делящаяся
клетка дает 2, получающие обе программы
развития. Образующиеся клетки называют
дочерними. Например, все дифференцированные
гепатоциты сохраняют способность к
пролиферации, благодаря чему печень
обладает хорошими регенеративными
способностями.
Клетки,
утратившие способность к делению,
обновляют живую материю с помощью
эндорепродукции, т.е их новая протоплазма
образуется в прежней форме. Это двуядерные
клетки, которые могут возникать с помощью
абортивного митоза, так и митоза.
Соматическая
протоплазма возникает наиболее часто
при атипичных формах митоза –
полицентрическом и моноцентрическом.
П. митозы характерны для мегакариоцитов
красного костного мозга. Единственным
полноценным способом воспроизводства
клеток является митоз.
№19
Жизненный
цикл клетки
Делящиеся
и неделящиеся клетки. Митоз. Дифференцировка
и специализация клеток. Этапы жизненного
цикла специализированной клетки. Некроз
и апоптоз. Регуляция численности клеток
в организме.
До
сих пор много тайн клетки остаются
неразгаданными. Загадочным во многом
остается и запрограммированный
генетически алгоритм ее жизни,
названный жизненным
циклом клетки (клеточным циклом).
Жизненный цикл клетки (рисунок
1)
начинается с момента ее образования
после деления родительской клетки и
заканчивается либо новым делением, либо
превращением в специализированную
клетку.

|
Рисунок |
Большинство
клеток продолжает делиться. Им свойственен
клеточный цикл, состоящий из периодически
повторяющихся стадий: так
называемой интерфазы (1) –
этапа подготовки к делению и непосредственно
процесса деления – митоза (2).
К этапам дифференцировки (3)
и функционирования специализированной
клетки (4) мы вернемся чуть позже.
На
стадии подготовки к делению происходит
удвоение генетического материала
(редупликация
ДНК).
Масса клетки во время интерфазы
увеличивается до тех пор, пока она
примерно вдвое не превысит начальную.
Отметим, что сам процесс деления намного
короче этапа подготовки к нему: митоз
занимает примерно 1/10 часть клеточного
цикла.
Цикличность
(периодическое повторение) стадий
интерфазы и митоза можно проиллюстрировать
на примере фибробластов –
одного из видов клеток соединительной
ткани (рисунок
2).
Так, нормальные фибробласты эмбриона
человека размножаются приблизительно
50 раз. Каков генетически запрограммированный
предел возможных делений клетки –
это одна из неразгаданных тайн биологии.
![]()
|
Рисунок |
Жизненный
цикл клеток базального слоя эпидермиса
в обычных условиях составляет 28-60 дней.
При повреждении кожи (конкретнее –
при повреждении мембран и разрушении
клеток эпидермиса под воздействием
внешних факторов) выделяются
особые биологически
активные вещества.
Они значительно ускоряют процессы
деления (это явление называется регенерацией),
именно поэтому ранки и ссадины так
быстро заживают. Максимальной
регенеративной способностью обладает
эпителий роговицы: одновременно в стадии
митоза находятся 5-6 тысяч клеток,
продолжительность жизни каждой из
которых 4-8 недель.
Хотя
все клетки появляются путем деления
предшествующей (материнской) клетки
(“Всякая клетка от клетки”), не все они
продолжают делиться. Клетки, достигшие
некоторой стадии развития при
дифференцировке, могут терять способность
к делению.
Дифференцировка –
возникновение различий в процессе
развития первоначально одинаковых
клеток, приводящее к их специализации.
Процесс дифференцировки заключается
в последовательном считывании и
использовании наследственной информации,
что обеспечивает синтез различных
белков (в первую очередь ферментов),
характерных для данного вида клеток.
Другими словами, различия между клетками
определяются набором белков, синтезируемых
в клетках определенного вида.
Различия
между клетками определяются набором
синтезируемых в них белков.
При
дифференцировке набор хромосом в клетке
не меняется, изменяется лишь соотношение
активных и неактивных генов, кодирующих
различные белки.
Существуют
два типа регуляции
экспрессии (активации
или блокирования) генов:
-
Кратковременная адаптивная
активация (реже блокирование), зависящая,
в частности, от концентрации вещества,
включающегося в обмен веществ (исходного
вещества или продукта метаболизма).
Этот механизм выработался эволюционно
как приспособительная реакция и особенно
ярко проявляется у животных (например,
быстрый синтез пигментов у хамелеона
в зависимости от условий). -
Длительное (в
течение всей жизни клетки и/или многих
генераций клеток!) блокирование или
активация гена, возникающее в ходе
клеточной дифференцировки. Например,
в ДНК любой клетки желудка есть ген,
отвечающий за синтез белков, из которых
состоит ноготь. Но он необратимо
блокирован гистонами и
другими белками (этот участок ДНК плотно
упакован), что никогда не позволит
считывать с него информацию. Поэтому
в желудке не растут ногти; а гены,
ответственные за синтез гемоглобина,
функционируют только у молодых форм
эритроцитов, но не действуют в зрелых
эритроцитах или других клетках.
На рисунке
1. цифрами
3 и 4 отмечены этапы дифференцировки и
активного функционирования
специализированной клетки.
Нервные
клетки мозга, однажды возникнув, уже не
делятся. В течение жизни число нейронов
постепенно уменьшается. Поврежденные
ткани мозга неспособны восстанавливаться
путем регенерации. Однако изначально
число нейронов в мозге настолько велико,
что до конца жизни человека они способны
поддерживать необходимые связи в нервной
системе.
В
качестве примера клеток, неспособных
к делению, можно рассмотреть эритроциты.
Как известно, эритроциты в процессе
специализации теряют ядро, следовательно,
не имеют в своем составе ДНК. Возникают
эритроциты из так называемой стволовой
клетки костного
мозга. Клеткой-предшественницей(стволовой
клеткой)
называют клетки кроветворной ткани,
которые на протяжении всей жизни человека
сохраняют способность делиться и, тем
самым, поставлять дочерние клетки,
которые в дальнейшем будут специализироваться
в одном направлении и замещать погибшие
клетки. Срок жизни и активного
функционирования эритроцитов невелик
(около 4 месяцев), затем они разрушаются,
в основном в селезенке.
Этапы
жизни специализированной клетки,
неспособной к делению (нейрона,
эритроцита), условно можно изобразить
на оси времени линией, разделенной на
несколько отрезков ( рисунок
3 ).
Эти отрезки дают представление о
временном соотношении периодов жизни
такой клетки: рождения, созревания и
активного функционирования, угасания
(старения) и естественной гибели.
![]()
|
Рисунок |
Время
протекания каждого этапа и продолжительность
жизненного цикла для однотипных клеток
в нормальных условиях практически
одинаковы.
Продолжительность
жизненного цикла однотипных клеток в
нормальных условиях практически
одинакова.
Например,
эритроциты живут 90-125 дней, а тромбоциты –
всего 4 суток. Это говорит о том, что
клетки используют для отсчета времени
своей жизни некий механизм, алгоритм,
заложенный в них природой. И в каждый
момент жизни клетка
строго следует законам, продиктованным
этим алгоритмом.
На
всех этапах клеточного цикла варьируют
значения некоторых параметров
жизнедеятельности клетки, и, в частности,
отмечается различная скорость и
интенсивность протекания
процессов метаболизма( рисунок
4 ).
Это обусловлено, в первую очередь,
непрерывно меняющейся активностью ферментов,
благодаря которым протекают все реакции
в клетке. Ферменты могут синтезироваться
в клетке “по мере надобности”,
активироваться, временно блокироваться
или полностью разрушаться (подробнее
о ферментах будет сказано позже).
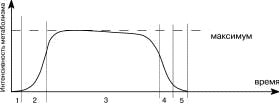
|
Рисунок |
Рассмотрим
подробнее наиболее характерные процессы,
происходящие на каждом из этапов
клеточного цикла.
Рождение.
Отправным моментом жизни любой клетки
(кроме половой, для которой характерен
мейоз) считают деление материнской
клетки с образованием двух идентичных
дочерних – митоз (от греческогоmitos –
нить). Во время митоза основная задача
материнской клетки – поровну передать
равноценный в количественном и
качественном отношении генетический
материал дочерним
клеткам.
Митоз
часто называют “танцем хромосом”.
Каждая следующая фигура в этом танце
не случайна, здесь нет ни одного лишнего
или бессмысленного “па” – это еще
один четкий, выверенный природой
алгоритм. В. Дудинцев в романе “Белые
одежды” так описывает процесс деления
клетки: “Хромосомы шевелились, как
клубок серых червей, потом вдруг
выстроились в строгий вертикальный
порядок. Вдруг удвоились – теперь
это были пары. Тут же какая-то сила
потащила эти пары врозь, хромосомы
подчинились, обмякли, и что-то повлекло
их к двум разным полюсам.”
Деление
клетки на две идентичные (митоз)
характеризуется сменой нескольких
морфологически и физиологически
различающихся стадий ( рисунок
5 ).
На первой стадии митоза хроматин плотно
упаковывается (этот процесс
называется суперспирализацией хроматина)
с образованием хромосом (1). Каждая
хромосома состоит из двух идентичных
половинок (хроматид) – будущих
дочерних хромосом. Затем при сокращении
так называемого веретена деления (2),
представляющего собой комплекс
микротрубочек и микрофибрилл, дочерние
хромосомы расходятся, буквально
подтягиваются нитями веретена деления
к противоположным полюсам клетки. После
окончательного расхождения дочерние
хромосомы вновь раскручиваются,
превращаясь в длинные и тонкие нити
хроматина (3). Веретено деления исчезает,
хроматин в дочерних клетках окружается
ядерной оболочкой, и между дочерними
клетками образуется поперечная перетяжка
(4) из клеточных мембран.
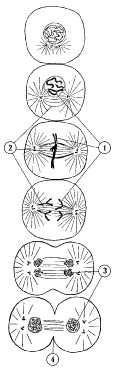
|
Рисунок |
Хромосомы,
как мы уже говорили, представляют собой
максимально плотно упакованные нити
ДНК, с которых на этапе деления невозможно
считывание информации. Соответственно,
на этапе деления не происходит биосинтеза
белка, интенсивность процессов метаболизма
минимальна, транспорт веществ в клетку
и из нее практически равен нулю. Все
процессы в делящейся клетке направлены
на выполнение главнейшей задачи –
максимально точно, без искажения,
передать генетическую информацию
дочерним клеткам, – в ущерб
второстепенным (на данном этапе!)
функциям.
Созревание.
В этот период происходит дифференцировка клеток
и становление ключевых ферментных
систем.
Клетка готовится выполнять предназначенные
природой функции, постепенно активизируя
свой обмен веществ.
Активное
функционирование.
Интенсивность реакций метаболизма и
сопряженного с нимэнергетического
обмена в
это время максимальны.
В
период активного функционирования
интенсивность обмена веществ в клетке
максимальна.
Процессы
в клетке направлены на обеспечение
постоянства внутренней среды и выполнение
специфических функций: нейрон воспринимает
и передает нервный импульс, эритроцит
переносит кислород и так далее.
Угасание
(старение).
Этот процесс запрограммирован генетически
и, в первую очередь, проявляется
уменьшением выработки и активности
ферментов в клетке. При этом замедляются
биохимические реакции, тормозится
метаболизм и энергетический обмен.
Период
старения клетки характеризуется
уменьшением выработки и активности
ферментов.
Стареющие
клетки, как правило, имеют неудвоенное
количество ДНК, но сохраняют жизнеспособность
и некоторую метаболическую активность
в течение определенного времени.
Естественная
гибель клетки (апоптоз).
К сожалению, до сих пор процесс естественной
гибели клеток до конца не изучен.
Известно,
что в клетке из-за блокирования ферментов
прекращается синтез белка, а нет белка –
нет и жизни. Морфологически апоптоз
характеризуется разрушением ядра и
цитоплазмы. “Осколки” погибшей клетки
поглощаются и перерабатываются
специальными клетками иммунной
системы – фагоцитами.
Но ведь клетки могут погибнуть и под
воздействием случайных факторов
(механических, химических и любых
других). Случайная гибель клеток (а также
ткани, органа) в биологии называется некрозом.
Важно то, что естественная клеточная
гибель (апоптоз) в отличие от некроза
не вызывает воспаления в окружающих
тканях.
Апоптоз
не вызывает воспаления в окружающих
тканях.
В
организме запрограммированная клеточная
гибель выполняет функцию, противоположную
митозу, и, тем самым, регулирует общее
число клеток в организме. Апоптоз играет
важную роль в защите организма при
вирусных инфекциях. В частности,
иммунодефицит при ВИЧ-инфекции
определяется нарушениями в контроле
апоптоза.
Теперь,
когда мы рассмотрели все этапы жизненного
цикла клеток, коротко остановимся на
процессах регуляции численности клеток
в организме. Во время эмбриогенеза (первого
этапа внутриутробного развития) число
клеток постоянно возрастает, причем в
геометрической прогрессии ( рисунок
6 ).
Зигота,
образовавшаяся после слияния яйцеклетки
и сперматозоида, делится с образованием
двух дочерних клеток. Затем, в результате
последовательных делений, образуются
четыре, восемь, шестнадцать клеток и
так далее. Параллельно с увеличением
численности на этапе эмбриогенеза
происходит дифференцировка клеток –
так образуются ткани (смотри
лекцию Ткани, их строение и функции).
Во
взрослом организме общая численность
клеток стабильна, она остается практически
неизменной на протяжении многих лет
( рис.
7).
Это
происходит за счет уравновешивания
процессов возникновения новых клеток
(митоза) и гибели клеток, естественной
(апоптоза) или случайной (некроза). При
смещении равновесия, например, гибели
большого количества клеток в результате
травмы или другого негативного
воздействия, включаются механизмы
регенерации (увеличение интенсивности
деления клеток для замещения погибших),
о которых уже было сказано. Таким образом,
общая численность клеток поддерживается
практически на постоянном уровне.
№
20
Клеточный
цикл и его фазы
Клеточный
цикл состоит из митоза и интерфазы.
Митоз
начинается с профазы. В профазе происходит
спирализация хромосом, образование
нитчатых структур. Хроматин конденсируется
в хромосомы и полностью исчезает.
Исчезает ядрышко. В конце профазы
разрушается кариолемма. В цитоплазме
распадается и исчезает ЭПС и к.Гольджи,
прекращается образование РНК и синтез
белка. Лизосомы и митохондрии сохраняются
и начинает формироваться аппарат
деления.
Прометафаза:
движение хромосом к полюсам клетки.
Метафаза:
Свободнолежащие хромосомы мигрируют
к экватору клетки и образуют фигуру,
которую называют экваториальной
пластинкой. Кинетохоры хромосом у
полюсов ядра связываются с микротрубочками.
Стадию метафазы называют одиночной
звездой – монастер или материнской
звездой. Каждая хромосома состоит из
сестринских гомологичных хроматид,
полностью разделенных, исключая район
кинетохора. Далее происходит разделение
кинетохора.
Анафаза:
Поделившиеся материнские хромосомы
начинают мигрировать к полюсам клетки.
По экватору клетки появляется борозда
деления, знаменующая начало цитотомии.
Телофаза:
Реконструкция ядра и образование новых
клеток. После стабилизации хромосом на
полюсах клетки начинается процесс
реконструкции ядерной оболочки.
Деконденсация хромосом, появляются
глыбки хроматина, формируется ядрышко.
Интерфаза
G1
: Образование РНК, синтез белка, рост
клетки. В этой фазе клетка задерживается
в точке рестрикции и может находиться
в ней долго. Для преодоления этой точки
нужен специальный белок.
S:
В эту фазу попадает диплоидная клетка,
имеющая хромосомы. Синтез ДНК. Клетка
становится тетраплоидной.
G2:
Образование АТФ. Синтез белков-тубулинов
для сборки микротрубочек и веретена
деления.
№21
Хромосомы
и их организация. Хромосомный набор
человека.
Хромосомный
набор или кариотип, в диплоидном
количестве содержат соматические клетки
и в гаплоидном – половые. В наборе
различают автосомы, ответственные за
все признаки организма, исключая
принадлежность к полу. Хромосомный
набор контролируют 3 закона: Закон
постоянства числа хромосом:
Диплоидный набор у человека состоит из
46 хромосом. Среди 46 – 23 наследуются по
материнской и 23 по отцовской линии и
являются гомологичными ( кроме х и у).
Закон
постоянства формы хромосом:
парные отцовские и материнские хромосомы
имеют одинаковую форму. Различают
метацентрические, субметацентрические,
акроцентрические. Некоторые хромосомы
имеют сателлит и поперечную исчерченность.
Закон
постоянства величины хромосом: Хромосомы
набора имеют разную величину. Самую
большую величину имеют хромосомы первой
пары, самую малую из аутосомного состава
– 22 пара. Наименьшие размеры имеет
у-хромосома.
Хромосомы
образованы из ДНК, гистонов и негистоновых
белков. Гистоны необходимы для укладки,
компактизации нитей ДНК в хромосомы.
Основу хромосомы составляет двойная
спираль ДНК. Первый уровень ее укладки
– нуклеосомный (отрезки ДНК связываются
в глобулы гистонов). Нуклеомерный —
формирование петель из молекул ДНК.
Хромонемный – образование нитей.
Хроматиды – нитчатые хромонемы
составляющие структуру хроматиды.
№22
Паранекроз,
дистрофия и смерть клетки. Апоптоз и
некроз.
Паранекроз
– совокупность неспецифических обратимых
изменений цитоплазмы, возникающих под
действием различных агентов. При
паранекрозе в цитоплазме происходит
разрушение белковых тел, ее обводнение
и увеличение сорбционных свойств. Важным
признаком паранекроза так же считают
изменение кислотно-щелочного равновесия.
Среда клетки меняется на кислую.
Поврежденные клетки резко снижают
митотическую активность, часто
задерживаются на разных стадиях митоза,
главным образом из-за нарушения
митотического аппарата, очень
чувствительного к изменениям внутренней
среды. Повреждение клеток внешними и
внутриорганизменными факторами может
привести к нарушениям регуляции их
метаболизма. При этом происходит
интенсивное отложение или наоборот,
резорбция ряда клеточных включений.
Кроме того наблюдается нарушение
регуляции проницаемости клеточных
мембран, что приводит к вакуолизации
мембранных органелл. Такие изменения
называются дистрофией. При необратимых
повреждениях клетки гибнут. В самом
начале изменения клеток имеют характер
обратимых , паранекротических. Отличие
состоит в том, что после снятия воздействия
они не исчезают, а прогрессируют. Явным
признаком клеточной гибели является
активация внутриклеточных гидролитических
ферментов. Они активируются в гиалоплазме
и начинают расщепление белков, липидов.
При этом разрешается внутриклеточная
мембрана. Это приводит к лизису.
Апопто́з (греч. απόπτωσις —
опадание листьев) — программируемая
клеточная смерть, регулируемый процесс
самоликвидации на клеточном уровне, в
результате которого клетка фрагментируется
на отдельные апоптотические тельца,
ограниченные плазматической мембраной.
Фрагменты погибшей клетки обычно очень
быстро (в среднем за 90
минут[1])фагоцитируются макрофагами либо
соседними клетками, минуя
развитие воспалительной
реакции.
Морфологически регистрируемый процесс
апоптоза продолжается 1—3 часа.[2]Одной
из основных функций апоптоза является
уничтожение дефектных (повреждённых,
мутантных, инфицированных) клеток. В
многоклеточных организмах апоптоз к
тому же задействован в процессах
дифференциации и морфогенеза,
в поддержании клеточногогомеостаза,
в обеспечении важных аспектов развития
и функционирования иммунной
системы.
Апоптоз реализует гены летальности,
функции которых активируют внеклеточные
факторы, среди которых преимущественное
значение имеют оксид азота и цитоксины.
Гены, вызывающие апоптоз и блокирующие
его, локализуются в ядре, на мембранах
ЭПС и митохондриях. Изменения начинаются
с ядра. Гетерохроматин собирается в
крупные электронно-плотные агрегаты ,
мигрирующие в цитоплазму. Поверхность
клетки вследствие гибели элементов
цитоскелета становится неровной, ЭПС
и митохондрии вакуолизируются и их
мембраны фрагментируются. Процесс
кончается образованием апоптозных тел,
которые поглощаются макрофагами.
Некро́з (от греч. νεκρός — мёртвый),
или омертве́ние —
это патологический процесс, выражающийся
в местной гибели ткани в живом организме
в результате какого-либо экзо- или
эндогенного её повреждения. Некроз
проявляется в
набухании, денатурации и коагуляции цитоплазматических
белков, разрушении клеточных органелл и,
наконец, всей клетки. Наиболее частыми
причинами некротического повреждения
ткани являются: прекращение кровоснабжения
(что может приводить к инфаркту, гангрене)
и воздействие патогенными продуктами
бактерий или вирусов (токсины,
белки, вызывающие реакции
гиперчувствительности,
и др.).
№23
Способы
и уровни адаптации клетки
Молекулярный
уровень – соматическая полиплоидизация
клетки. Субклеточный уровень – гиперплазия
цитоплазматических органелл. При
физических нагрузках в мышечных волокнах
увеличивается количество миофибрилл.
Клеточный уровень – осуществляется
путем увеличения числа клеток
(пролиферация), замена одних клеток на
другие (метаплазия).
№24
Информация
положения, детерминация, дифференцировка
и специализация клеток.
Внешняя
среда, устанавливающая число и качество
работающих генов, называется информацией
положения или позиционной формацией.
(сигналы, сообщающие клетке об ее
положении).
Позиционная
информация может быть: химической
(для
полной регенерации кости необходимо
поступление достаточного количества
кислорода). При неправильном наложении
гипсовой повязки развивается ложный
сустав. Гальванический
фактор:
обеспечивает направленность векторов
гальванических полей, по которым идет
рост конечностей. Контактная
информация
(результат взаимодействия м/д собой
отдельных клеток и клеточных групп.
Совокупность факторов, обеспечивающих
эмбриональную индукцию, которая
реализуется чаще всего через контактное
взаимодействие, например влияние хорды
на развитие НС.
При детерминации тканевых клеток происходит
стойкое закрепление их свойств,
вследствие чего ткани теряют способность
к взаимному превращению
(метаплазии).
В
биологиидетерминация —
это процесс определения дальнейшего
пути развития клеток.
Дифференцировка
клеток —
процесс реализации генетически
обусловленной программы формирования
специализированного фенотипаклеток,
отражающего их способность к тем или
иным профильным функциям. Иными словами,
фенотип клеток есть результат
координированнойэкспрессии (то
есть согласованной функциональной
активности) определённого набора генов.
В
процессе дифференцировки менее
специализированная клетка становится
более специализированной.
Например, моноцит развивается
вмакрофаг,
промиобласт развивается в миобласт,
который образуя синцитий,
формирует мышечное волокно. Деление,
дифференцировка иморфогенез—
основные процессы, путём которых
одиночная клетка (зигота)
развивается в многоклеточный организм,
содержащий самые разнообразные виды
клеток. Дифференцировка меняет функцию
клетки, её размер, форму и метаболическую
активность.
№25
Компетентные
и коммитированные клетки, конститутивные
и индуцибельные гены.
Клетки,
способные реагировать на информацию
положения, становятся компетентными.
Под ее влиянием геном разделяется на
гены экспрессированные и репрессированные.
