Ихтиостега: Как и почему позвоночные вышли на сушу

Ихтиостега: Как и почему позвоночные вышли на сушу
Интересная статья с канала «Цитадель адеквата» яндекс-дзена.
Есть такое странное понятие «переходная форма». Биологом любителям, не любившим биологию ещё в школе, что негативно сказывалось на успеваемости и усвоении предмета, переходных форм часто не хватает. Профессионалы же вовсе считают эту сущность избыточной. Ископаемое существо будет классифицировано либо как лопастепёрая рыба, либо уже как амфибия. Без перехода . Так как для обозначения ещё не земноводных, но уже и рыб, в противном случае пришлось бы вводить отдельный класс.
Тем не менее, ихтиостеги именно переходная форма. Поскольку, вопрос, к амфибиям ли или рыбам относить этих существ, до сих пор остаётся дискуссионным. Слишком уж хитро перемешаны признаки. Кратко описать этих животных можно, как ещё, вроде бы, рыб. Но уже, всё-таки, с ногами.
Обретение рыбами ног случилось около 370 миллионов лет назад — в конце девона . Начало же и середина этого периода были потрачены планетой на создание предпосылок для выхода позвоночных на сушу. И задача эта была не такой уж тривиальной. Выманить на берег растения, а затем червей и членистоногих, оказалось не сложно и удалось ещё в силуре. Но позвоночные за ними не пошли.
Первые волны колонизации суши прошли через её естественную границу с морем — приливно-отливную зону. Жизнь, тогда привязанная исключительно к мелководьям, постоянно рисковала оказаться под лучами Солнца в период низкой воды. И, естественно, приспосабливалась к таким приключениям. Постепенно, некоторым растениям и животным новая среда даже стала нравиться. Однако, позвоночные не входили в число любителей воздуха. Они, просто, были слишком подвижны, — умение быстро плавать являлось главным их достоинством, — поэтому редко оказывались на берегу, успевая уйти в глубину при отливе.
Появление в середине девона на суше растительности, превративший реки, до того кристально чистые и безжизненные, в мутные, переполненные смытой дождями органикой потоки, ситуацию изменило. Рыбы заселили богатые пищей пресные воды, и тут уж столкнулись с новыми, неведомыми морским жителям сложностями. Вода всё время куда-то текла. В зависимости от дождей, реки то разливались, затапливая обширные пространства, то почти пересыхали. И когда вода спадала, куда бежать, понятно не было. Очень легко было застрять в какой-нибудь луже. Причём, без кислорода. Избыток органики в палеозое и мезозое немедленно превращал стоячую или даже слабопроточную воду в болото.
Тут-то рыбам уже пришлось приспосабливаться, для начала освоив альтернативные способы дыхания: с помощью плавательного пузыря и кожи. То и другое не представляет сложности. В большей или меньшей степени такая адаптация есть у большинства современных пресноводных рыб, в случае дефицита кислорода в воде, начинающих захватывать воздух с поверхности. Но сейчас рыбы уже не стремятся выбраться на сушу, где их что-либо хорошее не ожидает. Птицы и звери не дремлют. В девоне же ситуация выглядела иначе. Воздушная среда воспринималась прежде всего, как безопасная (если умеешь дышать воздухом), ибо полностью демилитаризованная. Суша была населена лишь членистоногими, которых позвоночные достойными противниками уже не считали. Даже гигантских.
Таким образом, ныне слишком опасная и непривлекательная для рыб среда, — заболоченные пространства с массой мелких, скорее заслуживающих именоваться лужами, водоёмов, — в девоне была очень привлекательной. В любой из таких луж полуметровая лопастепёрая рыба оказывалась неуязвимым сверххищником. Вот только ресурсы крошечного и, скорее всего, временного водоёма были ограничены. Доев в нём всё живое, нужно было перебираться в следующий.
Ползком. Мускулистые плавники лопастепёрых рыб, в отличие от плавников рыб современного образца — лучепёрых, хорошо подходили для того, чтобы ползти, волоча по земле покрытое чешуёй брюхо. Но долго так двигаться рыба не могла. Она просто задыхалась.