Экспрессированные – рабочие,
репрессированные – нерабочие. Среди
экспрессированных генов имеются
конститутивные, однотипные для всех
клеток данного вида. Конститутивные
гены отвечают за общие признаки жизни
и организуют обмен веществ. Другие
экспрессрованные гены называются
индуцибельными. Они специфичны для
клеток, т.е. определяют характерные для
нее структурные и функциональные
признаки.
№26
Происхождение
тканей. Теории тканевой эволюции; роль
факторов внешней среды
Принято
считать, что в организме имеется 4
первичные функции и в отношении каждой
формируется присущий только ей тип
ткани (А.А. Заварзин)
Функции:
1
– Пограничная или разграничительная,
в направлении ее формируются ткани,
имеющие прямой контакт с внешней средой,
выполняют защиту от внешних воздействий,
получают из внешней среды трофический
материал и выводят из нее остаточные
продукты метаболизма. Это эпителиальные
ткани.
2
– Вторая функция организует внутреннюю
среду. В направлении этой функции
образуются ткани, формирующие мягкие
и твердые скелеты, основу интерстиция
органов, защитные и циркуляторные
системы организма. Эти ткани развиваются
из одного источника – мезенхимы и
поэтому называются мезенхимными или
тканями внутренней среды, соед. тканями.
3
– Третья функция обеспечивает подвижность,
является одним из главных признаков
жизни. В направлении ее формируются
мышечные ткани. Они делают возможным
перемещение тела в пространстве и
определяют моторику внутренних органов.
4
– Четвертая функция обогащает живые
системы способностью отвечать на
раздражители адекватной реакцией и
передавать возбуждение на исполнительные
органы. В направлении этой функции
развивается нервная ткань.
4
типа тканей, возникшие у первых
многоклеточных, в последствии у всех
представителей животного мира развивались
параллельно. Это значит, что у современного
животного и человека имеется 4 типа
тканей.
№27
Определение
и классификация тканей.
Ткань
– исторически сложившаяся система
клеточных и неклеточных элементов,
объединенных общностью происхождения,
строения и функций.
Первая
группа: ткани общего характера :
эпителиальные , мезенхимные. Вторая
группа (спец.ткани) мышечные и нервные.
Первая
группа: камбиальные ткани, имеющие
стволовые клетки. Эти ткани обладают
хорошей органотропической регенерацией.
Время их жизни ограничено днями (эпителий)
и несколькими годами (нерв.клетки).
Клетки гибнут путем апоптоза,
восстанавливаются с помощью физиологической
регенерации. Ткани одного характера
обладают широкой структурной и
функциональной лабильностью. Трансформация
одного вида тканей в другой возможна в
пределах типа. Эту перестройку называют
метаплазией. Метаплазия не имеет места
в специальных тканях, в отсутствие
стволовых клеток их клеточные элементы
не восстанавливаются. Все ткани имеют
дифферонную организацию. Дифферон –
группа клеток, однонаправлено развивающихся
из стволовой клетки. Диффероны характерны
для тканей общего характера.
|
Эпителиальная |
Соединительная |
Мышечная |
Нервная |
|
Много Покрывают Образует |
Мало Образует |
Возбуждается |
Возбуждается |
№28
Эпителий
– определение и общая характеристика,
функции.
Эпителиальные
ткани – группа пограничных клеток,
выполняющих в организме защитную,
всасывательную и выделительную функции.
Эпителиальные
ткани —
древнейшие гистологические структуры,
которые в фило- и онтогенезе возникают
первыми. Основное свойство эпителиев
— пограничность. Эпителиальные
располагаются на границах двух сред,
отделяя организм или органы от окружающей
среды.
Эпителии,
как правило, имеют вид клеточных пластов
и образуют наружный покров тела, выстилку
серозных оболочек, просветов органов,
сообщающихся с внешней средой во взрослом
состоянии или в эмбриогенезе. Через
эпителии осуществляется обмен веществ
между организмом и окружающей средой.
Важной функцией эпителиальных тканей
является защита подлежащих тканей
организма от механических, физических,
химических и других повреждающих
воздействий. Некоторые эпителии
специализированы на выработке
специфических веществ — регуляторов
деятельности других тканей
организма.
Производными
покровных эпителиев являются
железистые эпителии. Особый вид эпителия
— эпителий органов чувств.
Эпителии развиваются
с 3-4-й недели эмбриогенеза человека из
материала всех зародышевых листков.
Некоторые эпителии, например эпидермис,
формируются как полидифферонные ткани,
так как в их состав включаются клеточные
диффероны, развивающиеся из разных
эмбриональных источников (клетки
Лангерганса, меланоциты и др.).
Эпителий
не имеет кровеносных сосудов, питание
осущ. диффузно, через базальную мембрану.Э.
не имеет межклеточного вещества между
клетками. Клетки связаны с помощи
десмосом, плотных контактов и др.
№29
Классификация
эпителиальных тканей.
Добрый день! Сегодня я расскажу вам о том, как сдать экзамен по гистологии. Ну и, разумеется, как к нему готовиться.
Экзамен по гистологии имеет свою специфику. Она заключается в том, что, помимо теоретических вопросов (у нас их было 70), у вас будет также набор из препаратов (кажется, нас пугали 50-ю препаратами).
Особенности теории
1)Теоретическая часть гистологии во многом повторяет курс нормальной физиологии. Кроветворная система, тесно связанная с ней имунная система, эндокринная система – всё это физиология один в один. Здесь есть отличный бонус – если вы поймёте гистологию, вам будет очень, очень легко на физиологии. А «физо» — это один из важнейших фундаментальных предметов всей медицины;
Следствие этой особенности: гистологию нужно знать. Крайне важная и полезная вещь. Надеюсь, вы весь год учились более-менее нормально, и у вас в голове просто хаос, но не стерильность.
2)Теория гистологии изобилует классификациями. Здесь всё делится на всё и всё состоит из всего. Например, кровь. Она состоит из форменных элементов и плазмы. Форменные элементы делятся на лейкоциты, эритроциты и тромбоциты. Лейкоциты делятся на гранулоциты (с зёрнышками) и агранулоциты (без зёрнышек). Гранулоциты делятся на эозинофилы, базофилы и нейтрофилы. Ну и так далее.
Ваш ход: используйте черновики, пишите классификации, но не вздумайте делать это в строчку (как абзацем выше). Вы ничего не поймёте и ещё больше запутаетесь. Вот как нужно писать классификацию:
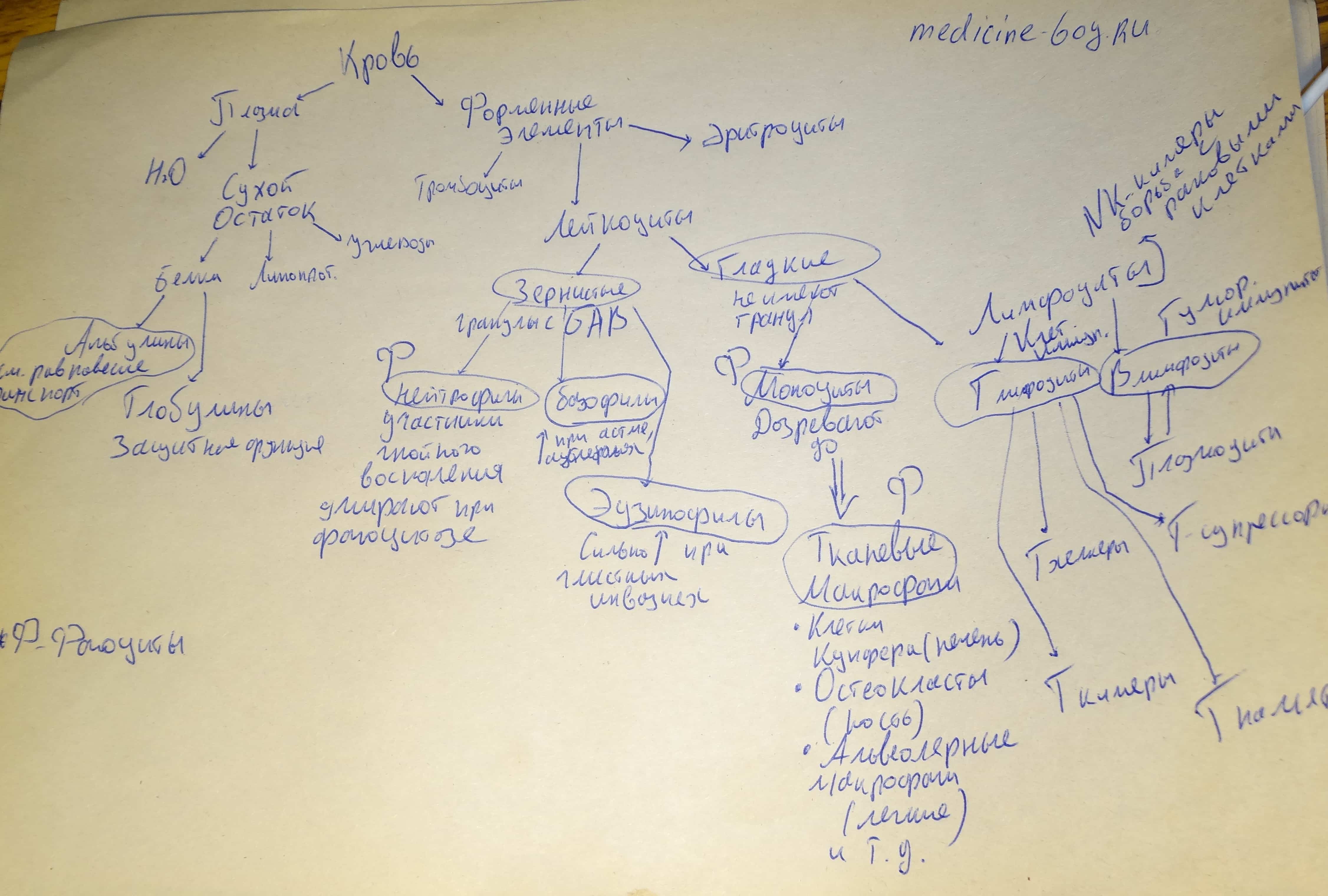
Как видите, она расположена вертикально, так вам сразу видна иерархия. И ещё — около каждого элемента подписываете его особые приметы, для лучшего запоминания. Идеально, если этой приметой будет функция.
3)Ещё одна особенность – здесь присутствует и собственно теория гистологии (как ни странно). То есть вам нужно будет знать особенности строения клеток и тканей в теории, ну то есть вы должны уметь их рассказать без препарата.
Знание строения бесполезно без знания функций. Например, если вы знаете функцию нейтрофилов (пожирание и уничтожение чужеродных агентов в нашем организме), то вы сразу же запомните, зачем нейтрофилам гранулки-зёрна (в них находятся разрушающие ферменты).
Как учить препараты по гистологии
1.Препараты следует учить одновременно с функцией ткани. Например, у вас препарат красного костного мозга – вы запоминаете особенности препарата (о них чуть ниже) и сразу же запоминаете то, как это всё работает. Какие там есть ростки, каких размеров клетки, что регулирует их дифференцировку, пролиферацию и т.д.
Однако, если у вас экзамен через неделю-две, а невыученного много, то можно учить отдельно препараты и отдельно теорию. Это чуть похуже, но вполне допускается.
2.Оставайтесь после пар и рассматривайте именно те препараты, которые вам будут давать на экзамене. Это чертовски важно. Или, на крайний случай, фотографируйте их и дома рассматривайте именно их фотографии. Не из учебника, не из атласа, и не из подборки «все препараты за 5 минут для чайников». Вам нужно знать именно ваши препараты, потому что малейшее изменение оттенка цвета препарата в атласе может вас здорово запутать.
Нет, не подумайте, я не против атласов. Ими круто пользоваться во время учебного года, готовясь к семинарам, зачётам и коллоквиумам. Но перед экзаменами перед вами должны быть только ваши препараты.
Объясню свою категоричность – в атласе может быть один кусок препарата, а у вас на стекле будет другой кусок того же препарата, в некоторых случаях (далеко не во всех) это может запутать. Ещё интересная штука – краситель. Один и тот же краситель может сильно отличаться по степени насыщенности. В вашем атласе фото с конго-ротом (отличный краситель для желудка, больше ничего им не красится) чутка выцветет и станет неотличим от гематоксилина-эозина (им красится 70% всех препаратов).
3.Соритруйте препараты. Обязательно. Не вздумайте учить целый массив препаратов просто смотря на них по нескольку минут. Выделяйте группы по 5-7 препаратов по системам (например, все препараты по теме «сердце и сосуды») или по тому, в каком порядке вы их сдавали в учебный год (например, препараты эпителиальных и соединительных тканей) и обязательно ищите особые приметы.
На самом деле, особые приметы препаратов – краеугольный камень всего курса гистологии. Набирайте группы препаратов и из них выделяйте самые запоминающиеся. Если вы весь год учили препараты, смело выделяйте несколько самых-самых необычных из всей массы. Например, отличный препарат – пластичная костная ткань. Он окрашен методом Шморля, то есть вы увидите изумрудно-зелёные пластины с вкраплениями оранжевого. Ни один препарат даже близко не похож на это. Красотища!

Ещё один препарат – человеческий волос. Это длинный, тёмный, толстый отросток под микроскопом, его вы также не перепутаете ни с чем. Правда, у нас в последний момент его исключили — видимо, он был слишком лёгкий и слишком ненужный. Посмотрите лучше на прекрасную нёбную миндалину.
Не забудьте, что вы можете запоминать и макропризнаки (то есть те, которые видны и без микроскопа). Например, в нашем наборе препаратов только один имел треугольную форму на стёклышке, это был тимус.
Если вы точно уверены, что работаете с экзаменационными препаратами, ориентироваться по макропризнакам вполне можно. Это такой легальный чит.

4.Ну и дополнение предыдущего признака. Как выглядит подготовка по препаратам чисто технически: пишете список из всех препаратов, которые у вас есть, затем, рядом с ним, пишете те препараты, которые вы уже на 100% отличаете от остальных.
Здесь и должны использоваться все признаки, о которых мы говорили ранее – необычные окраски, необычные формы. Например, наша преподавательница сравнивала препарат «волокна Пуркьнье стенки сердца» с «девичьими мечтами». Ну типа они такие же розовые и воздушные. Странное сравнение, но оно нам чертовски помогло запомнить. Я когда впервые услышал, поулыбался, но потом, запомнив эту ассоциацию, сразу же перенёс миокард в список препаратов, которые я уже не перепутаю ни с чем.
Итак, ваша задача – сделать так, чтобы столбик со списком препаратов равнялся столбику тех препаратов, которые вы сразу же узнаете. Это, конечно, весьма сложно, но процентов 70-80 вы должны знать.
Как сдавать экзамен:
И ещё парочку советов по тому, что делать на самом экзамене.
1)На теоретической части используйте ваши знания по анатомии. Если всё совсем плохо, и вы взяли очень, очень неудачный для вас билет (в идеале такого не должно быть), старайтесь тащить анатомией. Это по-разному применимо к разным системам. Например, мне с моим вопросом про строение бронхиального дерева (ужасный вопрос для любителя эндокринки и иммунки) это весьма помогло. С другими темами могло бы быть хуже, но, в любом случае, анатомия – это ваш костыль, на который вы должны опираться в случае проблем с теорией;
2)Когда вы возьмёте препарат и увидите что-то совсем незнакомое, рассуждайте. Первым делом вы обязательно должны уметь отличать паренхиматозные органы (почка, печень, железы) от трубчатых (мочеточник, желудок, пищевод) и от специфических (мозжечок, стенка сердца). Когда начнёте отвечать по препарату, первым делом называйте категорию вслух. Вы должны на 100% быть уверены, что перед вами что-то полое или паренхиматозное или мышечное или относящееся к иным органам.
Только важный момент: если вы увидели, что, например, перед вами пищевод, не говорите сразу – «пищевод». Сначала скажите, что видите полый орган. Затем, после пары секунд раздумий, сообщите преподавателю, что вероятнее всего, орган относится к пищеварительной системе. И только после этого должна пойти фраза «видимо, это пищевод».
Почему именно так? Во-первых, вы показываете преподавателю, что умеете рассуждать. Во-вторых, вы вызываете желание преподавателя помочь вам (не со всеми учителями работает, но со многими). Тот же пищевод можно очень легко спутать с мочеточником, в некоторых наборах препаратов эти полые органы не отличить. Преподаватель с большей охотой поможет, если вы ошибётесь на стадии отличия пищевода от мочеточника (этим нам поможет фраза «скорее всего, это пищеварительная система»). Если же вы не можете отличить полый орган от паренхимы, вам явно на пересдачу. Учитесь это делать в первую очередь.
В. Ю. Барсуков Гистология
1. Введение в курс гистологии. Органеллы клетки
Гистология – наука о строении, развитии и жизнедеятельности тканей живых организмов. Следовательно, гистология изучает один из уровней организации живой материи – тканевый.
Основным объектом гистологии в системе медицинского образования является организм здорового человека и потому данная учебная дисциплина именуется как гистология человека. Главной задачей гистологии как учебного предмета является изложение знаний о микроскопическом и ультрамикроскопическом строении клеток, тканей органов и систем здорового человека.
Задачей гистологии как науки является выяснение закономерностей строения различных тканей и органов для понимания протекающих в них физиологических процессов и возможности управления этими процессами.
Цитология – наука о строении, развитии и жизнедеятельности клеток. Следовательно, цитология изучает закономерности структурно-функциональной организации первого (клеточного) уровня организации живой материи. Клетка является наименьшей единицей живой материи, обладающей самостоятельной жизнедеятельностью и способностью к самовоспроизведению.
Клетка – это ограниченная активной мембраной, упорядоченная, структурированная система биополимеров, образующих ядро и цитоплазму, участвующих в единой совокупности метаболических и энергетиче-скихпроцессов.
Клетка – это живая система, состоящая из цитоплазмы и ядра и являющаяся основой строения, развития и жизнедеятельности всех животных организмов.
Основные компоненты клетки:
1) ядро;
2) цитоплазма.
По соотношению ядра и цитоплазмы (ядерно-цито-плазматическому отношению) клетки подразделяются на:
1) клетки ядерного типа (объем ядра преобладает над объемом цитоплазмы);
2) клетки цитоплазматического типа (цитоплазма преобладает над ядром).
По форме клетки бывают: круглыми (клетки крови), плоскими, кубическими или призматическими (клетки разных эпителиев), веретенообразными (гладкомы-шечные клетки), отростчатыми (нервные клетки) и др. Большинство клеток содержит одно ядро, однако в одной клетке может быть два, три и более ядер (многоядерные клетки). В организме имеются структуры (симпласты, синцитий), содержащие несколько десятков или даже сотен ядер. Морфология этих структур будет рассмотрена при изучении тканей.
Структурные компоненты цитоплазмы животной клетки:
1) плазмолемма (цитолемма);
2) гиалоплазма;
3) органеллы;
4) включения.
Плазмолемма – оболочка животной клетки, отграничивающая ее внутреннюю среду и обеспечивающая взаимодействие клетки с внеклеточной средой.
Органеллы – постоянные структурные элементы цитоплазмы клетки, имеющие специфическое строение и выполняющие определенные функции.
Митохондрии – наиболее обособленные структурные элементы цитоплазмы клетки, отличающиеся в значительной степени самостоятельной жизнедеятельностью.
2. Морфология и функции цитоплазмы клетки
Функции плазмолеммы:
1) разграничительная (барьерная);
2) рецепторная;
3) антигенная;
4) транспортная;
5) образование межклеточных контактов. Химический состав веществ плазмолеммы: белки, липиды, углеводы. В каждой липидной молекуле различают две части:
1) гидрофильную головку;
2) гидрофобные хвосты.
Гидрофобные хвосты липидных молекул связываются друг с другом и образуют билипидный слой. Гидрофильные головки соприкасаются с внешней и внутренней стороны. По выполняемой функции белки плазмолеммы подразделяются на:
1) структурные;
2) транспортные;
3) белки-рецепторы;
4) белки-ферменты;
5) антигенные детерминанты.
Различают следующие способы транспорта веществ:
1) способ диффузии веществ (ионов, некоторых низкомолекулярных веществ) через плазмолемму без затраты энергии;
2) активный транспорт веществ (аминокислот, нуклеотидов и др.) с помощью белков-переносчиков с затратой энергии;
3) везикулярный транспорт (производится посредством везикул (пузырьков)), подразделяется на эндоцитоз – транспорт веществ в клетку, экзоци-тоз – транспорт веществ из клетки. В свою очередь, эндоцитоз подразделяется на:
1) фагоцитоз – захват и перемещение в клетку;
2) пиноцитоз – перенос воды и небольших молекул.
В тех тканях, в которых клетки или их отростки плотно прилежат друг к другу (эпителиальная, гладкомышечная и др.), между плазмолеммами контактирующих клеток формируются связи – межклеточные контакты.
Типы межклеточных контактов:
1) простой контакт – 15–20 нм (связь осуществляется за счет соприкосновения макромолекул гликокаликсов);
2) десмосомный контакт – 0,5 мкм (с помощью скопления электроплотного материала в межмембранном пространстве);
3) плотный контакт (в этих участках межмембранные пространства отсутствуют, а билипидные слои соседних плазмолемм сливаются в одну общую били-пидную мембрану);
4) щелевидный, или нексусы, – 0,5–3 мкм (обе мембраны пронизаны в поперечном направлении белковыми молекулами, или коннексонами, содержащими гидрофильные каналы, через которые осуществляется обмен ионами и микромолекулами соседних клеток, чем и обеспечивается их функциональная связь);
5) синаптический контакт, или синапс, – специфические контакты между нервными клетками.
3. Морфология и функция органелы клетки
Классификация органелл:
1) общие органеллы, присущие всем клеткам и обеспечивающие различные стороны жизнедеятельности клетки;
2) специальные органеллы, имеющиеся в цитоплазме только определенных клеток и выполняющие специфические функции этих клеток.
В свою очередь, общие органеллы подразделяются на мембранные и немембранные. К мембранным органеллам относятся:
1) митохондрии;
2) эндоплазматическая сеть;
3) пластинчатый комплекс;
4) лизосомы;
5) пероксисомы.
К немембранным органеллам относятся:
1) рибосомы;
2) клеточный центр;
3) микротрубочки;
4) микрофибриллы;
5) микрофиламенты.
Форма митохондрий может быть овальной, округлой, вытянутой и даже разветвленной, но преобладает овально-вытянутая. Стенка митохондрии образована двумя билипидными мембранами, разделенными пространством в 10–20 нм. При этом внешняя мембрана охватывает по периферии всю митохондрию в виде мешка и отграничивает ее от гиалоплазмы. Функция митохондрий – образование энергии в виде АТФ.
Эндоплазматическая сеть (ЭПС) в разных клетках может быть представлена в форме уплощенных цистерн, канальцев или отдельных везикул. Стенка состоит из билипидной мембраны.
На наружной поверхности мембран зернистой ЭПС содержатся прикрепленные рибосомы.
Функции зернистой ЭПС:
1) синтез белков, предназначенных для выведения из клетки (на экспорт);
2) отделение (сегрегация) синтезированного продукта от гиалоплазмы;
3) конденсация и модификация синтезированного белка;
4) транспорт синтезированных продуктов в цистерны пластинчатого комплекса.
Пластинчатый комплекс Гольджи (сетчатый аппарат) представлен скоплением уплощенных цистерн и небольших везикул, ограниченных билипидной мембраной. Функции пластинчатого комплекса:
1) выведение из клетки синтезированных в ней продуктов (транспортная функция);
2) конденсация и модификация веществ, синтезированных в зернистой ЭПС;
3) образование лизосом (совместно с зернистой ЭПС);
4) участие в обмене углеводов;
5) синтез молекул, образующих гликокаликс цито-леммы;
6) синтез, накопление, выведение муцинов (слизи).
4. Морфология и функции органеллы клетки (продолжение)
Лизосомы – наиболее мелкие органеллы цитоплазмы, представляют собой тельца, ограниченные билипидной мембраной.
Функция лизосом – обеспечение внутриклеточного пищеварения, т. е. расщепление как экзогенных, так и эндогенных биополимерных веществ.
Классификация лизосом:
1) первичные лизосомы – электронно-плотные тельца;
2) вторичные лизосомы – фаголизосомы, в том числе аутофаголизосомы;
3) третичные лизосомы, или остаточные тельца. Пероксисомы – микротельца цитоплазмы (0,1—
1,5 мкм), сходные по строению с лизосомами, отличающиеся от них тем, что в их матриксе содержатся кристаллоподобные структуры, а в число белков-ферментов входит каталаза, разрушающая перекись водорода, образующуюся при окислении аминокислот.
Рибосомы – аппараты синтеза белка и полипептидных молекул.
По локализации подразделяются на:
1) свободные – находятся в гиалоплазме;
2) несвободные, или прикрепленные, – связаны с мембранами ЭПС.
Каждая рибосома состоит из малой и большой субъединиц. Каждая субъединица рибосомы состоит из рибосомальной РНК и белка – рибонуклеопротеида. Образуются субъединицы в ядрышке, а сборка в единую рибосому осуществляется в цитоплазме. Для синтеза белка отдельные рибосомы с помощью матричной (информационной) РНК объединяются в цепочки рибосом – полисомы. Клеточный центр (или цитоцентр, центросома) в неделящейся клетке состоит из двух основных структурных компонентов:
1) диплосомы;
2) центросферы.
Диплосома состоит из двух центриолей (материнской и дочерней), расположенных под прямым углом друг к другу. Каждая центриоль состоит из микротрубочек.
Функции цитоцентра:
1) образование веретена деления в профазе митоза;
2) участие в формировании микротрубочек клеточного каркаса;
3) в реснитчатых эпителиальных клетках центриоли являются базисными тельцами ресничек.
Микротрубочки – полые цилиндры (внешний диаметр – 24 нм, внутренний – 15 нм) являются самостоятельными органеллами, образуя цитоскелет.
Микрофибриллы (промежуточные филаменты) представляют собой тонкие неветвящиеся нити.
Микрофиламенты – еще более тонкие нитчатые структуры (5–7 нм), состоящие из сократительных белков (актина, миозина, тропомиозина). В совокупности микрофиламенты составляют сократительный аппарат клетки, обеспечивающий различные виды движений: перемещение органелл, ток гиалоплазмы, изменение клеточной поверхности, образование псевдоподии и перемещение клетки.
5. Морфология и функции ядра. Репродукция клеток
1. Структурные элементы ядра Структурные элементы ядра, перечисленные ниже, бывают хорошо выражены только в интерфазе:
1) хроматин;
2) ядрышко;
3) кариоплазма;
4) кариолемма.
Хроматин это вещество, хорошо воспринимающее краситель состоит из хроматиновых фибрилл, толщиной 20–25 нм, которые могут располагаться в ядре рыхло или компактно. При подготовке клетки к делению в ядре происходят спирализация хроматиновых фибрилл и превращение хроматина в хромосомы. После деления в ядрах дочерних клеток происходит деспира-лизация хроматиновых фибрилл
По химическому строению хроматин состоит из:
1) дезоксирибонуклеиновой кислоты (ДНК);
2) белков;
3) рибонуклеиновой кислоты (РНК).
Ядрышко – сферическое образование (1–5 мкм в диаметре), хорошо воспринимающее основные красители и располагающееся среди хроматина. Ядрышко не является самостоятельной структурой. Оно формируется только в интерфазе. В одном ядре содержится несколько ядрышек.
Микроскопически в ядрышке различают:
1) фибриллярный компонент (локализуется в центральной части ядрышка и представляет собой нити рибонуклеопротеида);
2) гранулярный компонент (локализуется в периферической части ядрышка и представляет собой скопление субъединиц рибосом).
Кариолемма – ядерная оболочка, которая отделяет содержимое ядра от цитоплазмы обеспечивает регулируемый обмен веществ между ядром и цитоплазмой. Ядерная оболочка принимает участие в фиксации хроматина. Функции ядер соматических клеток:
1) хранение генетической информации, закодированной в молекулах ДНК;
2) репарация (восстановление) молекул ДНК после их повреждения с помощью специальных репаратив-ных ферментов;
3) редупликация (удвоение) ДНК в синтетическом периоде интерфазы;
4) передача генетической информации дочерним клеткам во время митоза;
5) реализация генетической информации, закодированной в ДНК, для синтеза белка и небелковых молекул: образование аппарата белкового синтеза (информационной, рибосомальной и транспортных РНК).
Функции ядер половых клеток:
1) хранение генетической информации;
2) передача генетической информации при слиянии женских и мужских половых клеток.
В организме млекопитающих и человека различают следующие типы клеток:
1) часто делящиеся клетки клетки эпителия кишечника;
2) редко делящиеся клетки (клетки печени);
3) неделящиеся клетки (нервные клетки). Жизненный цикл у этих клеточных типов различен. Клеточный цикл подразделяется на два основных
периода:
1) митоз, или период деления;
2) интерфазу – промежуток жизни клетки между двумя делениями.
6. Общая эмбриология
Эмбриология – наука о закономерностях развития животных организмов от момента оплодотворения до рождения (или вылупливания на яйца). Следовательно, эмбриология изучает внутриутробный период развития организма, т. е. часть онтогенеза.
1. Онтогенез – развитие организма от оплодотворения до смерти.
2. Периоды эмбриогенеза
Эмбриогенез по характеру процессов, происходящих в зародыше, подразделяется на три периода:
1) период дробления;
2) период гаструляции;
3) период гистогенеза (образования тканей), органогенеза (образования органов), системогенеза (образования функциональных систем организма).
Дробление. Продолжительность жизни нового организма в виде одной клетки (зиготы) составляет у разных животных от нескольких минут до нескольких часов и даже дней, а затем начинается дробление. Дробление – процесс митотического деления зиготы на дочерние клетки (бластомеры). Дробление отличается от митотического деления тем что:
1) бластомеры не достигают исходных размеров зиготы;
2) бластомеры не расходятся, хотя и представляют собой самостоятельные клетки.
Различают следующие типы дробления:
1) полное, неполное;
2) равномерное, неравномерное;
3) синхронное, асинхронное.
Яйцеклетки и зиготы, содержащие умеренное количество желтка, также дробятся полностью, но образующиеся бластомеры имеют разную величину и дробятся неодновременно – дробление полное, неравномерное, асинхронное.
В результате дробления образуется вначале скопление бластомеров, и зародыш в таком виде носит название морулы. Затем между бластомерами накапливается жидкость, которая отодвигает бластомеры на периферию, а в центре образуется полость, заполненная жидкостью. В этой стадии развития зародыш носит название бластулы.
Бластула состоит из:
1) бластодермы – оболочки из бластомеров;
2) бластоцели – полости, заполненной жидкостью. Бластула человека – бластоциста. После образования бластулы начинается второй этап эмбриогенеза – гаструляция.
Гаструляция – процесс образования зародышевых листков, образующихся посредством размножения и перемещения клеток. В результате гаструляции в зародыше любого вида животного образуются три зародышевых листка:
1) эктодерма (наружный зародышевый листок);
2) энтодерма (внутренний зародышевый листок);
3) мезодерма (средний зародышевый листок). Гисто– и органогенез, процесс превращения зачатков
тканей в ткани и органы, а затем и формирование функциональных систем организма.
7. Эмбриология человека
Рассмотрение закономерностей эмбриогенеза начинается с прогенеза. Прогенез – гаметогенез (спер-мато– и овогенез) и оплодотворение.
Сперматогенез осуществляется в извитых канальцах семенников и подразделяется на 4 периода:
1) I период – размножения;
2) II период – роста;
3) III период – созревания;
4) IV период – формирования.
Сперматозоид человека состоит из двух основных частей: головки и хвоста. Головка содержит:
1) ядро (с гаплоидным набором хромосом);
2) чехлик;
3) акросому;
4) тонкий слой цитоплазмы, окруженный цитолеммой. Хвост сперматозоида подразделяется на:
1) связующий отдел;
2) промежуточный отдел;
3) главный отдел;
4) терминальный отдел.
Овогенез осуществляется в яичниках и подразделяется на 3 периода:
1) период размножения (в эмбриогенезе и в течение 1-го года постэмбрионального развития);
2) период роста (малого и большого);
3) период созревания.
Оплодотворение у человека внутреннее – в дистальной части маточной трубы. Подразделяется на три фазы:
1) дистантное взаимодействие;
2) контактное взаимодействие;
3) проникновение и слияние пронуклеусов (фаза синкариона).
В основе дистантного взаимодействия лежат три механизма:
1) реотаксис – движение сперматозоидов против тока жидкости о матке в маточной трубе;
2) хемотаксис – направленное движение сперматозоидов к яйцеклетке, которая выделяет специфические вещества – гиногамоны;
3) канацитация – активация сперматозоидов гинога-монами и гормоном прогестероном.
Через 1,5–2 ч сперматозоиды достигают дисталь-ной части маточной трубы и вступают в контактное взаимодействие с яйцеклеткой.
Ферменты, выделяемые сперматозоидом, разрушают лучистый венец и блестящую оболочку яйцеклетки. После этого начинается фаза проникновения. В области бугорка плазмолеммы яйцеклетки и сперматозоида сливаются, и часть сперматозоида (головка, связующий и промежуточный отделы) оказывается в цитоплазме яйцеклетки.
Условия, необходимые для оплодотворения яйцеклетки:
1) содержание в эякуляте не менее 150 млн сперматозоидов, при концентрации в 1 мл не менее 60 млн;
2) проходимость женских половых путей;
3) нормальное анатомическое положение матки;
4) нормальная температура тела;
5) щелочная среда в половых путях женщины.
С момента слияния пронуклеусов образуется зигота – новый одноклеточный организм. Время существования организма зиготы – 24–30 ч.
8. Эмбриология человека
Эмбриогенез
Эмбриогенез человека подразделяется на:
1) период дробления;
2) период гаструляции;
3) период гисто– и органогенеза. I. Период дробления. Дробление у человека полное
неравномерное, асинхронное. Бластомеры неравной величины, подразделяются на два типа: темные крупные и светлые мелкие. Крупные бластомеры дробятся реже, располагаются в центре и составляют эмбрио-бласт. Мелкие бластомеры чаще дробятся, располагаются по периферии от эмбриобласта и в дальнейшем формируют трофобласт.
На 5-е сутки бластоциста попадает в полость матки и находится в ней в свободном состоянии, а с 7-х суток происходит имплантация бластоцисты в слизистую оболочку матки (эндометрий). В процессе развития хориона выделяют два периода:
1) формирование гладкого хориона;
2) формирование ворсинчатого хориона. Из ворсинчатого хориона в последующем формируется плацента.
Функции хориона:
1) защитная;
2) как составная часть плаценты хорион принимает участие в выполнении многих ее функций: трофической, газообменной, экскреторной и др.
Стенка амниона состоит из:
1) внезародышевой эктодермы;
2) внезародышевой мезенхимы (висцерального листка).
Функции амниона: образование околоплодных вод и защитная функция. Стенка желточного мешка состоит из:
1) внезародышевой (желточной энтодермы);
2) внезародышевой мезенхимы. Функции желточного мешка:
1) кроветворная (образование стволовых клеток крови);
2) образование половых стволовых клеток (гонобла-стов);
3) трофическая функция (у птиц и рыб). Стенка аллантоиса состоит из:
1) зародышевой энтодермы;
2) внезародышевой мезенхимы.
Наружный зародышевый листок дифференцируется:
1) нейроэктодерму (из нее развиваются нервная трубка и ганглиозная пластинка);
2) кожная эктодерма (развивается эпидермис кожи);
3) переходная пластика (развивается эпителий пищевода, трахеи, бронхов);
4) плакоды (слуховая, хрусталиковая и др.). Внутренний зародышевый листок подразделяется на:
1) кишечную, или зародышевую, энтодерму;
2) внезародышевую, или желточную, энтодерму. Из кишечной энтодермы развиваются:
1) эпителий и железы желудка и кишечника;
2) печень;
3) поджелудочная железа.
9. Общие принципы организации тканей
Ткань – это система клеток и неклеточных структур, обладающая общностью строения, а иногда и происхождения, и специализированная на выполнении определенных функций. 1. Характеристика структурных компонентов ткани Клетки – основные, функционально ведущие компоненты тканей. Все ткани состоят из нескольких типов клеток.
Клеточная популяция – это совокупность клеток данного типа.
Клеточный дифферон, или гистогенетический ряд, – это совокупность клеток данного типа (данной популяция), находящихся на различных этапах дифферен-цировки.
Производные клеток:
1) симпласты (слияние отдельных клеток, например мышечное волокно);
2) синцитий (несколько клеток, соединенных между собой отростками, например сперматогенный эпителий извитых канальцев семенника);
3) постклеточные образования (эритроциты, тромбоциты).
Межклеточное вещество – также продукт деятельности определенных клеток. Межклеточное вещество состоит из:
1) аморфного вещества;
2) волокон (коллагеновых, ретикулярных, эластических).
Межклеточное вещество неодинаково выражено в разных тканях.
Классификации тканей:
1) эпителиальные ткани;
2) соединительные ткани (ткани внутренней среды, опорно-трофические ткани);
3) мышечные ткани;
4) нервная ткань.
Тканевой гомеостаз, или поддержание структурного постоянства тканей Регенерация тканей Формы регенерации:
1) физиологическая регенерация – восстановление клеток ткани после их естественной гибели (например, кроветворение);
2) репаративная регенерация – восстановление тканей и органов после их повреждения (травм, воспа-лений, хирургических воздействий и т. д.).
Интеграция тканей
Ткани входят в состав структур более высокого уровня организации живой материи: структурно-функциональных единиц органов и в состав органов, в которых происходит интеграция (объединение) нескольких тканей.
Механизмы интеграции:
1) межтканевые (обычно индуктивные) взаимодействия;
2) эндокринные влияния;
3) нервные влияния.
Например, в состав сердца входят: сердечная мышечная ткань, соединительная ткань, эпителиальная ткань.
10. Эпителиальные ткани
Характеристика эпителиальных тканей
Они образуют внешние и внутренние покровы организма. Функции эпителиев:
1) защитная (барьерная);
2) секреторная;
3) экскреторная;
4) всасывательная. Структурно-функциональные особенности эпителиальных тканей:
1) расположение клеток пластами;
2) расположение клеток на базальной мембране;
3) преобладание клеток над межклеточным веществом;
4) полярная дифференцированность клеток (на ба-зальный и апикальный полюсы);
5) отсутствие кровеносных и лимфатических сосудов;
6) высокая способность клеток к регенерации. Структурные компоненты эпителиальной ткани:
1) эпителиальные клетки (эпителиоциты);
2) базальная мембрана. Эпителиоциты являются основными структурными
элементами эпителиальных тканей. Базальная мембрана (толщина около 1 мкм) состоит из:
1) тонких коллагеновых фибрилл (из белка коллагена 4-го типа);
2) аморфного вещества (матрикса), состоящего из углеводно-белково-липидного комплекса.
Функции базальной мембраны: 1) барьерная (отделение эпителия от соединительной ткани);
2) трофическая (диффузия питательных веществ и продуктов метаболизма из подлежащей соединительной ткани и обратно); 3) организующая (прикрепление эпителиоцитов с помощью полудесмосом). Классификация желез По количеству клеток:
1) одноклеточные (бокаловидная железа);
2) многоклеточные (подавляющее большинство желез). По расположению клеток в эпителиальном пласте:
1) эндоэпителиальные (бокаловидная железа);
2) экзоэпителиальные.
По способу выведения секрета из железы и по строению:
1) экзокринные железы (имеют выводной проток);
2) эндокринные железы (не имеют выводных протоков и выделяют секреты (гормоны) в кровь или лимфу).
По способу выделения секрета из железистой клетки:
1) мерокриновые;
2) апокриновые;
3) голокриновые.
По составу выделяемого секрета:
1) белковые (серозные);
2) слизистые;
3) смешанные (белково-слизистые);
4) сальные.
Фазы секреторного цикла железистых клеток Существуют следующие фазы секреторного цикла железистых клеток:
1) поглощение исходных продуктов секретообразования;
2) синтез и накопление секрета;
3) выделение секрета (по мерокриновому или апокриновому типу);
4) восстановление железистой клетки.
11. Соединительные ткани
Собственно соединительные ткани
Развиваются структурно-функциональные особенности соединительных тканей:
1) внутреннее расположение в организме;
2) преобладание межклеточного вещества над клетками;
3) многообразие клеточных форм;
4) общий источник происхождения – мезенхима.
1. Морфологическая и функциональная характеристика рыхлой волокнистой неоформленной соединительной ткани
Морфологические особенности, отличающие рыхлую волокнистую соединительную ткань от других разновидностей соединительных тканей:
1) многообразие клеточных форм (9 клеточных типов);
2) преобладание в межклеточном веществе аморфного вещества над волокнами.
Функции рыхлой волокнистой соединительной ткани:
1) трофическая;
2) опорная – образует строму паренхиматозных органов;
3) защитная – неспецифическая и специфическая (участие в иммунных реакциях) защита;
4) депо воды, липидов, витаминов, гормонов;
5) репаративная.
Структурная и функциональная характеристика клеточных типов рыхлой волокнистой соединительной ткани.
Фибробласты – преобладающая популяция клеток рыхлой волокнистой соединительной ткани.
Они неоднородны по степени зрелости и функциональной специфичности и потому подразделяются на следующие субпопуляции:
1) малодифференцированные клетки;
2) дифференцированные или зрелые клетки, или собственно фибробласты;
3) старые фибробласты (дефинитивные) – фиброциты, а также специализированные формы фибробластов;
4) миофибробласты;
5) фиброкласты.
Макрофаги – клетки, осуществляющие защитную функцию,
Тканевые базофилы (тучные клетки) являются истинными клетки рыхлой волокнистой соединительной ткани.
Плазматические клетки (плазмоциты) – клетки иммунной системы. Образуются из В-лимфоцитов при воздействии на них антигенных веществ. Функция: синтез иммуноглобулинов.
Жировые клетки (адипоциты) содержатся в рыхлой соединительной ткани в неодинаковых количествах в разных участках тела и в разных органах.
Функции жировых клеток:
1) депо энергетических ресурсов;
2) депо воды;
3) депо жирорастворимых витаминов и др. Пигментные клетки (пигментоциты, меланоциты) —
клетки отростчатой формы, содержащие в цитоплазме пигментные включения (меланин).
12. Соединительные ткани
Соединительные ткани состоит из двух структурных компонентов:
1) из основного, или аморфного, вещества;
2) из волокон.
Основное, или аморфное, вещество состоит из белков и углеводов. Белки представлены коллагеном, альбуминами и глобулинами.
Углеводы представлены полимерными формами, в основном гликозаминогликанами.
Углеводные компоненты удерживают воду, в зависимости от содержания воды ткань может быть более или менее плотной.
Аморфное вещество обеспечивает транспорт веществ из крови клеткам и обратно.
Волокнистый компонент представлен коллагеновы-ми, эластическими и ретикулярными волокнами. В различных органах соотношение названных волокон неодинаково.
Эластические волокна характеризуются высокой эластичностью, способностью растягиваться и сокращаться, но незначительной плотностью.
Ретикулярные волокно по своему составу близко к коллагеновым волокнам, но в них более выражен углеводный компонент.
Отличается от рыхлой преобладанием в межклеточном веществе волокнистого компонента над аморфным.
Плотная оформленная соединительная ткань представлена в организме в виде сухожилий, связок, фиброзных мембран. Плотная волокнистая неоформленная соединительная ткань образует сетчатый слой дермы кожи.
Соединительные ткани со специальными свойствами
Ретикулярная ткань состоит из ретикулярных клеток и ретикулярных волокон. Эта ткань образует строму всех кроветворных органов (за исключением тимуса) и, обеспечивает трофику гемопоэтических клеток, влияет на направление их дифференцировки.
Жировая ткань состоит из скопления жировых клеток и подразделяется на две разновидности: белую и бурую жировую ткань.
Белая жировая ткань распространена в различных частях тела и во внутренних органах, неодинаково выражена у разных субъектов и на протяжении онтогенеза. Она представляет собой скопление типичных жировых клеток (адипоцитов).
Функции белой жировой ткани:
1) депо энергии;
2) депо воды;
3) депо жирорастворимых витаминов;
4) механическая защита некоторых органов. Локализуется она только в определенных местах: за
грудиной, около лопаток, на шее, вдоль позвоночника. Бурая жировая ткань состоит из скопления бурых жировых клеток, которые существенно отличаются от типичных адипоцитов и по морфологии, и по характеру обмена веществ в них. Слизистая соединительная ткань встречается только в эмбриональном периоде в провизорных органах, и прежде всего – в составе пупочного канатика. Она состоит в основном из межклеточного вещества, в котором локализуются фибро-бластоподобные клетки, синтезирующие муцин (слизь).
13. Скелетные соединительные ткани. Хрящевые ткани
Хрящевая ткань состоит из клеток – хондроцитов и хондробластов, а также из плотного межклеточного вещества.
Хондробласты располагаются одиночно по периферии хрящевой ткани. Представляют собой вытянутые уплощенные клетки с базофильной цитоплазмой, содержащей хорошо развитую зернистую ЭПС и пластинчатый комплекс. Эти клетки синтезируют компоненты межклеточного вещества, выделяют их в межклеточную среду, постепенно дифференцируются в дефинитивные клетки хрящевой ткани – хондроциты. Аморфное вещество содержит значительное количество минеральных веществ, не образующих кристаллы, воду, плотную волокнистую ткань. Сосуды в хрящевой ткани в норме отсутствуют. В зависимости от строения межклеточного вещества хрящевые ткани подразделяются на гиалиновую, эластическую и волок-ни-стую хрящевую ткань.
Эластическая хрящевая ткань характеризуется на-хождениемв клеточном веществе как коллагеновых, так и эластических волокон.
Волокнистая хрящевая ткань характеризуется содержанием в межклеточном веществе мощных пучков из параллельно расположенных коллагеновых волокон. Хондроциты располагаются между пучками волокон в виде цепочек. По физическим свойствам ткань характеризуется высокой прочностью. В организме встречается в ограниченных местах: составляет часть межпозвоночных дисков (фиброзное кольцо), локализуется в местах прикрепления связок и сухожилий к гиалиновым хрящам. В этих случаях прослеживается постепенный переход фиброцитов соединительной ткани в хондроциты хрящевой ткани. Хрящевая ткань – разновидность соединительной ткани, Хрящ – анатомический орган, который состоит из хрящевой ткани и надхрящницы. Надхрящница покрывает хрящевую ткань снаружи и состоит из волокнистой соединительной ткани. В надхрящнице выделяют два слоя:
1) наружный – фиброзный;
2) внутренний – клеточный, или камбиальный (ростковый).