Для дыхания с помощью уже превратившегося в лёгкое плавательного пузыря, требуется чуть раздвигать рёбра. Что затруднительно делать, пластаясь брюхом по земле. Слишком затратно с точки зрения расхода энергии. Дышать приходилось кожей, и это работало, лишь пока кожа не высыхала. Вывод же был очевиден: ползти нужно быстро. А для этого требуются хоть какие-то ноги. С пальцами и когтями, чтобы цепляться за грунт.

Ихтиостега. Фото и иллюстрации взяты из открытых источников и принадлежат их авторам
Ихтиостега представляла собой рыбу, уже адаптированную для перемещения между мелкими водоёмами посуху. На суше эти существа ещё, видимо, не охотились. Ходить в обычном понимании не могли, — лишь ползали. Тем не менее, уже являлись земноводными по образу жизни. Пребывание и передвижение вне воды для них более не являлось шагом отчаянным, предпринимаемым в крайнем случае, последней попыткой спастись, как для их предков — двоякодышащих рыб. Просто, рабочий момент. Не более того.
источник: https://zen.yandex.ru/media/id/5fd837cc15ece8677e55f87a/ihtiostega-kak-i-pochemu-pozvonochnye-vyshli-na-sushu-607911374825fd7864a8e166
Образование новых родов, семейств и далее более меньших таксонов носит название макроэволюция. Микроэволюция происходит внутри вида, а вот макроэволюция – надвидовой процесс. Притом, в основе макроэволюции лежат принципы микроэволюции, поэтому особых различий между этими процессами нет. Соответственно, макроэволюции присущи те же факторы: наследственная изменчивость, естественный отбор и борьба за существование. Как можно заметить, мы не можем наблюдать макроэволюция также часто как микро-. Дело в том, что надвидовая эволюция происходит в какие-то переломные, грандиозные моменты существования вида.
Палеонтологические доказательства макроэволюции
Так как палеонтология изучает ископаемые останки животных и растений, то эта наука дает нам представление о том, как развивалась жизнь на Земле. В более ранних пластах обнаруживаются останки беспозвоночных, а более поздних – хордовых. Из таких доказательств и складывается картина развития мира.
Филогенетика позволяет установить родство между организмами. Таким образом, мы узнаем о том, какие предки были у такого или иного вида. Очевидно, что ныне живущие животные и растения претерпевали изменения под влиянием окружающей среды. Эти изменения приводили к появлению новых видов, семейств и так далее.

Из курса зоологии известно, что усложнение строения отрядов позвоночных животных шло так: рыбы, земноводные, рептилии, птицы, млекопитающие. Об этом говорят, например, их анатомические свойства. Взять то же самое строение сердца и кровеносной системы. У рыб сердце двухкамерное, кровь не разделена на артериальную и венозную, один круг кровообращения. Амфибии имеют трехкамерное сердце, кровь смешивается в желудочке, два круга кровообращения. У рептилий появляется неполная перегородка в желудочке, но он, как и у земноводных, один. Поэтому кровь все еще частично смешивается. Птицы и млекопитающие имеют самую развитую кровеносную систему: сердце четырехкамерное, два круга кровообращения и полное разделение на артериальную и венозную кровь.
К этим анатомическим доказательствам присоединяются и палеонтологические. Две находки являются классическими доказательствами макроэволюции.
Ихтиостег
– переходная форма между пресноводными кистеперыми рыбами и земноводными. Именно этот организм первым вышел на сушу. Кистеперые рыбы – весьма древние, сейчас живут лишь латимерии. Их называют живыми ископаемыми. Обязательно нужно запомнить, что ихтиостег находится в ряду именно между кистеперыми рыбами и земноводными. Сделать это можно при помощи логики и воображения. Для того чтобы выйти на сушу, нужна опора. Кистеперые рыбы, само название, часть слова «кисть», прямо как у нас, указывает на развитие плавников. Первыми вышли именно земноводные, хоть они и привязаны к воде. Так мы запоминаем положение ихтиостега.
Археоптерикс
– переходная форма между рептилиями и птицами. Само название, говорит нам о том, где примерно находится эта форма. Внешний вид археоптерикса – смесь ящерицы и птицы. Размер тела был небольшим, примерно с голубя. У этого животного была цевка на задних конечностях, перья. Казалось бы, это птица. Однако, кроме вышеперечисленных признаков, археоптерикс имел много хвостовых позвонков, зубы, а также брюшные ребра. Много хвостовых позвонков наталкивают нас на мысли о том, как ящерицы отбрасывают хвост. Зубов у современных птиц нет, ведь костные челюсти – очень тяжелая конструкция. Тело птицы максимально приспособлено к полету, а значит, облегчено. У птиц нет ребер. Зато есть киль.