Во внутреннем слое локализуются малодифферен-цированные клетки – прехондробласты и неактивные хондробласты, которые в процессе эмбрионального и регенерационного гистогенеза превращаются вначале в хондробласты, а затем в хондроциты.
В фиброзном слое располагается сеть кровеносных сосудов. Следовательно, надхрящница как составная часть хряща выполняет следующие функции:
1) обеспечивает трофикой бессосудистую хрящевую ткань;
2) защищает хрящевую ткань;
3) обеспечивает регенерацию хрящевой ткани при ее повреждении.
Трофика гиалиновой хрящевой ткани суставных поверхностей обеспечивается синовиальной жидкостью суставов, и жидкостью из сосудов костной ткани.
14. Скелетные соединительные ткани. Костные ткани
Костная ткань является разновидностью соединительной ткани и состоит из клеток и межклеточного вещества, в котором содержится большое количество минеральных солей, фосфат кальция. Минеральные вещества составляют 70 % костной ткани, органические – 30 %.
Функции костных тканей:
1) опорная;
2) механическая;
3) защитная (механическая защита);
4) участие в минеральном обмене организма (депо кальция и фосфора).
Клетки костной ткани – остеобласты, остеоциты, остеокласты. Основными клетками в сформированной костной ткани являются остеоциты. Это клетки отростчатой формы с крупным ядром и слабо выраженной цитоплазмой (клетки ядерного типа). Тела клеток локализуются в костных полостях (лакунах), а отростки – в костных канальцах. Многочисленные костные канальцы, анастомозируя между собой, пронизывают костную ткань, сообщаясь периваскулярным пространством, образуют дренажную систему костной ткани.
Остеоциты являются дефинитивными формами клеток и не делятся. Образуются они из остеобластов.
Остеобласты содержатся только в развивающейся костной ткани. В сформированной костной ткани они содержатся обычно в неактивной форме в надкостнице. В развивающейся костной ткани остеобласты охватывают по периферии каждую костную пластинку.
Эти клетки обеспечивают минерализацию межклеточного вещества посредством выделения солей кальция. Постепенно выделяя межклеточное вещество, они как бы замуровываются и превращаются в остеоциты. Остеобласты, локализующиеся в камбиальном слое надкостницы, находятся в неактивном состоянии, синтетические и транспортные ор-ганеллы в них слабо развиты. При раздражении этих клеток (в случае травм, переломов костей и т. д.) в цитоплазме быстро развиваются зернистая ЭПС и пластинчатый комплекс, происходит активный синтез и выделение коллагена и гликозаминогликанов, формирование органического матрикса (костная мозоль), а затем и формирование дефинитивной костной ткани. За счет деятельности остеобластов надкостницы происходит регенерация костей при их повреждении.
Остеокласты – костеразрушающие клетки, в сформированной костной ткани отсутствуют, но содержатся в надкостнице и в местах разрушения и перестройки костной ткани.
Функциональная активность остеокласта: в центральной зоне основания клетки из цитоплазмы выделяются угольная кислота и протеолитические ферменты. Выделяющаяся угольная кислота вызывает деминерализацию костной ткани, а протеолитиче-ские ферменты разрушают органический матрикс межклеточного вещества. Фрагменты коллагеновых волокон фагоцитируются остеокластами и разрушаются внутриклеточно.
15. Скелетные соединительные ткани. Костные ткани (кость, надкостница, красный костный мозг)
Кость – это орган, основным структурным КПМРП-нентом которого являются костная ткань.
Вместе с суставами и связками, соединяющими кости скелета между собой, и мышцами, приклепле
ными к кости сухожилиями, кости образуют опорно-двигательный аппарат. По форме и строени кости бывают длинные или трубчатые, плоские, или широкие, и короткие (например позвонки).
Кость как орган состоит из таких элементов, как:
1) костная ткань;
2) надкостница;
3) костный мозг (красный, желтый);
4) сосуды и нервы.
Надкостница (периост) окружает по периферии костную ткань (за исключением суставных поверхностей) и имеет строение, сходное с надхрящницей.
Надкостница – тонкая прояная соединительнотканная пластинка, которая богата кровеносными и лимфатическими сосудами, нервами.
Таким образом, вследствие костеобразующихсвойств надкостницы кость растет в толщину. С костью надкостница прочно сращена при помощи проюодающих волокон, уходящих в глубь кости.
Красный костный мозг рассматривается как самостоятельный орган и относится корганам кроветворения и иммуногенеза.
Костная ткань в сформированных костях представлена в основном пластинчатой формой, однако в разных костях, в разных участках одной кости она имеет разное строение. В плоских костях и эпифизах трубчатых костей костные пластинки образуют перекладины (трабекулы), составляющие губчатое вещество кости.
Строение диафиза трубчатой кости. На поперечном срезе диафиза трубчатой кости различают следующие слои:
1) надкостницу (периост);
2) наружный слой общих, или генеральных, пластин;
3) слой остеонов;
4) внутренний слой общих, или генеральных, пластин;
5) внутреннюю фиброзную пластинку (эндост). Классификация костных тканей. Различают две разновидности костных тканей:
1) ретикулофиброзную (грубоволокнистую);
2) пластинчатую (параллельно волокнистую).
В основе классификации лежит характер расположения коллагеновых волокон. В минерализованном межклеточном веществе в лакунах беспорядочно располагаются остеоциты. Пластинчатая костная ткань состоит из костных пластинок, в которых коллагено-вые волокна или их пучки располагаются параллельно в каждой пластинке, но под прямым углом к ходу волокон соседних пластинках. Между пластинками в лакунах располагаются остеоциты, тогда как их отростки проходят в канальцах через пластинки.
В организме человека костная ткань представлена почти исключительно пластинчатой формой. Ретику-лофиброзная костная ткань встречается только как этап развития некоторых костей (теменных, лобных).
16. Мышечные ткани
Скелетная мышечная ткань
Гладкая мышечная ткань, развивается из мезенхимы. К специальным мышечным тканям относятся гладкомышечные клетки радужной оболочки, – миоэ-пителиальные клетки слюнных, слезных, потовых и молочных желез.
Поперечнополосатая мышечная ткань подразделяется на скелетную и сердечную. Обе эти разновидности развиваются из мезодермы, но из разных ее частей: скелетная – из миотомов сомитов, сердечная – из висцеральных листков спланхиотомов.
Структурно-функциональной единицей поперечнополосатой скелетной мышечной ткани является мышечное волокно. Оно представляет собой вытянутое цилиндрическое образование с заостренными концами. Мышечное волокно окружено оболочкой сарколеммой, в которой под электронным микроскопом отчетливо выделяются два листка: внутренний листок является типичной плазмолеммой, а наружный представляет собой тонкую соединительно-тканную пластинку (базальную пластинку).
Базальная пластинка образована тонкими колла-геновыми и ретикулярными волокнами, относится к опорному аппарату и выполняет вспомогательную функцию передачи сил сокращения на соединительно-тканные элементы мышцы.
Миосимпласт является основным структурным компонентом мышечного волокна (как по объему, так и по выполняемым функциям). Он образуется посредством слияния самостоятельных недифференцированных мышечных клеток – миобластов. Отличительной особенностью миосимпласта является также наличие в нем:
1) миофибрилл;
2) саркоплазматической сети;
3) канальцев Т-системы.
Миофибриллы – сократительные элементы мио-симпласта, локализуются в центральной части саркоплазмы миосимпласта.
По своему строению миофибриллы неоднородны по протяжению, подразделяются на темные (анизотропные), или А-диски, и светлые (изотропные), или I-диски.
Саркоплазматическая сеть – это видоизмененная гладкая эндоплазматическая сеть; состоящая из расширенных полостей и анастомозирующих канальцев, окружающих миофибриллы.
Мышца состоит из мышечных волокон, волокнистой соединительной ткани, сосудов, нервов. В мышечной ткани различают два вида регенерации – физиологическую и репаративную. Физиологическая регенерация проявляется форме гипертрофии мышечных волокон.
Репаративная регенерация развивается после повреждения мышечных волокон.
В условиях небольшого дефекта мышечного волокна на его концах за счет регенерации внутриклеточных органелл образуются мышечные точки, которые растут навстречу друг другу, затем сливаются, приводя к закрытию дефекта.
Скелетные мышцы получают двигательную, чувствительную и трофическую иннервацию.
17. Мышечные ткани. Сердечная и гладкая мышечные ткани
Сердечная мышечная ткань
Структурно-функциональной единицей сердечной поперечнополосатой мышечной ткани является кар-диомиоцит. По строению и функциям кардиомиоциты подразделяются на две группы:
1) типичные, или сократительные, кардиомиоциты, образующие своей совокупностью миокард;
2) атипичные кардиомиоциты, составляющие проводящую систему сердца.
Сократительный кардиомиоцит представляет собой почти прямоугольную клетку в центре которой локализуется обычно одно ядро.
Атипичные кардиомиоциты образуют проводящую систему сердца, которая включает в себя следующие структурные компоненты:
1) синусо-предсердный узел;
2) предсердно-желудочковый узел;
3) предсердно-желудочковый пучок (пучок Гисса) – ствол, правую и левую ножки;
4) концевые разветвления ножек (волокна Пуркинье). Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
Источники развития кардиомиоцитов – миоэпикар-диальные пластинки, представляющие собой определенные участки висцеральных спланхиотомов.
Гладкая мышечная ткань мезенхимального происхождения
Локализуется в стенках полых органов (желудка, кишечника, дыхательных путей, органов мочеполовой системы) и в стенках кровеносных и лимфатических сосудов. Структурно-функциональной единицей является миоцит: клетка веретенообразной формы длиной 30—100 мкм (в беременной матке – до 500 мкм), диаметром 8 мкм, покрытая базальной пластинкой.
Миозиновые и актиновые филаменты составляют сократительный аппарат миоцита.
Эфферентная иннервация гладкой мышечной ткани осуществляется вегетативной нервной системой.
Сокращение гладкомышечной ткани обычно бывает длительным, что обеспечивает поддержание тонуса полых внутренних органов и сосудов.
Гладкомышечная ткань не образует мышцы в анатомическом понимании этого слова. Однако в полых внутренних органах и в стенке сосудов между пучками миоцитов содержатся прослойки рыхлой волокнистой соединительной ткани, образующие своеобразный эндомизий, а между пластами гладкой мышечной ткани – перимизий.
Регенерация гладкомышечной ткани осуществляется несколькими способами:
1) посредством внутриклеточной регенерации (гипертрофии при усилении функциональной нагрузки);
2) посредством митотического деления миоцитов (пролиферации);
3) посредством дифференцировки из камбиальных элементов (из адвентициальных клеток и миофи-бробластов).
18. Нервная ткань
Структурно-функциональные особенности нервной ткани:
1) состоит из двух основных типов клеток: нейроци-тов и нейроглии;
2) межклеточное вещество отсутствует;
3) нервная ткань не подразделяется на морфологические подгруппы;
4) основной источник происхождения: нейроэкто-дерма.
Структурные компоненты нервной ткани:
1) нервные клетки (нейроциты или нейроны);
2) глиальные клетки – глиоциты.
Нейроциты – это структурные компоненты нервной ткани. Клетки нейроглии способствуют выполнению перечисленных функций.
Источники и этапы развития нервной ткани
Основной источник – нейроэктодерма. Некоторые клетки глиальные клетки развиваются из микроглии и из мезенхимы.
Этапы развития:
1) нервная пластинка;
2) нервный желобок;
3) нервная трубка, ганглиозная пластинка, нейраль-ные плакоды.
Из нервной трубки развивается нервная ткань, в основном – из органов центральной нервной системы (спинного и головного мозга). Из ганглиозной пластинки развивается нервная ткань некоторых органов периферической нервной системы (вегетативных и спинальных ганглиев). Из нейральных плакод развиваются ганглии черепных нервов. В процессе развития нервной ткани вначале образуются два типа клеток:
1) нейробласты;
2) глиобласты. Характеристика нейроцитов
По морфологии все нейроциты являются отростча-тыми клетками. в каждой нервной клетке выделяют две части:
1) клеточное тело (перикарион);
2) отростки.
Отростки нейроцитов подразделяются на две разновидности:
1) аксон, который проводит импульсы от клеточного тела (на другие нервные клетки или на рабочие органы);
2) дендрит, который проводит импульсы к клеточному телу.
Классификация нейроцитов Нервные клетки классифицируются:
1) по морфологии;
2) по функции.
По морфологии по количеству отростков подразделяются на:
1) униполярные (псевдоуниполярые) с одним отростком;
2) биполярные (с двумя отростками);
3) мультиполярные (более двух отростков). По функции подразделяются на:
1) афферентные (чувствительные);
2) эфферентные (двигательные, секреторные);
3) ассоциативные (вставочные);
4) секреторные (нейроэндокринные).
19. Нервная ткань (продолжение)
Клетки нейроглии являются вспомогательными клетками и нервной ткани и выполняют следующие функции:
1) опорную;
2) трофическую;
3) разграничительную;
4) секреторную;
5) защитную и др.
Глиальные клетки по своей морфологии также являются отростчатыми клетками, не одинаковыми по величине, форме и количеству отростков. На основании размеров они подразделяются, на макроглию и микроглию. Клетки макроглии имеют эктодермаль-ный источник происхождения (из нейроэктодермы), клетки микроглии развиваются из мезенхимы.
Эпендимоциты выполняют следующие функции в нервной системе:
1) разграничительную (образуя выстилку полостей мозга);
2) секреторную;
3) механическую (обеспечивает движение церебральной жидкости);
4) опорную (для нейроцитов);
5) барьерную (участвуя в образовании поверхностной глиальной пограничной мембраны).
Астроциты – клетки с многочисленными отростками, напоминающими в совокупности форму звезды, откуда и происходит их название. По особенностям строения их отростков астроциты подразделяются на:
1) протоплазматические (короткие, но широкие и сильно ветвящиеся отростки);
2) волокнистые (тонкие, длинные, слабо ветвящиеся отростки).
Волокнистые астроциты осуществляют опорную функцию для нейроцитов и их отростков, так как их длинные тонкие отростки образуют глиальные волокна. Кроме того, терминальные расширения отростков волокнистых астроцитов образуют периваскулярные (вокругсосудистые) глиальные пограничные мембраны, являющиеся одним из структурных компонентов гематоэнцефалического барьера.
Олигодендроциты – малоотростчатые клетки, самая распространенная популяция глиоцитов. Локализуются они преимущественно в периферической нервной системе и в зависимости от области локализации подразделяются на:
1) мантийные глиоциты (окружают тела нервных клеток в нервных и вегетативных ганглиях);
2) леммоциты, или шванновские клетки (окружают отростки нервных клеток, вместе с которыми образуют нервные волокна);
3) концевые глиоциты (сопровождают концевые ветвления дендритов чувствительных нервных клеток).
Микроглия представлена мелкими отростчатыми клетками, выполняющими защитную функцию – фагоцитоз. На основании этого их называют глиальными макрофагами. Большинство исследователей считают, что глиальные макрофаги (как и любые другие макрофаги) являются клетками мезенхимального происхождения.
20. Нервные волокна
Нервные волокна представляют собой комплексные образования, включающие следующие элементы:
1) отростки нервных клеток;
2) глиальные клетки;
3) соединительно-тканную пластинку.
Главной функцией нервных волокон является проведение нервных импульсов. Отростки нервных клеток проводят нервные импульсы, а глиальные клетки способствуют этому проведению.
По особенностям строения и функции нервные волокна подразделяются на две разновидности:
1) безмиелиновые;
2) миелиновые.
Безмиелиновое нервное волокно представляет собой цепь леммоцитов, в которую вдавлено несколько осевых цилиндров.
Строение миелинового нервного волокна. Миели-новое нервное волокно имеет те же структурные компоненты, что и безмиелиновое, но отличается рядом особенностей:
1) осевой цилиндр один и погружается в центральную часть цепи леммоцита;
2) мезаксон длинный и закручен вокруг осевого цилиндра, образуя миелиновый слой;
3) цитоплазма и ядро леммоцитов сдвигаются на периферию и составляют нейролемму миелинового нервного волокна;
4) на периферии расположена базальная пластинка. На поперечном сечении миелинового нервного волокна видны следующие структурные элементы:
1) осевой цилиндр;
2) миелиновый слой;
3) неврилемма;
4) базальная пластинка.
По ходу миелинового нервного волокна видны границы соседних леммоцитов – узловые перехваты (перехваты Ранвье), а также участки между двумя перехватами (межузловые сегменты), каждый из которых соответствует протяженности одного леммоцита. В каждом межузловом сегменте отчетливо прослеживаются насечки миелина – прозрачные участки.
Высокая скорость проведения нервных импульсов по миелиновым нервным волокнам объясняется саль-таторным способом проведения нервных импульсов: скачками от одного перехвата к другому.
Нервное волокно это совокупность нервных и гли-альных клеток, после его повреждения отмечается реакция. После пересечения наиболее заметные изменения проявляются в дистальном отделе нервного волокна, где отмечается распад осевого цилиндра. Леммоциты, окружающие этот участок осевого цилиндра, не погибают, а округляются, пролиферируют и образуют тяж глиальных клеток по ходу распавшегося нервного волокна.
В перикарионе нервной клетки с отсеченным отростком проявляются признаки раздражения: набухание ядра и сдвиг его на периферию клетки, расширение перинуклеарного пространства.
В проксимальном отделе нервного волокна на конце осевого цилиндра образуется расширение: колба роста, которая постепенно врастает в тяж глиальных клеток.
21. Нервная ткань (нерв, нервные окончания)
Не следует смешивать понятия «нервное волокно» и «нерв».
Нерв – комплексное образование, состоящее из:
1) нервных волокон;
2) рыхлой волокнистой соединительной ткани, образующей оболочки нерва.
Среди оболочек нерва различают:
1) эндоневрий (соединительную ткань, окружающую отдельные нервные волокна);
2) периневрий (соединительную ткань, окружающую пучки нервных волокон);
3) эпиневрий (соединительную ткань, окружающую нервный ствол).
Нервные окончания, или концевые нервные аппараты. Представляют собой окончания нервных волокон. Если осевой цилиндр нервного волокна является дендритом чувствительной нервной клетки, то его концевой аппарат образует рецептор. Если осевой цилиндр является аксоном нервной клетки, то его концевой аппарат образует эффекторное или синаптическое окончание. Следовательно, нервные окончания подразделяются на три основные группы:
1) эффекторные (двигательные или секреторные);
2) рецептурные (чувствительные);
3) синаптические.
Двигательное нервное окончание – концевой аппарат аксона на поперечно-полосатом мышечном волокне или на миоците. В нем различают три части:
1) нервный полюс;
2) синаптическую щель;
3) мышечный полюс.
В каждом терминальном ветвлении аксона содержатся следующие структурные элементы:
1) пресинаптическая мембрана;
2) синаптические пузырьки с медиатором (ацетилхо-лином);
3) скопление митохондрий с продольными кристами. Мышечный полюс, или полотна моторной бляшки,
включает:
1) постсинаптическую мембрану – специализированный участок плазмолеммы миосимпласта, содержащий белки-рецепторы к ацетилхолину;
2) участок саркоплазмы миосимпласта, в котором отсутствуют миофибриллы и содержится скопление ядер и саркосом.
Рецепторные нервные окончания классифицируются по нескольким признакам:
1) по локализации:
а) интеророцепторы (рецепторы внутренних органов);
б) экстрорецепторы (воспринимают внешние раздражители: репетиры кожи, органов чувств);
в) проприорецепторы (локализуются в аппарате движения);
2) по специфичности восприятия (по модальности):
а) хеморецепторы;
б) механорецепторы;
в) барореценторы;
г) терморецепторы (тепловые, холодовые);
3) по строению:
а) свободные;
б) несвободные (инкапсулированные, не инкапсулированные).
22. Нервная система
С анатомической точки зрения нервную систему делят на центральную и периферическую. Спинномозговые узлы
Первым нейроном каждой рефлекторной дуги является рецепторная нервная клетка. Большая часть этих клеток сконцентрирована в спинномозговых узлах, расположенных по ходу задних корешков спинного мозга. Спинномозговой узел окружен соединительнотканной капсулой. От капсулы в паренхиму узла проникают тонкие прослойки соединительной ткани, которая образует его остов, по нему проходят в узле кровеносные сосуды.
Дендриты и нейриты клеток в узле и за его пределами покрыты оболочками из леммоцитов. Нервные клетки спинномозговых узлов окружены слоем клеток.
Спинной мозг. Представляется собой две симметричные половины, отграниченных друг от друга спереди глубокой серединной щелью, а сзади – соединительно-тканной перегородкой.
Внутренняя часть спинного мозга темнее – это его серое вещество. По периферии его располагается более светлое белое вещество. Серое вещество на поперечном сечении мозга видно в виде бабочки. Выступы серого вещества принято называть рогами. Различают передние, или вентральные, задние, или дорсальные, и боковые, или латеральные, рога.
Серое вещество спинного мозга состоит из мульти-полярных нейронов, безмиелиновых и тонких миели-новых волокон и нейроглии.
Белое вещество спинного мозга образуется совокупностью продольно ориентированных преимущественно миелиновых волокон нервных клеток.
В средней части заднего рога спинного мозга располагается собственное ядро заднего рога. Оно состоит из пучковых клеток, аксоны которых, переходя через переднюю белую спайку на противоположную сторону спинного мозга в боковой канатик белого вещества, образуют вентральный спиномоз-жечковый и спиноталамический пути и направляются в мозжечок и зрительный бугор.
В задних рогах диффузно расположены вставочные нейроны. Это мелкие клетки, аксоны которых заканчиваются в пределах серого вещества спинного мозга той же (ассоциативные клетки) или противоположной (комиссуральные) стороны.
Дорсальное ядро, или ядро Кларка, состоит из крупных клеток с разветвленными дендритами. Их аксоны пересекают серое вещество, входят в боковой канатик белого вещества той же стороны и в составе дорсального спиномозжечкового пути поднимаются к мозжечку.
Самые крупные нейроны спинного мозга находятся в передних рогах, они также образуют ядра из тел нервных клеток, корешки которых образуют основную массу волокон передних корешков.
Белое вещество спинного мозга состоит из миели-новых волокон, идущих продольно. Пучки нервных волокон, осуществляющие связь между различными отделами нервной системы, называются проводящими путями спинного мозга.
23. Нервная система. Головной мозг
В головном мозге также выделяют серое и белое вещество, но распределение этих двух составных частей здесь более сложное, чем в спинном мозге.
Ствол мозга. Все ядра серого вещества ствола мозга состоят из мультиполярных нервных клеток. На них имеются окончания нейритов клеток спинальных ганглиев. в стволе головного мозга имеется большое количество ядер, предназначенных для переключения нервных импульсов из спинного мозга и ствола на кору и от коры на собственный аппарат спинного мозга.
В продолговатом мозге имеется большое количество ядер собственного аппарата черепных нервов, которые в основном находятся в дне IV желудочка, в продолговатом мозге имеются ядра, которые переключают поступающие в него импульсы на другие отделы головного мозга (нижние оливы).
Ретикулярная субстанция представляет собой сложный рефлекторный центр, связанный со спинным мозгом, мозжечком, корой большого мозга и гипота-ламической областью.
Мост головного мозга состоит из большого количества поперечно идущих нервных волокон и лежащих между ними ядер. В базальной части моста поперечные волокна пирамидными путями раздвигаются на две группы – заднюю и переднюю.
Средний мозг состоит из серого вещества четверохолмия и ножек мозга, которые образованы массой миелиновых нервных волокон, идущих от коры большого мозга. Покрышка содержит центральное серое вещество, состоящее из крупных мультиполярныхи более мелких веретенообразных клеток и волокон.
Промежуточный мозг представляет собой зрительный бугор. Вентрально от него располагается богатая мелкими ядрами гипоталамическая (подбугор-ная) область. В вентральных ядрах таламической области заканчиваются восходящие чувствительные пути, от них нервные импульсы передаются коре.
В каудальной группе ядер (в подушке зрительного бугра) заканчиваются волокна зрительного пути.
Гипоталамическая область представляет собой вегетативный центр головного мозга, регулирующий основные обменные процессы: температуру тела, кровяное давление, водный, жировой обмен и др.
Функция мозжечка это обеспечение равновесия и координации движений. Он имеет связь со стволом мозга с помощью афферентных и эфферентных проводящих путей, образующих в совокупности три пары ножек мозжечка. На поверхности мозжечка расположено множество извилин и бороздок.
Серое вещество образует кору мозжечка, меньшая его часть лежит глубоко в белом веществе в виде центральных ядер. В центре каждой извилины имеется тонкая прослойка белого вещества, покрытая слоем серого вещества – корой.
В коре мозжечка имеется три слоя: наружный (молекулярный), средний (ганглионарный) и внутренний (зернистый).
Эфферентные нейроны коры мозжечка – грушевидные клетки, или клетки Пуркинье, составляют ган-глионарный слой.
Молекулярный слой состоит из двух основных видов нервных клеток: корзинчатых и звездчатых.
24. Нервная система. Кора большого мозга. Цитоорхитектоника коры большого мозга
Кора большого мозга. Представлена слоем серого вещества
Цитоархитектоника коры большого мозга. Нейроны коры весьма разнообразны по форме, они являются мультиполярными клетками. Они делятся на пирамидные, звездчатые, веретенообразные, паукообразные и горизонтальные нейроны.
Пирамидные нейроны составляют основную часть коры большого мозга. Их тело имеет форму треугольника, вершина которого обращена к поверхности коры. От вершины и боковых поверхностей тела отходят дендриты, заканчивающиеся в различных слоях серого вещества. От основания пирамидных клеток берут начало нейриты, в одних клетках короткие, образующие ветвления в пределах данного участка коры, в других – длинные, поступающие в белое вещество.
Пирамидные клетки различных слоев коры различны. Мелкие клетки представляют собой вставочные нейроны, нейриты которых связывают отдельные участки коры одного полушария (ассоциативные нейроны) или двух полушарий (комиссуральные нейроны).
Крупные пирамиды и их отростки образуют пирамидные пути, проецирующие импульсы в соответствующие центры ствола и спинного мозга.
В каждом слое клеток коры головного мозга имеется преобладание каких-либо видов клеток. Выделяется несколько слоев:
первый – молекулярный,
второй – наружный зернистый,
третий – пирамидный,
четвертый – внутренний зернистый, пятый – ганглионарный,
шестой – слой полиморфных клеток.
В молекулярном слое коры содержится небольшое количество мелких клеток веретенообразной формы. Наружный зернистый слой это скопление мелких нейронов, имеющих различную форму. Дендриты этих клеток поднимаются в молекулярный слой, а аксоны уходят в белое вещество или, образуя дуги, идут в тангенциальное сплетение волокон молекулярного слоя.
Пирамидный слой – самый большой по толщине, очень хорошо развитый в прецентральной извилине. От верхушки пирамидной клетки отходит главный дендрит, который располагается в молекулярном слое. Ганглионарный слой коры состоит из крупных пирамидных клеток, причем область прецентральной извилины содержит гигантские пирамиды, (клетки Беца). Для гигантских пирамид характерно наличие крупных глыбок базофильного вещества. Нейриты клеток этого слоя образуют главную часть кортико-спинальных путей спинного мозга и оканчиваются синапсами на клетках его моторных ядер.
Слой полиморфных клеток образован нейронами веретенообразной формы. Нейроны внутренней зоны более мелкие и лежат на большом расстоянии друг от друга, а внешней – более крупные. Нейриты клеток полиморфного слоя уходят в белое вещество в составе эфферентных путей головного мозга. Дендриты достигают молекулярного слоя коры.
25. Нервная система коры большого мозга. Миелоархитехтоника коры
Миелоархитектоника коры. В больших полушарий можно выделить следующие типы волокон: ассоциативные волокна, комиссуральные и проекционные волокна – как афферентные, так и эфферентные (связывают кору с ядрами низших отделов центральной нервной системы).
Вегетативная (или автономная) нервная система по различным свойствам делится на симпатическую и парасимпатическую. Вегетативная нервная система также состоит из центральных отделов, представленных ядрами серого вещества головного спинного мозга, и периферических отделов: нервных узлов и сплетений. Ядра центрального отдела вегетативной нервной системы находятся в среднем и продолговатом мозге, а также в боковых рогах грудных, поясничных и сакральных сегментов спинного мозга. Ядра кранио-бульбарного и сакрального отделов относятся к парасимпатической, а ядра тораколюмбального отдела – к симпатической нервной системе. Мульт-иполярные нервные клетки этих ядер представляют собой ассоциативные нейроны рефлекторных дуг вегетативной нервной системы. Их отростки выходят из центральной нервной системы через передние корешки или черепные нервы и оканчиваются синапсами на нейронах одного из периферических ганглиев. Это преганглионарные волокна вегетативной нервной системы. Преганглионарные волокна симпатической и парасимпатической вегетативной нервной системы – холинергические. Аксоны нервных клеток периферических нервных узлов выходят из ганглиев в виде постганглионарных волокон и образуют концевые аппараты в тканях рабочих органов. В его состав входят центральные нейроны с их аксонами в виде преганглионарных волокон и периферические нейроны, расположенные в периферических узлах. Преганглионарные волокна в большинстве случаев покрыты миелиновой оболочкой, чем и объясняется белый цвет связующих ветвей, несущих симпатические преганглионарные волокна от передних корешков к ганглиям симпатического пограничного столба. Постганглионарные волокна тоньше и в большинстве случаев не имеют миелиновой оболочки: это волокна серых связующих ветвей, идущие от узлов симпатического пограничного ствола к периферическим спинномозговым нервам.
Оболочки головного и спинного мозга. Головной и спинной мозг покрыты тремя видами оболочек: мягкой (непосредственно прилегающей к тканям мозга), паутинной и твердой (граничит с костной тканью черепа и позвоночника). Мягкая мозговая оболочка покрывает ткань мозга, она отграничена от нее лишь краевой глиальной мембраной. В этой оболочке имеются в большом количество кровеносных сосудов, питающих мозг, и многочисленные нервные волокна, концевые аппараты и одиночные нервные клетки. Паутинная оболочка представляет собой очень нежный рыхлый слой волокнистой соединительной ткани. Между ней и мягкой мозговой оболочкой лежит су-барахноидальное пространство, которое сообщается с желудочками мозга и содержит цереброспинальную жидкость. Твердая мозговая оболочка образована плотной волокнистой соединительной тканью, и состоит из эластических волокон. В полости черепа она плотно сращена с надкостницей.
26. Сердечно-сосудистая система. Кровеносные сосуды
Кровеносные сосуды
Это замкнутые в виде кольца трубочки различного диаметра, осуществляющие транспортную функцию, а также налаживающие кровоснабжение органов и обмен веществ между кровью и окружающими тканями.
К артериям относят сосуды, по которым кровь идет от сердца к органам. Эта кровь насыщена кислородом, исключением являются системы легочной артерии, несущей венозную кровь. К венозным относят сосуды, по которым кровь идет к сердцу и содержит мало кислорода,
Гемокапилляры соединяют артериальное звено кровеносной системы с венозным, помимо сетей, капилляры которых располагаются либо между двумя артериями, либо между двумя венами.
Все артерии делятся на три типа: эластический, мышечный и смешанный (мышечно-эластический). Стенки всех артерий и вен состоят из трех оболочек: внутренней, средней и наружной. К артериям эластического типа причисляют сосуды крупного калибра (аорту и легочную артерию): функция этих сосудов – транспортная. Высокое давление и большая скорость протекающей крови определяют строение стенки сосудов эластического типа. Так, внутренняя оболочка крупных артерий включает эндотелий с базальной мембраной, далее идет подэндотелиальный слой и сплетение эластических волокон. Подэндотелиальный слой представлен рыхлой тонкофибриллярной соединительной тканью, богатой малодифференцированными клетками звездчатой формы. Межклеточное вещество внутренней оболочки крупного сосуда или (реже) других оболочкек содержит большое количество гликозаминогликанов и фосфолипиды, обнаруживаемых при соответствующей обработке. Средняя оболочка крупного сосуда состоит из большого количества эластических окончатых мембран, связанных посредством эластических волокон.
Наружная оболочка состоит из рыхлой волокнистой соединительной ткани, имеющей множество эластических и коллагеновых волокон с продольным направлением.
Средняя оболочка артерий смешанного типа имеет одинаковое количество гладких мышечных клеток, эластических волокон и окончатых эластических мембран. К артериям мышечного типа относятся преимущественно артерии тела, конечностей и внутренних органов среднего и мелкого калибра, т. е. большинство артерий организма. Особенностью является большое количество гладких мышечных клеток, которые обеспечивают дополнительную нагнетательную силу и регулируют приток крови к органам. Внутренняя оболочка состоит из эндотелия, подэнтелиально-го слоя и внутренней эластической мембраны.
Кровеносные капилляры – наиболее многочисленные и самые тонкие сосуды, однако диаметр их просвета может варьировать. Это обусловлено как органными особенностями капилляров, так и функциональным состоянием сосудистой системы. Площадь поперечного сечения среза капиллярного русла в любой области во много раз превышает площадь поперечного среза исходной артерии. В стенке капилляров различают три тонких слоя как рудименты трех оболочек сосудов.
27. Сердечно-сосудистая система
Артериоловенулярные анастомозы это соединения сосудов, несущих артериальную и венозную кровь в обход капиллярного русла. Их наличие отмечается почти во всех органах.
Различают две группы анастомозов:
1) истинные артериоловенулярные анастомозы (шунты), по которым сбрасывается чистая артериальная кровь;
2) атипичные артериоловенулярные соустья (полушунты), по которым течет смешанная кровь.
Внешняя форма первой группы анастомозов может быть различной: в виде прямых коротких соустьев, петлеобразных, иногда в виде ветвящихся соединений.
В гистоструктурном отношении они подразделяются на две подгруппы:
а) сосуды, не имеющие специальных запирательных устройств;
б) сосуды, снабженные специальными сократительными структурами.
Во второй подгруппе анастомозы имеют специальные сократительные сфинктеры в виде продольных валиков или подушек в подэндотелиальном слое. Сокращение мышечных подушек, выступающих в просвет анастомоза, приводит к прекращению кровотока. Простые анастомозы эпителиоидного типа характеризуются наличием в средней оболочке внутреннего продольного и наружного циркулярного слоев гладких мышечных клеток, которые по мере приближения к венозному концу заменяются на короткие овальные светлые клетки, похожие на эпителиальные, способные к набуханию и отбуханию, благодаря чему происходит изменение просвета анастомоза. В венозном, сегменте артериоловенулярного анастомоза стенка его резко истончается. Наружная оболочка состоит из плотной соединительной ткани. Арте-риоловенулярные анастомозы, особенно клубочково-го типа, богато иннервированы.
Строение вен тесно связано с гемодинамическими условиями их функционирования. Количество же гладких мышечных клеток в стенке вен неодинаково и зависит от того, движется ли в них кровь к сердцу под действием силы тяжести или против нее. По степени развития мышечных элементов в стенке вен они могут быть разделены на две группы: вены безмышечного типа и вены мышечного типа. Вены мышечного типа, в свою очередь, подразделяются на вены со слабым развитием мышечных элементов и вены со средним и сильным развитием мышечных элементов. В венах (также, как и в артериях), различают три оболочки: внутреннюю, среднюю и наружную, при этом степень выраженности этих оболочек в венах существенно отличается. Вены безмышечного типа – это вены твердой и мягкой мозговых оболочек, вены сетчатки глаза, костей, селезенки и плаценты. Под действием крови эти вены способны к растяжению, но скопившаяся в них кровь сравнительно легко под действием собственной силы тяжести оттекает в более крупные венозные стволы. Вены мышечного типа отличаются развитием в них мышечных элементов. К таким венам относятся вены нижней части туловища. Также в некоторых видах вен имеется большое количество клапанов, что препятствует обратному току крови под силой собственной тяжести.
28. Лимфатические сосуды
По лимфатическим сосудам происходит отток лимфы в венозное русло. К лимфатическим сосудам относят лимфатические капилляры, интра– и экстраорганные лимфатические сосуды, отводящие лимфу от органов, и лимфатические стволы тела, к которым относятся грудной проток и правый лимфатический проток, впадающие в крупные вены шеи.
Лимфатические капилляры являются началом лимфатической системы сосудов, в которые поступают из тканей продукты обмена веществ, а в патологических случаях – инородные частицы и микроорганизмы. Лимфатические капилляры представляют собой систему замкнутых и анастомозирующих друг с другом и пронизывающих весь организм. Стенка лимфатических капилляров представлена эндотелиальными клетками, которые, в отличие от подобных клеток кровеносных капилляров, не имеют базальной мембраны. По размерам диаметра все лимфатические сосуды делятся на мелкие, средние и крупные. Эти сосуды по своему строению могут быть безмышечными и мышечными. Мелкие сосуды являются внутриорганны-ми лимфатическими сосудами, мышечные элементы в них отсутствуют, и их эндотелиальная трубка окружена только соединительно-тканной оболочкой. Средние и крупные лимфатические сосуды имеют три хорошо развитые оболочки: внутреннюю, среднюю и наружную. Во внутренней оболочке, покрытой эндотелием, находятся продольно и косо направленные пучки коллагеновых и эластических волокон.
На внутренней оболочке сосудов имеются клапаны. Они состоят из центральной соединительно-тканной пластинки, покрытой с внутренней и наружной поверхностей эндотелием. Мышечный слой стенки лимфатического сосуда достигает хорошего развития в коллекторах подвздошного лимфатического сплетения, около аортальных лимфатических сосудов и шейных лимфатических стволов, сопровождающих яремные вены. Наружная оболочка лимфатических сосудов образована рыхлой волокнистой неоформленной соединительной тканью, которая без резких границ переходит в окружающую соединительную ткань.
Васкуляризация. Внутренняя оболочка артерий получает питательные вещества непосредственно из крови, протекающей в данной артерии. В диффузии питательных веществ через внутреннюю оболочку артерий большую роль играют белково-мукополисаха-ридные комплексы, входящие в состав основного вещества стенок этих сосудов. Иннервация сосудами получается от вегетативной нервной системы. Нервные волокна этого отдела нервной системы, как правило, сопровождают сосуды и заканчиваются в их стенке. По строению нервы сосудов являются либо миелиновыми, либо безмиелиновыми.
Регенерация. Кровеносные и лимфатические сосуды обладают высокой способностью к восстановлению как после травм, так и после различных патологических процессов, происходящих в организме. Восстановление дефектов сосудистой стенки после ее повреждения начинается с регенерации и роста ее эндотелия. Мышечные пучки поврежденного сосуда, восстанавливаются более медленно и неполно по сравнению с другими тканевыми элементами сосуда.
29. Сердце
Сердце представляет собой полый орган, состоящий из двух предсердий и двух желудочков. Стенка его состоит их трех оболочек: внутренней (эндокарда), средней, или мышечной (миокарда), и наружной, или серозной (эпикарда).
Внутренняя оболочка сердца – эндокард – изнутри покрывает все камеры сердца, а также клапаны сердца.
Эндокард состоит из нескольких видов клеток. Так, на стороне, обращенной в полость сердца, эндокард выстлан эндотелием, состоящим из полигональных клеток. Далее идет подэндоте-лиальный слой, образованный соединительной тканью, богатой малодиф-ференцированными клетками.
Мышечный слой клеток – миокард. Мышечные волокна миокарда прикрепляются к опорному скелету сердца, который образован фиброзными кольцами между предсердиями и желудочками и плотной соединительной тканью в устьях крупных сосудов. Наружная оболочка сердца, эпикард, висцеральный листок перикарда, сходный по строению с серозными оболочками. Между перикардом и эпикардом имеется щелевидная полость, в которой находится небольшое количество жидкости, благодаря которой при сокращении сердца уменьшается сила трения.
Между предсердиями и желудочками сердца, а также желудочками и крупными сосудами располагаются клапаны. При этом они имеют специфические названия. Так, предсердно-желудочковый (атриовентрику-лярный) клапан в левой половине сердца – двустворчатый (митральный), в правой – трехстворчатый. Они представляют собой покрытые эндотелием тонкие пластинки плотной волокнистой соединительной ткани с небольшим количеством клеток.
Внутренний слой, обращенный к желудочку сердца, представляет собой продолжение эндокарда. В нем под эндотелием продольно и поперечно идут эластические волокна, за которыми следует смешанная эластико-коллагеновая прослойка.
Средний слой тонкий, состоит из рыхлой волокнистой соединительной ткани, богатой клеточными элементами.
Наружный слой, обращенный к аорте, содержит коллагеновые волокна, которые берут начало от фиброзного кольца вокруг аорты.
Питательные вещества сердце получает из системы венечных артерий.
Кровь из капилляров собирается в коронарные вены, впадающие в правое предсердие или венозный синус. Лимфатические сосуды в эпикарде сопровождают кровеносные.
Иннервация. В оболочках сердца обнаруживаются несколько нервных сплетений и небольшие нервные ганглии. Среди рецепторов имеются как свободные, так и инкапсулированные окончания, располагающиеся в соединительной ткани, на мышечных клеткахи в стенке венечных сосудов.
Тела чувствительных нейронов лежат в спинномозговых узлах (С7-^6), а их аксоны, покрытые миелиновой оболочкой, вступают в продолговатый мозг.
30. Эндокринная система
К эндокринной системе относятся железы, не имеющие выводных протоков, но выделяющие во внутреннюю среду организма высокоактивные биологические вещества, действующие на клетки, ткани и органы вещества (гормоны), стимулирующие или ослабляющие их функции.
Связь нервной и эндокринной систем
Гормональным дистантным влияниям принадлежит преимущественная роль в регуляции таких общих функций организма, как обмен веществ, соматический рост, репродуктивные функции. Совместное участие нервной и эндокринной систем в обеспечении регуляции и координации функций организма определяется тем, что регуляторные влияния, оказываемые как нервной, так и эндокринной системами.
Вместе с тем все нервные клетки проявляют способность синтезировать белковые вещества, о чем свидетельствуют сильное развитие гранулярной эн-доплазматической сети и обилие рибонуклеопротеи-дов в их перикарионах. Аксоны таких нейронов, как правило, заканчиваются на капиллярах, и синтезированные продукты, аккумулировавшиеся в терминалях, выделяются в кровь, с током которой разносятся по организму и оказывают в отличие от медиаторов не локальное, а дистантное регулирующее действие, подобно гормонам эндокринных желез. нервные клетки получили наименование нейросекреторных, а вырабатываемые и выделяемые ими продукты – нейро-гормонов. Нейросекреторные клетки, воспринимая, как всякий нейроцит, афферентные сигналы от других отделов нервной системы, посылают свои эфферентные импульсы через кровь, в эндокринной системе выделяют следующие группы желез внутренней секреции: нейроэндокринные трансмиттеры (секреторные ядра гипоталамуса, эпифиз), которые с помощью своих гормонов переключают информацию, поступающую в центральную нервную систему, на центральное звено регуляции аденогипо-физзависимых желез (аденогипофиз) и нейрогемаль-ный орган (задняя доля гипофиза, или нейрогипофиз). Аденогипофиз благодаря гормонам гипоталамуса (либеринам и статинам) выделяет адекватное количество тропных гормонов, которые стимулируют функцию аденогипофиззависимых желез (коры надпочечников, щитовидной и половой желез). Взаимоотношения аденогипофиза и зависимых от него желез внутренней секреции осуществляются по принципу обратной связи, или плюс-минус. Нейроге-мальный орган собст-венных гормонов не продуцирует, но накапливает гормоны крупноклеточных ядер гипоталамуса (окситоцин, АДГ-вазопрессин), затем выделяют их в кровяное русло и таким образом регулируют деятельность так называемых органов-мишеней (матки, почек). В функциональном отношении нейро-секреторные ядра, эпифиз, аденогипофиз и нейроге-мальный орган составляют центральное звено эндокринной системы, тогда как эндокринные клетки неэндокринных органов (пищеварительной системы, воздухоносных путей и легких, почек и мочеотводя-щих путей, вилочковой железы), аденогипофиззави-симые железы (щитовидная железа, кора надпочечников, половые железы) и деногипофизнезависимые железы.
31. Гипоталамус
Гипоталамус занимает базальную область межуточного мозга и окаймляет нижнюю часть III желудочка головного мозга. Полость III желудочка продолжается в воронку, стенка которой становится гипофизарной ножкой и на своем дистальном конце дает начало задней доле гипофиза, или нейрогипофизу.
В сером веществе гипоталамуса обособляются его ядра (свыше 30 пар), которые группируются в переднем, среднем (медиобазальном или тубераль-ном) и заднем отделах гипоталамуса. Некоторые из гипоталамических ядер представляют скопления ней-росекреторных клеток, а другие образованы сочетанием нейросекреторных клеток и нейронов обычного типа (преимущественно адренергическими).
В ядрах среднего гипоталамуса вырабатываются гипоталамические аденогипофизотропные гормоны, которые регулируют секрецию (и, вероятно, также продукцию) гормонов в передней и средней долях гипофиза. Аденогипофизотропные гормоны являются низкомолекулярными белками (олигопептидами), которые либо стимулируют (либерины), либо угнетают (статины) соответствующие гормонообразователь-ные функции аденогипофиза. Важнейшие ядра этой части гипоталамуса локализуются в сером бугре: ар-куатное, или инфундибулярное, ядро и вентромеди-альное ядро. Вентромедиальное ядро отличается большими размерами и оказывается основным местом выработки аденогипофизотропных гормонов, но наряду с ним эта функция присуща также аркуатному ядру. Указанные ядра образованы мелкими нейросе-креторными клетками в сочетании с адренергически-ми нейронами обычного типа. Аксоны как мелких нейросекреторных клеток медиобазального гипоталамуса, так и соседствующих с ними адренерги-ческих нейронов направляются в медиальную эминенцию, где заканчиваются на петлях первичной капиллярной сети.
Таким образом, нейросекреторные образования гипоталамуса делятся на две группы: холинергическую (крупноклеточные ядра переднего гипоталамуса) и адренергическую (мелкие нейросекреторные клетки медиобазального гипоталамуса).
Деление нейросекреторных образований гипоталамуса на пептидо-холинергические и пептидоадренер-гические отражает их принадлежность соответственно к парасимпатической или симпатической части гипоталамуса.
Связь переднего гипоталамуса с задней долей гипофиза, а медиобазального гипоталамуса – с аденогипофизом позволяет расчленить гипоталамо-гипофи-зарный комплекс на гипоталамо-нейрогипофизарную и гипоталамо-аденогипофизарную системы. Значение задней доли гипофиза состоит в том, что в ней аккумулируются и выделяются в кровь нейрогормоны, вырабатываемые крупноклеточными пептидохолинергическими ядрами переднего гипоталамуса. Следовательно, задняя доля гипофиза не является железой, а представляет собой вспомогательный нейрогемаль-ный орган гипоталамо-нейрогипофизарной системы.