Эмбриологические доказательства макроэволюции
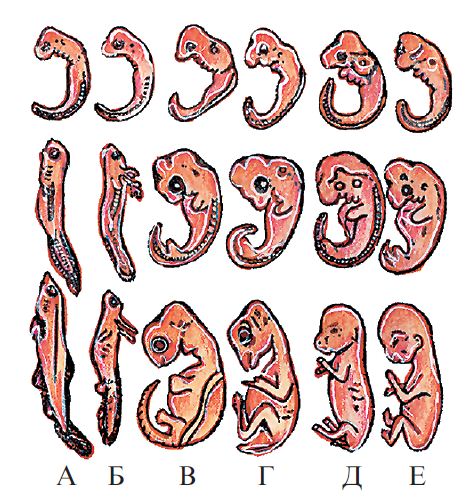
Эмбриональное развитие организмов, принадлежащих к различным классам позвоночных: А — рыба; Б — саламандра; В — черепаха; Г — птица; Д — заяц; Е — человек
Зародыши у рыб, земноводных, рептилий, млекопитающих и человека в том числе, имеют сходства на ранних стадиях развития. К таким признакам относятся форма тела, наличие жабр, хвоста, зачатков конечностей. Также стоит отметить, что вначале закладывается хорда, только потом позвоночник. Далее, по мере развития организма, то есть онтогенеза, различия начинают преобладать над сходствами.
Признаки расходятся последовательно, согласно таксонам, к которым принадлежит организм. Сначала появляются признаки класса, затем отряд, после род, и, наконец-то, вида.
Задание EB1621
Установите соответствие между примерами и доказательствами эволюции: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
|
ПРИМЕРЫ |
ДОКАЗАТЕЛЬСТВА ЭВОЛЮЦИИ |
|
А) гомологичные органы Б) ископаемые переходные формы В) атавизмы Г) единство плана строения тела позвоночных Д) окаменелости Е) рудименты |
1) сравнительно-анатомические 2) палеонтологические |
Сравнительно-анатомический доказательства эволюции основаны на морфологических признаках сравниваемых объектов.
Палеонтологический доказательства – на ископаемых остатках.
Гомологичные органы – органы, которые сходны по своему строению, но могу выполнять очень разные функции. Например, передние конечности позвоночных. Лапа кошки и крыло птицы – гомологичные органы. Они имеют общий план строения, но лапы нужны для перемещения по горизонтальной поверхности, а крылья – для полета. Так как все сводится к строению, то это сравнительно-анатомическое доказательство эволюции.
Ископаемые переходные формы – палеонтологическое доказательство. К таким относят, например, археоптерикса – переходная форма между рептилиями и птицами.
Атавизмы – признаки, которые были у предков. В ходе эволюции эти признаки были утрачены, однако они могут проявиться у отдельных особей. Примерами атавизмов являются многососковость, развитый хвост, полностью покрытое волосами лицо.
Рудименты – органы, которые раньше выполняли какие-то важные функции, но в ходе эволюции необходимость в них отпала. В отличии от атавизмов, рудименты характерны не для отдельной особи, а для всего вида. Рудиментарные органы закладываются в эмбриогенезе, но перестают развиваться, оставаясь в таком недоразвитом состоянии. Например, аппендикс, полулунная складка (третье веко).
Единство плана строения позвоночных – сравнительно-анатомическое доказательство.
Окаменелости – палеонтологическое доказательство.
Ответ: 121121
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB1521
Прочитайте текст. Выберите три предложения, в которых даны описания идиоадаптаций у земноводных. Запишите в таблицу цифры, под которыми они указаны.
(1)Появление лёгких у земноводных в процессе эволюции дало им возможность выйти на сушу. (2)Также при выходе земноводных на сушу важную роль сыграли пятипалые конечности рычажного типа. (3)У квакш, ведущих древесный образ жизни, на кончиках пальцев находятся присоски, с помощью которых квакши могут прилипать к любым поверхностям. (4)С помощью эластичных перепонок на пальцах некоторые виды квакш могут планировать на расстояние до двух метров. (5)Благодаря направлению глаз вперёд квакши совершают безошибочные прыжки до добычи или соседней ветки. (6)Кровеносная система квакш, как и всех земноводных, содержит трёхкамерное сердце и два круга кровообращения.