Аналогичным нейрогемальным органом гипоталамо-аденогипофизарной системы оказывается медиальная эминенция, в которой накапливаются и поступают в кровь аденогипофизотропные гормоны (либерины и статины).
32. Гипофиз
В гипофизе выделяют несколько долей: аденогипо-физ, нейрогипофиз.
В аденогипофизе различают переднюю, среднюю (или промежуточную) и туберальную части. Передняя часть имеет трабекулярное строение. Трабекулы, сильно разветвляясь, сплетаются в узкопетлистую сеть. Промежутки между ними, заполнены рыхлой соединительной тканью, по которой проходят многочисленные синусоидные капилляры.
Хромофильные клетки делятся на базофильные и ацидофильные. Базофильные клетки, или базофи-лы, продуцируют гликопротеидные гормоны, и их секреторные гранулы на гистологических препаратах окрашиваются основными красками.
Среди них различают две основные разновидности: гонадотропные и тиротропные.
Одни из гонадотропных клеток вырабатывают фол-ликулостимулирующий гормон (фоллитропин), другим приписывается продукция лютеинизирующего гормона (лютропина).
Тиротропный гормон (тиротропин) – отличается неправильной или угловатой формой. При недостаточности в организме гормона щитовидной железы продукция тиротропина усиливается, а тиротропоци-ты частично трансформируются в клетки тиреоидэк-томии, которые характеризуются более крупными размерами и значительным расширением цистерн эндоплазматической сети, вследствие чего цитоплазма приобретает вид крупноячеистой пены. В этих вакуолях обнаруживаются альдегид-фуксинофильные гранулы, более крупные, чем секреторные гранулы исходных тиротропоцитов.
Для ацидофильных клеток, или ацидофилов, характерны крупные плотные гранулы, окрашивающиеся на препаратах кислыми красителями. Ацидофильные клетки также делятся на две разновидности: соматотропные, или соматотропоциты, вырабатывающие соматотропный гормон (соматотропин), и маммо-тропные, или маммотропоциты, вырабатывающие лак-тотропный гормон (пролактин).
Кортикотропные клетки в передней доле гипофиза вырабатывают адренокортикотропный гормон (АКТГ, или кортикотропин), активирующий кору надпочечников.
Туберальная часть – отдел аденогипофизарной паренхимы, прилежащей к гипофизарной ножке и соприкасающейся с нижней поверхностью медиальной эминенции гипоталамуса.
Задняя доля гипофиза (нейрогипофиз) образована нейроглией. Глиальные клетки этой доли представлены преимущественно небольшими отростчатыми или вере-теновидными клетками – питуицитами. В заднюю долю входят аксоны нейросекреторных клеток супраоптическо-го и паравентрикулярного ядер переднего гипоталамуса.
Иннервация. Гипофиз, а также гипоталамус и эпифиз получают нервные волокна от шейных ганглиев (главным образом от верхних) симпатического ствола.
Кровоснабжение. Верхние гипофизарные артерии вступают в медиальную эминенцию, где распадаются на первичную капиллярную сеть.
33. Щитовидная железа
В щитовидной железе выделяют две доли (правую и левую соответственно) и перешеек.
Снаружи она окружена плотной соединительнотканной капсулой, от которой внутрь железы отходят перегородки. Составляя строму железы, они разветвляются и делят тиреоидную паренхиму на дольки.
Тироциты – железистые клетки щитовидной железы, составляющие стенку (выстилку) фолликулов и располагающиеся в один слой на базальной мембране, ограничивающей фолликул снаружи. Форма, объем и высота тироцитов изменяются в соответствии со сдвигами функциональной активности щитовидной железы.
Интрафолликулярный коллоид становится более жидким, в нем появляются многочисленные вакуоли
Апикальная поверхность тироцита образует микроворсинки, вдающиеся в просвет фолликула. По мере усиления функциональной активности щитовидной железы количество и размеры микроворсинок возрастают.
Одновременно базальная поверхность тироцитов, при активизации ее становится складчатой, что приводит к увеличению контакта тироцитов с перикапил-лярными пространствами.
Секреторный цикл любой железистой клетки состоит из следующих фаз: поглощения исходных веществ, синтеза гормона и его выделения.
Фаза продукции. Выработка тиреоглобулина начинается в цитоплазме базальной части тироцита и завершается в полости фолликула на его апикальной поверхности. Исходные продукты, приносимые к щитовидной железе кровью и поглощаемые тироцитами через их основание, концентрируются в эндо-плазматической сети, и на рибосомах совершается синтез полипептидной цепочки – основы будущей молекулы тиреоглобулина. Получающийся продукт накапливается в цистернах эндоплазматиче-ской сети и затем перемещается в зону пластинчатого комплекса, где конденсируется тиреоглобулин (но еще не йодированный) и формируются мелкие секреторные везикулы, смещающиеся затем в верхнюю часть тироцита. Йод поглощается тироцитами из крови в форме йодида и синтезируется тироксин.
Фаза выведения. Осуществляется путем реабсорб-ции интрафолликулярного коллоида. В зависимости от степени активации щитовидной железы эндоцитоз протекает в разных формах. Выведение гормона из железы, находящейся в состоянии функционального покоя или слабого возбуждения, протекает без образования апикальных псевдоподий и без появления капель интрацеллюлярного коллоида внутри тироцитов. Оно осуществляется путем протеолиза тиреоглобули-на, совершающегося в периферическом слое интра-фолликулярного коллоида на границе с микроворсинками, и последующего микропиноцитоза продуктов этого расщепления.
Парафолликулярные клетки (кальцитониноциты), встречающиеся в тиреоидной паренхиме, резко отличаются от тироцитов отсутствием способности поглощать йод. Как упоминалось выше, они вырабатывают белковый гормон – кальцитонин (тирокальцитонин), понижающий уровень кальция в крови и являющийся антагонистом паратирина (гормона околощитовидных желез).
34. Эндокринная система
Каждая околощитовидная железа окружена тонкой соединительно-тканной капсулой. Их паренхима образована эпителиальными тяжами (трабекулами) либо скоплениями железистых клеток (паратироцитов), разделенными тонкими прослойками рыхлой соединительной ткани с многочисленными капиллярами.
Среди паратироцитов различаются главные, промежуточные и ацидофильные (оксифильные) клетки, которые, однако, следует рассматривать не как отдельные разновидности железистых клеток околощитовидных желез, а как функциональные или возрастные состояния паратироцитов.
Во время усиления секреторной активности околощитовидных желез главные клетки набухают и увеличиваются в объеме, в них гипертрофируются эндо-плазматическая сеть и пластинчатый комплекс. Выделение паратирина из железистых клеток в межклеточные щели осуществляется путем экзоцитоза. Высвобожденный гормон поступает в капилляры и выносится в общую циркуляцию.
Кровоснабжение щитовидной и паращитовидных желез осуществляется из верхних и нижних щитовидных артерий.
Надпочечники
Парные органы, образованные сочетанием двух самостоятельных желез разного происхождения и различного физиологического значения: корковой и мозговой (медуллярной). Гормоны надпочечников принимают участие в защитно-приспособительных реакциях организма, в регуляции обмена веществ и деятельности сердечно-сосудистой системы.
В надпочечниках выделяют корковый слой и мозговой слой.
Клубочковая (наружная) зона образована железистыми клетками (адренокортикоцитами) удлиненной формы, которые наслаиваются друг на друга, образуя округлые скопления, чем и обусловливается наименование данной зоны.
Средняя часть коркового вещества. Адренокорти-коциты этой зоны отличаются крупными размерами и кубической или призматической формой, их ось ориентируется вдоль эпителиального тяжа.
Пучковая зона коры надпочечников вырабатывает глюкокортикоидные гормоны – кортикостерон, кор-тизол(гидрокортизон) и кортизон.
Во внутренней сетчатой зоне эпителиальные тяжи теряют правильное расположение и, разветвляясь, образуют рыхлую сеть, в связи с чем данная зона коры получила свое название. Адренокортикоциты в этой зоне уменьшаются в объеме и становятся разнообразными по форме (кубическими, округлыми или многоугольными).
В сетчатой зоне вырабатывается андрогенный гормон (мужской половой гормон, близкий по химической природе и физиологическим свойствам к тестостерону семенника).
Мозговая часть надпочечников отделена от корковой части тонкой, местами прерывающейся, внутренней соединительно-тканной капсулой. Мозговое вещество надпочечников образовано скоплением сравнительно крупных клеток, преимущественно округлой формы, расположенных между кровеносными сосудами.
35. Пищеварительная система
Пищеварительная система человека представляет собой пищеварительную трубку с расположенными рядом с ней (железами (слюнными железами, печенью и поджелудочной железой), секрет которых участвует в процессе пищеварения.
Процессом пищеварением называются процессы химической и механической обработки пищи с последующим всасыванием продуктов ее расщепления.
Роль желудочно-кишечного тракта в организме человека очень велика: из него идет поступление веществ, обеспечивающих организм необходимой энергией и строительными материалами для восстановления постоянно разрушающихся его структур.
Слизистая оболочка. Оостоит из эпителия, собственной пластинки слизистой оболочки и мышечной пластинки слизистой оболочки. Эпителий в переднем и заднем отделах пищеварительной трубки многослойный плоский, а в среднем отделе, т. е. в желудке и кишечнике, – однослойный цилиндрический. Железы расположены либо эндоэпителиально, либо экзоэ-пителиально, либо за пределами пищеварительного канала.
В состав слизистой оболочки входит собственная пластинка, которая лежит под эпителием, отделена от него базальной мембраной и представлена рыхлой волокнистой неоформленной соединительной тканью. В ней проходят кровеносные и лимфатические сосуды, нервные элементы, скопления лимфоидной ткани.
Расположением мышечной пластинки слизистой оболочки является граница с подслизистой основой. Данная пластинка состоит из нескольких слоев, образованных гладкими мышечными клетками.
Подслизистая основа представлена рыхлой волокнистой неоформленной соединительной тканью, она как бы соединяет слизистую оболочку с подлежащими образованиями (мышечной оболочкой или костной основой). Благодаря ей слизистая оболочка имеет подвижность и может образовывать складки.
Мышечную оболочку составляет гладкомышечная ткань, в этом случае расположение мышечных волокон может быть циркулярным (внутренний слой) и продольным (наружный слой).
Брюшина состоит из соединительно-тканной основы, в которой имеются сосуды и нервные элементы, и из мезотелия, окружающего ее снаружи.
Кровоснабжение. В тонкой кишке артериальные сплетения формируются также в мышечной оболочке. Капиллярные сети образуются под эпителием слизистой оболочки, вокруг желез, крипт, желудочных ямок, внутри ворсинок, сосочков языка и в мышечных слоях. Вены также формируют сплетения подслизистой основы и слизистой оболочки.
Эфферентная иннервация всех органов пищеварения происходит из ганглиев вегетативной нервной системы, расположенных либо вне пищеварительной трубки либо в толще ее (интрамуральные парасимпатические ганглии).
Афферентная иннервация осуществляется окончаниями дендритов чувствительных нервных клеток, происходит за счет интрамуральных ганглиев, у которых окончания – дендриты от спинальных ганглиев. Чувствительные нервные окончания располагаются в мышцах, эпителии, волокнистой соединительной ткани и нервных ганглиях.
36. Ротовая полость
Слизистая оболочка, выстилающая ротовую полость состоит из многослойного плоского эпителия, слабое развитие мышечной пластинки слизистой оболочки и отсутствием в некоторых участках подсли-зистого слоя. При этом в ротовой полости имеются места, где слизистая оболочка твердо сращена с подлежащими тканями и лежит непосредственно на мышцах или на кости (в деснах и твердом нёбе). Слизистая оболочка может образовывать складки, в которых располагаются скопления лимфоидной ткани. Эти участки называют миндалинами.
Губы. В губе различают три части: кожную, переходную, или красную, и слизистую. В толще губы находится поперечно-полосатая мускулатура. Она покрыта многослойным плоским ороговевающим эпителием и снабжена сальными, потовыми железами и волосами. Эпителий этой части расположен на базальной мембране, под которой лежит рыхлая волокнистая соединительная ткань, образующая высокие сосочки, которые вдаются в эпителий.
Щеки представляют собой мышечное образование, которое покрыто снаружи кожей, а изнутри – слизистой оболочкой. В слизистой оболочке щеки различают три зоны: верхнюю (максиллярную), среднюю (промежуточную) и нижнюю (мандибулярную). При этом отличительной особенностью щек является то, что в слизистой оболочке отсутствует мышечная пластинка.
Максиллярная часть щеки имеет строение, сходное со строением слизистой части губы. Она покрыта многослойным плоским неороговевающим эпителием, сосочки собственной пластинки слизистой оболочки небольших размеров. В этих зонах имеется большое количество слюнных желез щеки.
Десны – образования, покрытые слизистой оболочкой, плотно сращенной с надкостницей верхней и нижней челюстей.
Твердое небо. Состоит из костной основы, покрытой слизистой оболочкой.
Собственная пластинка слизистой оболочки твердого нёба образована волокнистой неоформленной соединительной тканью.
Собственная пластинка слизистой оболочки имеет одну особенность: пучки коллагеновых волокон сильно переплетаются между собой и вплетаются в надкостницу, особенно хорошо это выражено в тех местах, где слизистая оболочка плотно сращена с костью.
Мягкое нёбо и язычок представлены сухожильно-мышечной основой, покрытой слизистой-оболочкой. В мягком нёбе и язычке различают ротовую (переднюю) и носовую (заднюю) поверхности.
Слизистая оболочка ротовой части мягкого неба и язычка покрыта многослойным плоским неорогове-вающим эпителием. Собственная пластинка слизистой оболочки, состоящая из рыхлой волокнистой неоформленной соединительной ткани, образует высокие узкие сосочки, глубоко вдающиеся в эпителий. Глубже имеется выраженная подслизистая основа, образованная рыхлой волокнистой неоформленной соединительной тканью с большим количеством жировых элементов и слизистыми слюнными железами. Выводные протоки этих желез открываются на ротовой поверхности мягкого нёба и язычка.
37. Пищеварительная система. Язык
Основу языка составляет поперечно-полосатая мышечная ткань, сокращение которой носит произвольный характер.
Рельеф покрывающей его слизистой оболочки различен на нижней, боковых и верхней поверхностях языка. Эпителий на нижней стороне языка многослойный, плоский, неороговевающий, небольшой толщины. Слизистая оболочка верхней и боковых поверхностей языка неподвижно сращена с его мышечным телом. Она содержит особые образования – сосочки.
На поверхности языка имеется четыре вида сосочков: нитевидные, грибовидные, окруженные валом и листовидные.
Слизистая оболочка корня языка не имеет сосочков. Возвышения эпителия образуются за счет того, что в собственной пластинке слизистой оболочки имеются скопления лимфоидной ткани, достигающие иногда 0,5 см в диаметре. Между этими скоплениями эпителий образует углубления – крипты. В крипты впадают протоки многочисленных слизистых желез. Совокупность скоплений лимфоидной ткани в корне языка называется язычной миндалиной.
Кровоснабжение языка осуществляется язычными артериями.
Мышцы языка иннервируются ветвями подъязычного нерва и барабанной струны. Чувствительная иннервация передних 2/3 языка осуществ-ляется ветвями тройничного нерва, задней 1/3 – ветвями языкогло-точного нерва.
Слюнные железы. В полости рта имеются отверстия выводных протоков трех пар больших слюнных желез: околоушных, подчелюстных и подъязычных.
Все слюнные железы представляют собой сложные альвеолярные или альвеолярно-труб-чатые железы. Они включают в себя секреторные концы отделов и протоков, выводящие секрет.
Секреторные отделы по строению и характеру выделяемого секрета бывают трех типов: боковые (серозные), слизистые и смешанные (т. е. белково-сли-зистые).
Зубы являются основной частью жевательного аппарата. Существует несколько видов зубов: вначале образуются выпадающие (молочные) зубы, а затем постоянные. В лунках челюстных костей зубы укрепляются плотной соединительной тканью: периодон-том, который в области шейки зуба образует циркулярную зубную связку. Коллагеновые волокна зубной связки имеют преимущественно радиальное направление, при этом с одной стороны они проникают в цемент корня зуба, а с другой – в альвеолярную кость. Периодонт выполняет не только механическую, но и трофическую функцию, так как в нем проходят кровеносные сосуды, питающие корень зуба.
Строение зуба. Он включает в себя две части: твердую и мягкую. В твердой части зуба выделяют эмаль, дентин и цемент, мягкие части зуба представлены так называемой пульпой.
Дентин образует большую часть коронки, шейки и корня зубов. Цемент покрывает корень зуба и шейку, где в виде тонкого слоя частично может заходить на эмаль. По направлению к верхушке корня цемент утолщается. По химическому составу цемент приближается к кости.
38. Пищеварительная система. Глотка
Это место пересечения дыхательного и пищеварительного путей. Соответственно функциональным условиям в глотке различают три отдела, которые имеют различное строение: носовой, ротовой и гортанный. Все они отличаются строением слизистой оболочки, которая представлена различными видами эпителиев.
Слизистая оболочка носового отдела глотки покрыта многорядным мерцательным эпителием, содержит смешанные железы (респираторный тип слизистой оболочки).
Слизистая оболочка ротового и гортанного отделов выстлана многослойным плоским эпителием, располагающимся на собственной пластинке слизистой оболочки, в которой имеется хорошо выраженный слой эластических волокон.
Пищевод представляет собой полую трубку, которая состоит из слизистой оболочки, подслизистой основы, мышечной и адвентициальной оболочек.
Слизистая оболочка пищевода состоит из эпителия, собственной и мышечной пластинок. Эпителий слизистой оболочки многослойный, плоский, неорогове-вающий.
Собственная пластинка слизистой оболочки пищевода представляет собой слой рыхлой волокнистой неоформленной соединительной ткани, вдающейся в виде сосочков в эпителий.
Мышечная пластинка слизистой оболочки пищевода состоит из расположенных вдоль него пучков гладких мышечных клеток, окруженных сетью эластических волокон.
Подслизистая основа пищевода, образованная рыхлой волокнистой неоформленной соединительной тканью, обеспечивает большую подвижность слизистой оболочки по отношению к мышечной оболочке. Вместе со слизистой она образует многочисленные продольные складки, которые расправляются во время проглатывания пищи. В подслизистой основе находятся собственные железы пищевода.
Мышечная оболочка пищевода состоит из внутреннего циркулярного и наружного продольного слоев, разделенных прослойкой рыхлой волокнистой неоформленной соединительной ткани. При этом в верхнем отделе мышцы пищевода относятся к поперечнополосатой ткани, в среднем – к поперечно-полосатой ткани и гладкой мускулатуре, а в нижнем – только к гладкой.
Адвентициальная оболочка пищевода состоит из рыхлой волокнистой неоформленной соединительной ткани, которая, с одной стороны, связана с прослойками соединительной ткани в мышечной оболочке, а с другой – с окружающей пищевод соединительной тканью средостения.
Кровоснабжение пищевода производится из артерии, входящей в пищевод, при этом образуются сплетения в подслизистой основе (крупнопетлистые и мелкопетлистые), из которых кровь поступает в крупнопетлистое сплетение собственной пластинки слизистой оболочки.
Иннервация. Интрамуральный нервный аппарат образован связанными между собой тремя сплетениями: адвентициальным, субадвентициальным, межмышечным.
39. Пищеварительная система
Желудок
Секреторная. Функция заключается в выработке железами желудочного сока.
Механическая функция желудка состоит в перемешивании пищи с желудочным соком и проталкивании переработанной пищи в двенадцатиперстную кишку.
Эндокринная функция желудка заключается в выработке ряда биологически активных веществ: гастрина, гистамина, серотонина, мотилина, энтероглюкагона.
Строение. Стенка желудка состоит из слизистой оболочки, подслизистой основы, мышечной и серозной оболочек.
Слизистая оболочка желудка имеет неровную поверхность из-за наличия в ней трех видов образований: складок, полей и ямок.
Эпителий, выстилающий поверхность слизистой оболочки желудка и ямок, однослойный цилиндрический.
Собственная пластинка слизистой оболочки желудка представлена рыхлой волокнистой неоформленной соединительной тканью.
Она состоит из трех слоев, образованных гладкой мышечной тканью: внутреннего и наружного циркулярных и среднего продольного. Каждый из этих слоев состоит из пучков гладких мышечных клеток.
Различают три вида желудочных желез: собственные желудочные, пилорические и кардиальные.
Собственные железы желудка содержат несколько видов железистых клеток: главные, париетальные (об-кладочные), слизистые, шеечные и эндокринные (ар-гирофильные).
Главные клетки секретируют пепсиноген: профермент, который в присутствии соляной кислоты превращается в активную форму – пепсин. Основная роль париетальных клеток собственных желез желудка состоит в выработке хлоридов, из которых образуется соляная кислота.
Подслизистая основа желудка состоит из рыхлой волокнистой неоформленной соединительной ткани, содержащей большое количество эластических волокон. В этом слое находятся артериальные и венозные сплетения, сеть лимфатических сосудов и подслизи-стое нервное сплетение.
Мышечная оболочка желудка характеризуется слабым развитием в области его дна, хорошей выраженностью в теле и достижением наибольшего развития в привратнике. В мышечной оболочке желудка различают три слоя, образованные гладкой мышечной тканью.
Серозная оболочка желудка образует наружную часть его стенки. Ее основу составляет рыхлая волокнистая неоформленная соединительная ткань, прилегающая к мышечной оболочке желудка. С поверхности эта соединительно-тканная прослойка покрыта однослойным плоским эпителием – мезотелием.
Артерии, питающие стенку желудка, проходят через серозную и мышечную оболочки, отдавая им соответствующие ветви, а далее переходят в мощное сплетение в подслизистой основе. К основным источникам питания относят правую и левую желудочковые артерии. От желудка кровь оттекает в портальную вену.
В стенке желудка располагаются три нервных сплетения: межмышечное, подслизистое и субсерозное.
40. Тонкая кишка
В тонкой кишке подвергаются химической обработке все виды питательных веществ: белки, жиры и углеводы. В переваривании белков участвуют ферменты энтерокиназа, киназоген и трипсин, расщепляющие простые белки, эрепсин (смесь пептидаз), расщепляющий пептиды до аминокислот, и нуклеаза, которая переваривает сложные белки (нуклеопротеиды). Переваривание углеводов происходит за счет амилазы, мальтозы, сахарозы, лактозы и фосфатазы, а жиров – фермента липазы.
В тонкой кишке происходит также процесс всасывания продуктов расщепления белков, жиров и углеводов в кровеносные и лимфатические сосуды. Также тонкий кишечник выполняет механическую функцию: проталкивает химус в каудальном направлении.
Строение. Стенка тонкой кишки состоит из слизистой оболочки, подслизистой основы, мышечной и серозной оболочек.
С поверхности каждая кишечная ворсинка выстлана однослойным цилиндрическим эпителием. В эпителии различают три вида клеток: каемчатые, бокаловидные и эндокринные (аргирофильные).
Энтероциты с исчерченной каемкой составляют основную массу эпителиального пласта, покрывающего ворсинку. Они характеризуются выраженной полярностью строения, что отражает их функциональную специализацию: обеспечение резорбции и транспорта веществ, поступающих с пищей.