Идиоадаптация — эволюционный процесс, приводящий к повышению приспособленности организмов к разным условиям обитания и существенно не отражающийся на общем уровне организации, в отличие от ароморфоза.
Появление легких у земноводных – это ароморфоз, до амфибий никто из животных на сушу не выходил.
Пятипалые конечности рычажного типа позволили земноводным перемещаться по суше, это тоже ароморфоз.
Присоски и перепонки у квакш – идиоадаптация, т.к эти присоски пример частного приспособления конкретного семейства.
Направленные вперед глаза квакш – идиоадаптация, это не повлияло на общий уровень организации, просто так эффективнее охотиться.
Изменения в кровеносной системе – ароморфоз. До земноводных, у рыб, сердце было двухкамерное, кровь не разделялась на артериальную и венозную, круг кровообращения был всего один. Так как амфибии вышли на сушу, у них появился второй круг кровообращения – легочный.
Ответ: 345
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB2018D
Проанализируйте таблицу «Пути эволюции». Заполните пустые ячейки таблицы, используя термины и понятия, приведённые в списке. Для каждой ячейки, обозначенной буквой, выберите соответствующий термин или соответствующее понятие из предложенного списка.
|
Пути эволюции |
Характеристика | Примеры признаков у организмов |
| Ароморфоз | ___________(Б) | Появление головного мозга у рыб |
| __________(А) | Упрощение уровня организации, утрата отдельных органов | Утрата пищеварительной системы у ленточных червей |
| Идиоадаптация | Частные приспособления к условиям среды |
__________(В) |
Список терминов и понятий:
- переход к сидячему образу жизни
- общая дегенерация
- биологический регресс
- крупные изменения в строении, повышение уровня организации
- отсутствие конечностей у змей
- специализации к условиям обитания
- появление оболочек у зародыша пресмыкающихся
- утрата корней у некоторых растений-паразитов
Упрощение уровня организации, утрата отдельных органов – общая дегенерация, 2).
Ароморфоз – крупные изменения в строении, повышение уровня организации,4).
Идиодаптации – отсутствие конечностей у змей, т.к утрата органов не приводит к снижению уровня организации, а также к паразитическому или малоподвижному образу жизни, 5).
Ответ: 245
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB1618D
Установите соответствие между животными и группами организмов: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
| ЖИВОТНЫЕ | ГРУППЫ ОРГАНИЗМОВ |
|
А) кальмары Б) ихтиозавры В) тюлени Г) дельфины Д) морские черепахи Е) акулы |
1) первичноводные 2) вторичноводные |
Первичноводные — водные животные, у которых все предки жили в воде.
Вторичноводные — водные животные, предки которых жили на суше.
Все предки моллюсков жили в воде, следовательно, кальмар – первичноводное животное.
Ихтиозавры – вымершее морское пресмыкающиеся, то есть рептилия, вернувшаяся в воду, вторичноводное животное.
Тюлени и дельфины – морские млекопитающие, млекопитающие обитают на суше, а эти вернулись в воду, значит, они вторичноводные животные.
Морские черепахи – рептилии, которые вернулись в воду, но яйца откладывают на суше. Вторичноводные животные.
Акулы – хрящевые рыбы, все предки рыб обитали в воде, первичноводные животные.
Ответ: 122221
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB1619t
Установите соответствие между признаками макроэволюции и микроэволюции: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
| ПРИЗНАКИ | ТИП ЭВОЛЮЦИИ |
|
А) перестройка структуры популяций Б) на процесс влияют дегенерации и ароморфозы В) эволюция в пределах вида Г) важную роль играют популяционные волны Д) формирование надвидовых групп |
1) макроэволюция 2) микроэволюция |
Запишите в таблицу выбранные цифры под соответствующими буквами.
На уровне до вида происходит микроэволюция, а выше вида – макроэволюция. Значит, А), В) и Г) – 2; Д) – 1.
Ароморфозы и дегенерации – крупные изменения, влияющие на положение вида в системе, поэтому это макроэволюция.