Бокаловидные кишечные – по строению это типичные слизистые клетки. В них наблюдаются циклические изменения, связанные с накоплением и последующим выделением слизи.
Эпителиальная выстилка кишечных крипт содержит следующие виды клеток: каемчатые, безкаем-чатые кишечные клетки, бокаловидные, эндокринные (аргирофильные) и кишечные клетки с ацидофильной зернистостью (клетки Панета).
Собственная пластинка слизистой оболочки тонкой кишки с основном состоит из большого количества ретикулярных волокон. Они образуют густую сеть по всей собственной пластинке и, подходя к эпителию, участвуют в образовании базальной мембраны.
В подслизистой основе находятся сосуды и нервные сплетения.
Мышечная оболочка представлена двумя слоями гладкомышечной ткани: внутренним (циркулярным) и наружным (продольным).
Серозная оболочка покрывает кишку со всех сторон, за исключением двенадцатиперстной кишки. Лимфатические сосуды тонкой кишки представлены очень широко разветвленной сетью. В каждой кишечной ворсинке есть центрально расположенный, слепо оканчивающийся на ее вершине лимфатический капилляр.
Иннервация. Тонкая кишка иннервируется симпатическими и парасимпатическими нервами.
Афферентная иннервация осуществляется чувствительным мышечно-кишечным сплетением, образованным чувствительными нервными волокнами спиналь-ных ганглиев и их рецепторными окончаниями.
Эфферентная парасимпатическая иннервация осуществляется за счет мышечно-кишечного и подслизи-стого нервных сплетений.
41. Пищеварительная система. Толстая кишка
В толстой кишке происходит всасывание воды из химуса и формирование каловых масс.
В толстом кишечнике выделяют ободочную кишку и прямую.
Ободочная кишка. Стенка ободочной кишки (так же, как и всего желудочно-кишечного тракта,) состоит из слизистой оболочки, подслизистой основы, мышечной и серозной оболочек.
В слизистой оболочке имеется большое количество складок и крипт, значительно увеличивающих ее поверхность, но отсутствуют ворсинки.
Складки образуются на внутренней поверхности кишки из слизистой оболочки и подслизистой основы. Крипты в ободочной кишке развиты лучше, чем в тонкой. При этом эпителий – однослойный призматический, в его состав входят клетки кишечного эпителия с исчерченной каемкой, бокаловидные и кишечные клетки без каемки.
Собственная пластинка слизистой оболочки состоит из рыхлой волокнистой неоформленной соединительной ткани.
Мышечная пластинка слизистой оболочки состоит из двух полосок. Внутренняя ее полоска более плотная, образована преимущественно циркулярно расположенными пучками гладких мышечных клеток. Наружная полоска представлена пучками гладких мышечных клеток, ориентированных частично продольно, частично косо по отношению к оси кишки.
Подслизистая основа состоит из рыхлой волокнистой неоформленной соединительной ткани, в которой много жировых клеток.
Червеобразный отросток. Червеобразный отросток является рудиментарным образованием толстой кишки, в нем имеются большие скопления лимфоидной ткани. Слизистая оболочка червеобразного отростка имеет крипты, которые расположены радиально по отношению к его просвету.
Эпителий слизистой оболочки цилиндрический, каемчатый, с небольшим количеством бокаловидных клеток.
Собственная пластинка слизистой оболочки состоит из рыхлой волокнистой неоформленной соединительной ткани, которая без резкой границы (вследствие слабого развития мышечной пластинки слизистой) переходит в подслизистую основу.
В подслизистой основе червеобразного отростка, образованной рыхлой волокнистой неоформленной соединительной тканью, залегают кровеносные сосуды и нервное подслизистое сплетение.
Мышечная оболочка также образована двумя слоями.
Прямая кишка. Прямая кишка является продолжением ободочной кишки.
В анальной части кишки различают три зоны: столбчатую, промежуточную и кожную. В столбчатой зоне продольные складки образуют заднепроходные столбы.
В кожной зоне, окружающей анальное отверстие, к сальным железам присоединяются волосы.
42. Пищеварительная сисема. Печень
Печень – одна из крупных желез пищеварительного тракта, выполяющая многочисленные функции. В ней происходит:
1) обезвреживание различных продуктов обмена веществ;
2) разрушение различных биологически активных веществ, гормонов;
3) печень принимает активное участие в различных защитных реакциях организма;
4) она принимает участие в образовании гликогена (основного источника глюкозы);
5) также в печени происходит образование различных белков;
6) она является одним из органов кроветворения;
7) в ней накапливаются витамины;
 она принимает активное участие в образовании желчи.
она принимает активное участие в образовании желчи.
Строение. Печень – это непарный орган, находящийся в брюшной полости, покрытый брюшиной со всех сторон.
Основной структурно-функциональной единицей печени является печеночная долька. Она представляет собой шестигранную призму из печеночных клеток (гепатоцитов, собранных в виде балок). Каждая долька покрыта соединительно-тканной оболочкой, в которой проходят желчные протоки и кровеносные сосуды. От периферии дольки к ее центру кровь проходит по кровеносным сосудам, очищаясь, и по центральной вене печеночной дольки попадает в собирательные вены, далее в печеночные вены и в нижнюю полую вену.
Между рядами гепатоцитов, проходят желчные капилляры. Эти капилляры не имеют собственной стенки.
Их стенка образована соприкасающимися поверхностями гепатоцитов, на которых имеются небольшие углубления, совпадающие друг с другом и вместе образующие просвет желчного капилляра.
Желчевыводящие пути представляют собой систему желчных сосудов, по которым происходит транспорт желчи из печени в просвет двенадцатиперстной кишки.
Желчный пузырь – это полый орган с тонкой стенкой. Стенка состоит из трех оболочек: слизистой, мышечной и адвентициальной.
Слизистая оболочка желчного пузыря образует складки, анастомозирующие друг с другом, а также крипты или синусы в виде карманов.
В области шейки пузыря в ней находятся альвео-лярно-трубчатые железы, выделяющие слизь. Эпителий слизистой оболочки обладает способностью всасывать воду и некоторые другие ве-щества из желчи, заполняющей полость пузыря. В связи с этим пузырная желчь всегда более густой консистенции и более темного цвета, чем желчь, выходящая непосредственно из печени.
Мышечная оболочка желчного пузыря состоит из гладких мышечных клеток (, которые особенно хорошо развиты в области шейки пузыря.
Адвентициальная оболочка желчного пузыря состоит из плотной волокнистой соединительной ткани.
Иннервация. В капсуле печени находится вегетативное нервное сплетение, ветви которого, сопровождая кровеносные сосуды, продолжаются в меж-дольковую соединительную ткань.
43. Пищеварительная система
Поджелудочная железа
Поджелудочная железа – это орган пищеварительной системы, в составе которого находятся экзокрин-ная и эндокринная части. Экзокринная часть отвечает за выработку панкреатического сока, в котором содержатся пищеварительные ферменты (трипсин, липаза, амилаза и др.), поступающего по выводным протокам в двенадцатиперстную кишку, где его ферменты участвуют в расщеплении белков, жиров и углеводов до конечных продуктов. В эндокринной части синтезируется ряд гормонов (инсулин, глюкагон, соматостатин, панкреатический полипептид), принимающих участие в регуляции углеводного, белкового и жирового обмена в тканях.
Строение. Поджелудочная железа – непарный орган брюшной полости, на поверхности покрытый соединительно-тканной капсулой, срастающейся с висцеральным листком брюшины. Ее паренхима разделена на дольки, между которыми проходят соединительнотканные тяжи. В них расположены кровеносные сосуды, нервы, интрамуральные нервные ганглии, пластинчатые тельца (тельца Фатера—Пачини) и выводные протоки.
Ацинозные клетки выполняют секреторную функцию, синтезируя пищеварительные ферменты панкреатического сока. Они имеют форму конуса с суженной верхушкой и широким основанием, лежащим на базальной мембране ацинуса.
Секреция гормонов происходит циклически. Фазы секреции те же, что и у других желез. Однако выделение секрета по мерокриновому типу происходит в зависимости от физиологических потребностей организма в пищеварительных ферментах, этот цикл может сократиться или, наоборот, увеличиться.
Выделившийся секрет проходит по протокам (вставочным, межацинарным, внутридольковым), которые, объединяясь, впадают в Вирсунгов проток.
Стенки этих протоков выстланы однослойным кубическим эпителием. Их цитолемма образует внутренние складки и микроворсинки.
Островки состоят из эндокринных инсулярных клеток – инсулоцитов. Между ними находятся кровеносные капилляры фенестрированного типа. Капилляры окружены перикапиллярным пространством. Гормоны, выделяемые инсулярными клетками, сначала попадают в это пространство, а затем через стенку капилляров в кровь.
Среди инсулярных клеток различают пять основных видов: В-клетки (базофильные), А-клетки (ацидофильные), D-клетки (дендритические), 01-клетки (ар-гирофильные) и РР-клетки.
В-клетки составляют основную массу клеток островков
D-клетки секретируют гормон соматостатин.
РР-клетки вырабатывают панкреатический полипептид, стимулирующий выделение желудочного и панкреатического сока.
Кровоснабжение поджелудочной железы происходит из ветвей чревного ствола. Венозная кровь оттекает от поджелудочной железы в воротную вену.
Иннервация. Эфферентная иннервация поджелудочной железы осуществляется блуждающим и симпатическим нервами.
44. Дыхательная система
Основной функцией дыхательной системы является внешнее дыхание, т. е. поглощение из вдыхаемого воздуха кислорода и снабжение им крови, а также удаление из организма углекислого газа (газообмен осуществляется легкими, их ацинусами). Внутреннее, тканевое дыхание происходит в виде окислительных процессов в клетках органов при участии крови. Функций: терморегуляцию и увлажнение вдыхаемого воздуха, очищение его от пыли и микроорганизмов, депонирование крови в обильно развитой сосудистой системе, участие в поддержании свертываемости крови благодаря выработке тромбопластина и его антагониста (гепарина), участие в синтезе некоторых гормонов и в водносолевом, липидном обмене веществ, а также в голосообразовании, обонянии и иммунологической защите.
Воздухоносные пути
К воздухоносным путям дыхательной системы относят носовую полость, носоглотку, гортань, трахею и бронхи. При продвижении воздуха происходит его очищение, увлажнение, приближение температуры вдыхаемого воздуха к температуре тела, рецепция газовых, температурных и механических раздражителей, а также регуляция объема вдыхаемого воздуха.
Полость носа
Она делится на преддверие и собственно носовую полость, состоящую из дыхательной и обонятельной областей.
Преддверие образовано полостью, находится под хрящевой частью носа, покрыто многослойным плоским эпителием.
Под эпителием в соединительно-тканном слое имеются сальные железы и корни щетинковых волос. Щетинковые волосы выполняют очень важную функцию: они задерживают пылевые частицы из вдыхаемого воздуха в носовой полости.
Эпителий состоит из несколько видов клеток: реснитчатых, микроворсинчатых, базальных и бокаловидных.
Собственная пластинка слизистой оболочки образована рыхлой волокнистой неоформленной соединительной тканью, содержащей большое количество эластических волокон. В ней залегают концевые отделы слизистых желез, выводные протоки которых открываются на поверхности эпителия. Секрет этих желез, как и секрет бокаловидных клеток, увлажняет слизистую оболочку.
Слизистая оболочка носовой полости хорошо кро-воснабжается, что способствует согреванию вдыхаемого воздуха в холодное время года.
Лимфатические сосуды связаны с субарахноидаль-ным пространством и периваскулярными влагалищами различных частей мозга, а также с лимфатическими со-судами больших слюнных желез.
Слизистая оболочка носовой полости имеет обильную иннервацию, многочисленные свободные и инкапсулированные нервные окончания
В области верхней носовой раковины слизистая оболочка покрыта особым обонятельным эпителием, содержащим рецепторные (обонятельные) клетки.
Слизистая оболочка околоносовых пазух, в том числе лобных и верхнечелюстных, такая же слизистая оболочка дыхательной части носовой полости, с той лишь разницей, что собственная соединительно-тканная пластинка в них значительно тоньше.
45. Дыхательная система
Гортань, трахея
Гортань участвует не только воздухопроведении, но и в звукопроизведении. Гортань имеет три оболочки: слизистую, фиброзно-хрящевую и адвентициальную.
Слизистая оболочка гортани выстлана многорядным реснитчатым эпителием. Собственная пластинка слизистой оболочки, образованная рыхлой волокнистой неоформленной соединительной тканью, содержит многочисленные эластические волокна, не имеющие определенной ориентировки.
В глубоких слоях слизистой оболочки эластические волокна постепенно переходят в надхрящницу, а в средней части гортани проникают между поперечнополосатыми мышцами голосовых связок.
В средней части гортани имеются складки слизистой оболочки, образующие так называемые истинные и ложные голосовые связки. Складки покрывает многослойный плоский эпителий. В слизистой оболочке залегают смешанные железы. Благодаря сокращению поперечнополосатых мышц, заложенныхв толще голосовых складок, происходит изменение величины щели между ними, что влияет на высоту звука, производимого воздухом, проходящим через гортань.
Фиброзно-хрящевая оболочка состоит из гиалиновых и эластических хрящей, окруженных плотной волокнистой соединительной тканью. Эта оболочка является своеобразным каркасом гортани.
Адвентициальная оболочка состоит из волокнистой соединительной ткани.
Гортань отделена от глотки надгортанником, основу которого составляет эластический хрящ. В области надгортанника происходит переход слизистой оболочки глотки в слизистую оболочку гортани. На обеих поверхностях надгортанника слизистая оболочка покрыта многослойным плоским эпителием. Трахея
Это воздухопроводящий орган дыхательной системы, представляющий собой полую трубку, состоящую из слизистой оболочки, подслизистой основы, волокнисто-хрящевой и адвентициальной оболочек.
Слизистая оболочка. Она выстлана многорядным призматическим реснитчатым эпителием, в котором различают реснитчатые, бокаловидные, эндокринные и базальные клетки.
Волокнисто-хрящевая оболочка трахеи состоит из 16–20 гиалиновых хрящевых колец, не замкнутых на задней стенке трахеи. Свободные концы этих хрящей соединены пучками гладких мышечных клеток, прикрепляющихся к наружной поверхности хряща.
Адвентициальная оболочка трахеи состоит из рыхлой волокнистой неоформленной соединительной ткани, которая соединяет этот орган с прилежащими частями средостения.
Кровеносные сосуды трахеи, образуют в ее слизистой оболочке несколько параллельно расположенных сплетений, а под эпителием – густую капиллярную сеть. Лимфатические сосуды также формируют сплетения, из которых поверхностное находится непосредственно под сетью кровеносных капилляров.
Нервы, подходящие к трахее, содержат спинномозговые и вегетативные волокна и образуют два сплетения, ветви которых заканчиваются в ее слизистой оболочке нервными окончаниями. Мышцы задней стенки трахеи иннервируются из ганглиев вегетативной нервной системы.
46. Органы дыхания. Легкие
Строение. Легкое состоит из разветвлений бронхов, входящих в состав воздухоносных путей (бронхиального дерева), и системы легочных пузырьков (альвеол), выполняющих роль респираторных отделов дыхательной системы.
Строение бронхов Внутренняя оболочка бронхов – слизистая – выстлана, подобно трахее, реснитчатым эпителием, толщина которого постепенно уменьшается за счет изменения формы клеток от высоких призматических до низких кубических.
Секреторные клетки характеризуются куполообразной верхушкой, лишенной ресничек и микроворсинок и заполненной секреторными гранулами. Эти клетки вырабатывают ферменты, расщепляющие сурфак-тант, покрывающий респираторные отделы.
Безреснитчатые клетки имеют призматическую форму.
Каемчатые клетки отличаются овоидной формой и наличием на апикальной поверхности коротких тупых микроворсинок.
Собственная пластинка слизистой оболочки бронхов богата продольно направленными эластическими волокнами, которые обеспечивают растяжение бронхов при вдохе и возвращение их в исходное положение при выдохе.
В подслизистой соединительной основе залегают концевые отделы смешанных слизисто-белковых желез. Они располагаются группами, особенно в местах, которые лишены хряща, а выводные протоки проникают в слизистую оболочку и открываются на поверхности эпителия. Их секрет увлажняет слизистую оболочку и способствует прилипанию, обволакиванию пылевых и других частиц, которые впоследствии выделяются наружу. Слизь обладает бактериостатическими и бактерицидными свойствами. Фиброзно-хрящевая оболочка характеризуется постепенной сменой незамкнутых хрящевых колец у главных бронхов на хрящевые пластинки и островки хрящевой ткани.
Наружная адвентициальная оболочка состоит из волокнистой соединительной ткани, переходящей в междолевую и междольковую соединительную ткань паренхимы легкого. Среди соединительно-тканных клеток есть тканевые базофилы, принимающие участие в регуляции состава межклеточного вещества и свертываемости крови.
Респираторный отдел. Структурно-функциональной единицей респираторного отдела легкого является ацинус. состоит из систем альвеол, расположенных в стенке респираторной бронхиолы, альвеолярных ходов и мешочков, которые осуществляют газообмен между кровью и воздухом альвеол. Ацинусы отделены друг от друга тонкими соединительно-тканными прослойками; которое образуют легочную дольку. Респираторные бронхиолы выстланы однослойным кубическим эпителием. Мышечная пластинка истончается и распадается.
Альвеолы имеют вид открытого пузырька. Внутренняя поверхность выстлана двумя основными видами клеток: респираторными альвеолярными клетками и большими альвеолярными клетками
В стенке альвеол также обнаруживаются щеточные клетки и макрофаги, содержащие захваченные инородные частицы, избыток сурфактанта.
47. Дыхательная система. Плевра
Плевра – серозная оболочка, выстилающая грудную полость и покрывающая легкие. Между оболочками имеется плевральная полость, в которой содержится плевральная жидкость, смягчающая трение легких в процессе дыхания.
Легкие снаружи покрыты плеврой, называемой легочной или висцеральной. Висцеральная плевра плотно срастается с легкими, эластические и коллагено-вые волокна ее переходят в интерстициальную ткань, поэтому изолировать плевру, не травмируя легкие, трудно. В висцеральной плевре встречаются гладкие мышечные клетки. В париетальной плевре, выстилающей наружную стенку плевральной полости, эластических элементов меньше, гладкие мышечные клетки встречаются редко.
Кровоснабжение в легком осуществляется по двум системам сосудов. С одной стороны, легкие получают артериальную кровь из большого круга кровообращения по бронхиальным артериям, а с другой – в них поступает венозная кровь для газового обмена из легочных артерий, т. е. из малого круга кровообращения. Ветви легочной артерии, сопровождая бронхиальное дерево, доходят до основания альвеол, где они образуют капиллярную сеть альвеол. Через альвеолярные капилляры, диаметр которых колеблется в пределах 5–7 мкм, эритроциты проходят в один ряд, что создает оптимальное условие для осуществления газового обмена между гемоглобином эритроцитов и альвеолярным воздухом. Альвеолярные капилляры собираются в посткапиллярные венулы, которые, сливаясь, образуют легочные вены.
Бронхиальные артерии отходят непосредственно от аорты, питают бронхи и легочную паренхиму артериальной кровью. Проникая в стенку бронхов, они разветвляются и образуют артериальные сплетения в их подслизистой основе и слизистой оболочке. В слизистой оболочке бронхов происходит сообщение сосудов большого и малого круга путем анасто-мозирования разветвлений бронхиальных и легочных артерий.
Лимфатическая система легкого состоит из поверхностной и глубокой сетей лимфатических капилляров и сосудов. Поверхностная сеть располагается в висцеральной плевре. Глубокая сеть находится внутри легочных долек, в междольковых перегородках, залегая вокруг кровеносных сосудов и бронхов легкого.
Иннервация осуществляется симпатическими и парасимпатическими нервами и небольшим количеством волокон, идущих от спинномозговых нервов. Симпатические нервы проводят импульсы, вызывающие расширение бронхов и сужение кровеносных сосудов, парасимпатические – импульсы, обусловливающие, наоборот, сужение бронхов и расширение кровеносных сосудов. Разветвления этих нервов образуют в соединительно-тканных прослойках легкого нервное сплетение, расположенное по ходу бронхиального дерева и кровеносных сосудов. В нервных сплетениях легкого встречаются крупные и мелкие ганглии, от которых отходят нервные ветви, иннерви-рующие, по всей вероятности, гладкую мышечную ткань бронхов. Нервные окончания выявлены по ходу альвеолярных ходов и альвеол.
48. Кожа (строение эпидермиса)
Функция кожи: защита подлежащих частей организма от повреждений.
Строение. Кожа состоит из двух частей: эпителиальной и соединительно-тканной.
Эпителий кожи называется надкожицей, или эпидермисом, а соединительно-тканная основа – дермой, или собственно кожей. Соединение кожи с подлежащими частями организма происходит посредством слоя жировой ткани: подкожной клетчаткой, или гиподермой. Эпидермис состоит из плоского ороговеваю-щего эпителия. Наиболее толстый эпидермис расположен на ладонях и подошвах, он состоящий из множества слоев клеток. Данные клетки состоят из пяти основных слоев, к числу которых относятся: ба-зальный, шиповатый, зернистый, блестящий и роговой. Непосредственно на базальной мембране, отграничивающей эпителий от дермы, лежат клетки, составляющие базальный слой. Среди них различают базальные эпидермоциты, меланоциты (пигментные клетки),
Кроме эпидермоцитов, в шиповатом слое наблюдаются белые отростчатые клетки (клетки Лангерганса). Они лишены тонофибрилл и не образуют десмосом.
В их цитоплазме много лизосом и встречаются гранулы меланина, захваченные из отростков меланоцитов. Особенностью базального и глубоких уровней шиповатого слоя эпидермиса является способность эпидермоцитов к размножению путем митотического деления. Зернистый слой представляет собой 3–4 слоя сравнительно плоских клеток. Их цитоплазма содержит рибосомы, митохондрии, лизосомы и их разновидность (кератиносомы (в виде слоистых телец)), а также пучки фрагментированных тонофибрилл и лежащие радом с ними крупные гранулы кера-тогиалина. Следующий слой (блестящий) также состоит из 3–4 слоев плоских клеток, в которых ядра вследствие их гибели перестают окрашиваться, а цитоплазма диффузно пропитана белковым веществом – элеидином, который, с одной стороны, не окрашивается красителями, а с другой – хорошо преломляет свет. Из-за этого структура клеток в блестящем слое границы является незаметной, а весь слой по виду представляет блестящую полосу.
Роговой слой представлен множеством роговых чешуек. Чешуйки содержат роговое вещество кератин и пузырьки воздуха. Кератин – это богатый серой белок, характеризующийся устойчивостью к различным химическим агентам: кислотам, щелочам и др. Внутри клеток располагаются кератиновые фибриллы. Роговые чешуйки, находящиеся на поверхности постоянно отпадают, слущиваются и заменяются новыми, происходящими из слоев, лежащих ниже. При слущива-нии большое значение имеют кератиносомы, которые выходят из клеток, концентрируясь в межклеточных пространствах. в процессе ороговения эпидермиса кожи участвует ряд компонентов клеток: тонофибрил-лы, кератогиалин, кератиносомы, десмосомы. По сравнению с кожей ладоней и подошв в других участках кожи эпидермис значительно тоньше. Блестящий слой в нем отсутствует, а роговой представлен лишь 2–3 рядами ороговевших клеток (чешуек). По всей вероятности, ороговение в этом случае протекает по сокращенному циклу. Следовательно, большая часть кожи имеет эпидермис, который состоит из трех основных слоев: росткового, зернистого и рогового.
49. Кожа (строение дермы)
Дерма состоит из двух слоев (сосочкового и сетчатого), не имеющих между собой четкой границы. Со-сочковый слой располагается непосредственно под эпидермисом и состоит из рыхлой волокнистой неоформленной соединительной ткани, отвечающей за трофическую функцию. Соединительная ткань сосоч-кового слоя состоит из тонких коллагеновых, эластических и ретикулярных волокон, клеток с наиболее часто встречающимися фибробластами, макрофагами, тканевыми базофилами (тучные клетки) Сетчатый слой состоит из плотной неоформленной соединительной ткани с наличием мощных пучков коллагено-вых волокон, проходящих либо параллельно поверхности кожи, либо косо, и сети эластических волокон. Вместе они образуют сеть, где посредством функциональной нагрузки на кожу определяется ее строение. Клеточные элементы сетчатого слоя представлены главным образом фибробластами. В большинстве участков кожи человека в ее сетчатом слое находятся потовые и сальные железы, а также корни волос. Строение сетчатого слоя полностью соответствует выполняемой им функции: обеспечению прочности всей кожи.
Пучки коллагеновых волокон из сетчатого слоя дермы переходят в слой подкожной клетчатки. Между ними остаются значительные промежутки, заполненные дольками жировой ткани. Подкожная клетчатка смягчает действие на кожу различных механических факторов, поэтому она особенно хорошо развита в таких местах, как подушечки пальцев, ступни и т. д.
Кожный пигмент относится к группе меланинов. Образуется меланин при окислении аминокислоты тирозина под влиянием фермента тирозиназы и ДОФА-оксидазы. В дерме кожи пигмент располагается в цитоплазме дермальных меланоцитов (клеток отростчатой формы), однако в отличие от меланоци-тов эпидермиса они не дают положительной ДОФА-реакции. Из-за этого пигментные клетки дермы содержат, но не синтезируют пигмент. Из глубокой кожной артериальной сети начинаются артерии, которые по прохождению сетчатого слоя дермы в основании сосочкового слоя распадаются на артериолы, участвующие в образовании подсосочковой (поверхностной) артериальной сети, от которой разветвляются веточки, распадающиеся в сосочках на капилляры. Короткие артериальные веточки, отходящие от подсо-сочковой сети, снабжают кровью группы сосочков.
Капилляры сосочкового слоя, сальных желез и корней волос собираются в вены, впадающие в подсосоч-ковые венозные сплетения. В это же сплетение от жировых долек и потовых желез направляется кровь. Соединение кожного сплетения с фасциальным происходит посредством венозного сплетения, от которого отходят более крупные венозные стволы. В коже широко распространены артериоловенулярные анастомозы (гломусы), особенно многочисленные на кончиках пальцев рук и ног и в области ногтевого ложа. Лимфатические сосуды кожи образуют два сплетения: поверхностное, лежащее несколько ниже под-сосочковых венозных сплетений, и глубокое, расположенное в подкожной жировой клетчатке.
Иннервация кожи происходит посредством ветвей цереброспинальных нервов.
50. Волосы
Строение. Волосы являются эпителиальными придатками кожи. В волосе различают две части: стержень и корень. Стержень волоса находится над поверхностью кожи. Корень волоса скрыт в толще кожи и доходит до подкожной жировой клетчатки. Стержень волоса образован корковым веществом и кутикулой. Корень длинных и щетинистых волос состоит из коркового вещества, мозгового вещества и кутикулы, в пушковых волосах – только из коркового вещества и кутикулы.
Корень волоса располагается в волосяном мешке, или фолликуле, стенка которого состоит из внутреннего и наружного эпителиальных влагалищ и соединительно-тканной волосяной сумки.
Корень волоса заканчивается расширением (волосяной луковицей). С ней сливаются оба эпителиальных влагалища. Снизу в волосяную луковицу вдается соединительная ткань с капиллярами в виде волосяного сосочка. В месте перехода корня волоса в стержень эпидермис кожи образует небольшое углубление: волосяную воронку. Ростковый слой эпидермиса воронки переходит в наружное эпителиальное влагалище. Внутреннее эпителиальное влагалище на этом уровне заканчивается. В волосяную воронку открывается проток одной или нескольких сальных желез. Ниже сальных желез в косом направлении проходит мышца, поднимающая волос.
Волосяная луковица состоит из эпителиальных клеток, способных к размножению. Размножаясь, клетки волосяной луковицы передвигаются в мозговое и корковое вещество корня волоса, его кутикулу и во внутреннее эпителиальное влагалище. Питание волосяной луковицы осуществляется сосудами, расположенными в волосяном сосочке.
Кутикула волоса непосредственно прилежит к корковому веществу. Ближе к волосяной луковице она представлена цилиндрическими клетками, лежащими перпендикулярно к поверхности коркового вещества. В более поверхностных участках корня волоса эти клетки приобретают наклонное положение и превращаются в роговые чешуйки, налегающие друг на друга в виде черепицы.
Внутреннее корневое влагалище является производным волосяной луковицы. В нижних отделах корня волоса оно переходит в вещество волосяной луковицы, а в верхних отделах на уровне протоков сальных желез исчезает. В нижних отделах во внутреннем корневом влагалище различают три слоя: кутикулу, гра-нулосодержаший эпителиальный слой (слой Гексли) и бледный эпителиальный слой (слой Генле). В средних и верхних отделах корня волоса все эти три слоя сливаются, и здесь внутреннее корневое влагалище состоит только из полностью ороговевших клеток, содержащих мягкий кератин.
Наружное корневое влагалище образуется из росткового слоя эпидермиса кожи, который продолжается вплоть до волосяной луковицы. Клетки имеют светлую вакуолизированную цитоплазму вследствие наличия в ней значительного количества гликогена.
Волосяная сумка – соединительно-тканная оболочка волоса. В ней различают наружный продольный слой волокон, внутренний и циркулярный слой волокон и базальную мембрану.
51. Железы кожи
В коже человека находится три вида желез: молочные, потовые и сальные. Эти кожные железы обеспечивают терморегуляцию, защиту кожи от повреждений (жировая смазка предохраняет кожу от высыхания, а также от мацерации водой и влажным воздухом), выделение из организма некоторых продуктов обмена веществ. Секрет потовых желез представляет собой жидкость с низкой относительной плотностью. Потовые железы могут быть подразделены на мерокрино-вые и апокриновые. Апокриновые железы находятся лишь в определенных местах кожного покрова, например в подмышечных впадинах, области заднего прохода, коже лба, больших половых губах. Развиваются апокриновые железы в период полового созревания организма и отличаются несколько большими размерами. Секрет их богаче белковыми веществами, которые при разложении на поверхности кожи придают ему особенный, резкий запах. Разновидностью апокриновых потовых желез являются железы век и железы, выделяющие ушную серу. Потовые железы имеют простое трубчатое строение. Они состоят из длинного выводного протока, идущего прямолинейно или слегка извиваясь, и из не менее длинного концевого отдела, закрученного в виде клубочка. Концевые отделы располагаются в глубоких частях сетчатого слоя на границе его с подкожной жировой клетчаткой, а выводные протоки, пройдя через оба слоя дермы и эпидермис, открываются на поверхности кожи так называемой потовой порой. Выводные протоки многих апокриновых желез не образуют потовых пор, а впадают вместе с выводными протоками сальных желез в волосяные воронки. в них содержится высокоактивная щелочная фосфатаза. Кроме секреторных клеток, на базальной мембране концевых отделов располагаются миоэпителиальные клетки. В процессе секреции апикальные концы клеток разрушаются и входят в состав секрета. Функция апокриновых половых желез связана с функцией потовых желез – в предменструальный и менструальный периоды и во время беременности секреция апокриновых желез возрастает. Переход концевого отдела в выводной проток совершается резко. Стенка выводного протока состоит из двухслойного кубического эпителия, клетки которого окрашиваются более интенсивно. Проходя через эпидермис, выводной проток приобретает штопорообразный ход. Здесь его стенка образована плоскими клетками. Сальные железы достигают наибольшего развития в период полового созревания. Больше всего сальных желез на голове, лице и верхней части спины. На ладонях и подошвах они отсутствуют. Секрет сальных желез служит жировой смазкой для волос и эпидермиса кожи. Оно смягчает кожу, придает ей эластичность и облегчает трение соприкасающихся поверхностей кожи, а также препятствует развитию на ней микроорганизмов. В отличие от потовых сальные железы располагаются более поверхностно: в пограничных отделах сосочкового и сетчатого слоев дермы. Сальные железы по строению являются простыми альвеолярными с разветвленными концевыми отделами. Секретируют они по голокриновому типу. Выводной проток короткий, открывается в волосяную воронку. Стенка его состоит из многослойного плоского эпителия. Ближе к концевому отделу количество слоев в стенке протока уменьшается, и он переходит в наружный ростковый слой концевого отдела.
52. Выделительная система
Почки – парные органы, в которых непрерывно образуется моча. Расположены на внутренней поверхности задней брюшной стенки и имеют форму боба. В ворота почек вступают почечные артерии и выходят почечные вены и лимфатические сосуды. Здесь же начинаются мочеотводящие пути: почечные чашечки, почечные лоханки и мочеточники.
Почка покрыта соединительно-тканной капсулой и серозной оболочкой. Вещество почки подразделяется на корковое и мозговое. Корковое вещество темно-красного цвета, распо-лагается общим слоем под капсулой. Мозговое вещество более светлой окраски, разделено на 8—12 пирамид. Вершины пирамид, или сосочки, свободно выступают в почечные чашечки. Опору почки составляет рыхлая соединительная ткань, богатая ретикулярными клетками и ретикулярными волокнами. Паренхима почки представлена эпителиальными почечными канальцами, которые при участии кровеносных капилляров образуют нефроны. Нефрон начинается почечным тельцем, включающим капсулу, охватывающую клубочек из кровеносных капилляров. На другом конце нефрон переходит в собирательную трубку. Собирательная трубка продолжается в сосочковый канал, открывающийся на вершине пирамиды в полость почечной чашечки. В нефроне различают четыре основных отдела: почечное тельце, проксимальный отдел, петлю нефрона с нисходящей и восходящей частями, дистальный отдел. Проксимальный и дистальный отделы представлены извитыми канальцами нефрона. Нисходящая и восходящая части петли являются прямыми канальцами нефрона. Корковое вещество составляют почечные тельца, проксимальные и дистальные отделы нефронов, имеющие вид извитых канальцев. Мозговое вещество состоит из прямых нисходящих и восходящих частей петель нефронов, а также конечных отделов собирательных трубок и сосочковых каналов.
Кровь приносится к почкам по почечным артериям, которые, войдя в почки, распадаются на междолевые артерии, идущие между мозговыми пирамидами. Капсула клубочка по форме напоминает двустенную чашу, в которой, кроме внутреннего листка, имеется наружный листок, а между ними расположена щеле-видная полость: полость капсулы, переходящая в просвет проксимального канальца нефрона. Внутренний листок капсулы проникает между капиллярами сосудистого клубочка и охватывает их почти со всех сторон. Петля нефрона состоит из нисходящей тонкой части и восходящей толстой части. Нисходящая часть – прямой каналец. Стенка образована плоскими эпителиальными клетками, ядросодержащие части которых выбухают в просвет канальца. Цитоплазма у клеток светлая, бедная органеллами. Цитолемма образует глубокие внутренние складки. Через стенку этого канальца происходит пассивное всасывание в кровь воды. Восходящая часть петли также имеет вид прямого эпителиального канальца, но большего диаметра – до 30 мкм. По строению и роли в реабсорбции этот каналец близок к дистальному отделу нефрона. Ди-стальный отдел нефрона представляет собой извитой каналец. Его стенка образована цилиндрическим эпителием, участвующим в факультативной реабсорб-ции: обратном всасывании в кровь электролитов.
53. Органы мочевыделения
Фазы мочеобразования
Мочеобразование – сложный процесс, который осуществляется в нефронах. В почечных тельцах не-фронов происходит первая фаза этого процесса, или фильтрация, в результате чего образуется первичная моча (более 100 л в сутки).
В канальцах нефронов протекает вторая фаза моче-образования, т. е. реабсорбция (облигантная и факультативная), следствием чего является качественное и количественное изменение мочи.
Из нее полностью исчезают сахар и белок, а также снижается ее количество (до 1,5–2 л в сутки), что приводит к резкому возрастанию в окончательной моче концентрации выделяемых шлаков: креатиновых тел – в 75 раз, аммиака – в 40 раз и т. д. Заключительная (третья) секреторная фаза мочеобразования осуществляется в собирательных трубках, где реакция мочи становится слабокислой. Все фазы образования мочи – биологические процессы, т. е. результат активной деятельности клеток нефронов. Юкстагломерулярный аппарат почек (ЮГА), или око-локлубочковый аппарат, секретирует в кровь ренин, который является катализатором образования в организме ангиотензинов, оказывающих сильное сосудосуживающее действие, а также стимулирует продукцию гормона альдостерона в надпочечниках. В состав ЮГА входят юкстагломерулярные клетки, плотное пятно и клетки Гурмагтига. Расположением юкстагло-мерулярных клеток является стенка приносящих и выносящих артериол под эндотелием. Они имеют овальную или полигональную форму, а в цитоплазме – крупные секреторные (рениновые) гранулы, которые не окрашиваются обычными гистологическими методами, но дают положительную ШИК-реакцию. Плотное пятно является участком стенки ди-стального отдела нефрона там, где его прохождение осуществляется рядом с почечным тельцем между приносящей и выносящей артериолами.
Таким образом, ЮГА и ИК – эндокринный комплекс почек, регулирующий общее и почечное кровообращение, через которое оказывается влияние на моче-образование. Непосредственно на функцию нефро-нов воздействуют альдостерон (надпочечники) и вазопрессин, или антидиуретический гормон (гипоталамус). Под влиянием первого гормона усиливается реабсорбция натрия в дистальных отделах нефронов, а под влиянием второго – реабсорбция воды в канальцах нефронов и в собирательных трубках. Лимфатическая система почки представлена сетью капилляров, окружающих канальцы коркового вещества и почечные тельца. В сосудистых клубочках лимфатических капилляров нет. Лимфа из коркового вещества оттекает через футлярообразную сеть лимфатических капилляров, окружающих междольковые артерии и вены, в отводящие лимфатические сосуды 1-го порядка, которые, в свою очередь, окружают дуговые артерии и вены. В эти сплетения лимфатических сосудов впадают лимфатические капилляры мозгового вещества, окружающие прямые артерии и вены. Из этих сосудов лимфа поступает в регионарные лимфатические узлы.
Иннервацию почки осуществляют эфферентные симпатические и парасимпатические нервы и афферентные заднекорешковые нервные волокна.
54. Выделительная система
К мочевыводящим путям относятся почечные чашечки и лоханки, мочеточники, мочевой пузырь и мочеиспускательный канал, который у мужчин одновременно выполняет функцию выведения из организма семенной жидкости. Строение стенок почечных чашечек и лоханок, мочеточников и мочевого пузыря в общих чертах сходно. В них различают слизистую оболочку, состоящую из переходного эпителия и собственной пластинки, подслизистую основу, мышечную и наружную оболочки. В стенке почечных чашечек и почечных лоханок вслед за переходным эпителием располагается собственная пластинка слизистой оболочки, незаметно переходящая в соединительную ткань под-слизистой основы. Мышечная оболочка состоит из двух тонких слоев гладких мышечных клеток: внутреннего (продольного) и наружного (циркулярного). Однако вокруг сосочков почечных пирамид сохраняется только один циркулярный слой гладких мышечных клеток. Наружная оболочка без резких границ переходит в соединительную ткань, окружающую крупные почечные сосуды. Мочеточники обладают выраженной способностью к растяжению благодаря наличию в них глубоких продольных складок слизистой оболочки. Подслизистая основа нижней части мочеточников имеет мелкие альвеолярно-трубчатые железы, по строению напоминающие предстательную железу. Мышечная оболочка мочеточников в верхней половине состоит из двух слоев: внутреннего (продольного) и наружного (циркулярного). Мышечная оболочка нижней части мочеточников имеет три слоя: внутренний и наружный слои продольного направления и средний слой – циркулярного. В мышечной оболочке мочеточников в местах их прохождения через стенку мочевого пузыря пучки гладких мышечных клеток идут только в продольном направлении.
Сокращаясь, они раскрывают отверстие мочеточника независимо от состояния гладких мышц мочевого пузыря. Снаружи мочеточники покрыты соединительно-тканной адвентициальной оболочкой. Слизистая оболочка мочевого пузыря состоит из переходного эпителия и собственной пластинки. В ней мелкие кровеносные сосуды особенно близко подходят к эпителию. Они отсутствуют в переднем отделе дна пузыря, где в него впадают мочеточники и выходит мочеиспускательный канал. Этот участок стенки мочевого пузыря, имеющий форму треугольника, лишен подсли-зистой основы, и его слизистая оболочка плотно сращена с мышечной оболочкой. Здесь в собственной пластинке слизистой оболочки заложены железы, подобные железам нижней части мочеточников.
Мышечная оболочка мочевого пузыря состоит из трех ограниченных слоев: внутреннего, наружного с продольным расположением гладкомышечных клеток и среднего – циркулярного. Гладкие мышечные клетки часто напоминают расщепленные веретена. Прослойки соединительной ткани разделяют мышечную ткань в этой оболочке на отдельные крупные пучки. В шейке мочевого пузыря циркулярный слой образует мышечный сфинктер. Наружная оболочка на верхнезадней и частично на боковых поверхностях мочевого пузыря характеризуется листком брюшины (серозная оболочка), в остальной его части она является адвентициальной.
55. Костный мозг
Костный мозг – центральный кроветворный орган, где находится самоподдерживающаяся популяция стволовых клеток, где образуются клетки как миело-идного, так и лимфоидного ряда.
Красный костный мозг является кроветворной частью костного мозга. Он заполняет губчатое вещество плоских костей и эпифизов трубчатых костей имеет темно-красный цвет и полужидкую консистенцию, что позволяет легко приготовить из него тонкие мазки на стекле.
Ретикулярная ткань структурной основы костного мозга обладает низкой пролиферативной активностью. Строма пронизана множеством кровеносных сосудов микроциркуляторного русла, между которыми располагаются гемопоэтические клетки: стволовые, полустволовые (морфологически неидентифицируе-мые), различные стадии созревания эритробластов и миелоцитов, мегакариобласты, мегакариоциты, лимфобласты, В-лимфоциты, макрофаги и зрелые форменные элементы крови. Лимфоциты и макрофаги принимают участие в защитных реакциях организма.
Гранулоцитопоэтические клетки также располагаются в виде островков, но не связаны с макрофагами. Незрелые клетки гранулоцитарных рядов окружены протеингликанами.
Мегакариобласты и мегакариоциты располагаются в тесном контакте с синусами так, что периферическая часть их цитоплазмы проникает в просвет сосуда через поры. Отделение фрагментов цитоплазмы в виде кровяных пластинок происходит непосредственно в кровяное русло.
В обычных физиологических условиях через стенку синусов костного мозга проникают лишь созревшие форменные элементы крови. Миелоциты и нор-мобласты попадают в кровь только при патологических состояниях организма. Причины такой избирательной проницаемости стенки синуса остаются недостаточно ясными, но факт проникновения незрелых клеток в кровяное русло всегда служит верным признаком расстройства костномозгового кроветворения.
Вышедшие в кровоток клетки выполняют свои функции либо в сосудах микроциркуляторного русла (эритроциты, кровяные пластинки), либо при попадании в соединительную ткань (лимфоциты, лейкоциты) и в периферические лимфоидные органы (лимфоциты). В частности, предшественники лимфоцитов (нулевые лимфоциты) и зрелые В-лимфоциты мигрируют в тимуснезависимые зоны селезенки, где они клонируются на клетки иммунологической памяти и клетки, непосредственно дифференцирующиеся в антитело-продуценты (плазматические клетки) уже при первичном иммунном ответе.
Желтый костный мозг у взрослых находится в диа-физах трубчатых костей. Он представляет собой перерожденную ретикулярную ткань, клетки которой содержат жировые включения.
Васкуляризация. Костный мозг снабжается кровью посредством сосудов, проникающих через надкостницу в специальные отверстия в компактном веществе кости. Войдя в костный мозг, артерии разветвляются на восходящую и нисходящую ветви, от которых радиально отходят артериолы, которые сначала переходят в узкие капилляры.
56. Органы кроветворения
Вилочковая железа
Вилочковая железа – центральный орган лимфоци-топоэза и иммуногенеза. Из костномозговых предшественников Т-лимфоцитов в ней происходит антигенне-зависимая дифференцировка их в Т-лимфоциты, разновидности которых осуществляют реакции клеточного иммунитета и регулируют реакции гуморального иммунитета.
Вилочковая железа – непарный, не до конца разделенный на дольки орган, в основе которого лежит отростчатая эпителиальная ткань, инвагинировавшая в процессе развития так, что базальный слой эпителия с базальной мембраной обращен наружу и граничит с окружающей соединительной тканью, которая образует соединительно-тканную капсулу. От нее внутрь отходят перегородки, разделяющие железу на дольки. В каждой дольке различают корковое и мозговое вещество.
Корковое вещество долек инфильтрировано Т-лим-фоцитами, которые густо заполняют просветы сете-видного эпителиального остова, придавая этой части дольки характерный вид и темную окраску на препаратах. В подкапсулярной зоне коркового вещества находятся крупные лимфоидные клетки – лимфобласты, которые под влиянием гемопоэтических факторов (тимозина), выделяемых эпителиальными клетками стромы, пролиферируют.
Эти предшественники Т-лимфоцитов мигрируют сюда из красного костного мозга. Новые генерации лимфоцитов появляются в вилочковой железе каждые 6–9 ч. Т-лимфоциты коркового вещества мигрируют в кровоток, не входя в мозговое вещество. Эти лимфоциты отличаются по составу маркеров и рецепторов от Т-лимфоцитов мозгового вещества. С током крови они попадают в периферические органы лимфоцитопоэза – лимфатические узлы и селезенку.
Клетки коркового вещества определенным образом отграничены от крови гематотканевым барьером, предохраняющим дифференцирующиеся лимфоциты коркового вещества от избытка антигенов. В его состав входят эндотелиальные клетки гемокапилляров с ба-зальной мембраной, перикапиллярное пространство с единичными лимфоцитами, макрофагами и межклеточным веществом, а также эпителиальные клетки с их базальной мембраной.
Мозговое вещество дольки на препаратах имеет более светлую окраску, так как по сравнению с корковым веществом содержит меньшее количество лимфоцитов. Лимфоциты этой зоны представляют собой ре-циркулирующий пул Т-лимфоцитов и могут входить и выходить в кровоток через посткапиллярные венулы и лимфатические сосуды. Особенностью ультрамикроскопического строения отростчатых эпителиальных клеток является наличие в цитоплазме гроздевидных вакуолей и внутриклеточных канальцев, поверхность которых образует микровыросты. Базальная мембрана редуцируется.
Васкуляризация. Внутри органа артерии ветвятся на междольковые и внутридольковые, которые образуют дуговые ветви. От них почти под прямым углом отходят кровеносные капилляры, образующие густую сеть, особенно в корковой зоне.