Ответ: 21221
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB2619t
Назовите атавизмы и рудименты, которые можно наблюдать у человека.
Примеры атавизмов у человека: удлиненный хвостовой отросток, сплошной волосяной покров на лице, многососковость.
Примеры рудиментов у человека: третье веко, аппендикс, околоушные мышцы.
Ответ: см. решение
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB1919D
Установите последовательность эволюционных явлений у позвоночных животных. Запишите в таблицу соответствующую последовательность цифр.
- расцвет динозавров
- появление приматов
- расцвет панцирных рыб
- появление питекантропов
- появление стегоцефалов
По эволюционному порядку мы знаем, что вначале идут рыбы, потом амфибии, потом рептилии, потом птицы и млекопитающие. В этом задании необходимо сопоставить наши знания с вариантами.
Первыми будут панцирные рыбы.
Затем стегоцефал – промежуточное звено между амфибиями и рептилиями.
Далее динозавры, как предки современных рептилий.
Остались варианты 2) появление приматов и 4) появление питекантропов. Так как предки человека – приматы, то и в очереди они будут стоять раньше.
Ответ: 35124
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB11235
Какое явление называют макроэволюцией?
- Макроэволюция — это совокупность эволюционных преобразований, протекающих на уровне надвидовых таксонов.
- Сущность макроэволюционных преобразований составляет изменение генетической структуры видов и надвидовых структур.
- Это процесс образования надвидовых систематических групп.
Ответ: см. решение
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB11042
Назовите не менее 3-х особенностей наземных растений, которые позволили им первыми освоить сушу. Ответ обоснуйте.
- Появление покровной ткани — эпидермиса с устьицами для защиты от испарения.
- Появление слаборазвитой проводящей системы для транспорта веществ.
- Появление механической ткани для выполнения опорной функции. Наличие ризоидов для закрепления в почве.
Ответ: см. решение
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB22142
На рисунке изображён археоптерикс — вымершее животное, обитавшее 150—147 млн лет назад.

Используя фрагмент «Геохронологической таблицы», определите, в какой эре и каком периоде обитал данный организм?
Это животное учёные считают переходной формой. Назовите классы, к которым можно отнести изображённое животное. Какие черты внешнего строения позволяют отнести его к этим классам?
Геохронологическая таблица
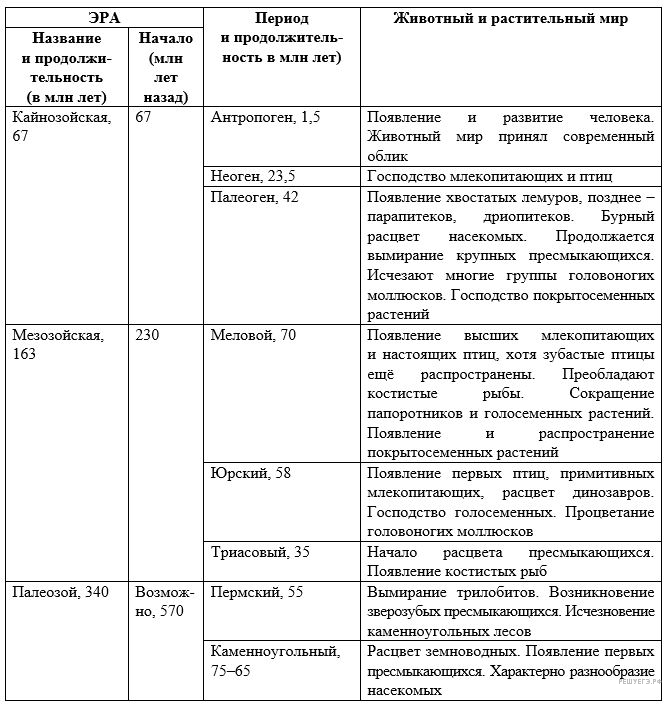
Для того, чтобы ответить на вопросы необходимо воспользоваться соответствующими колонками Геохронологической таблицы и произвести элементарные математические подсчеты.
Эра: указан период обитания археоптерикса: 150—147 млн лет назад. Производим вычисления с помощью второй колонки, в которой указано начало периодов. Мезозойская начало 230 млн лет назад, а Кайнозойская 67 млн лет назад. Значит, Археоптерикс жил в Мезозойскую эру.
Период: берем начало эры 230 млн лет назад вычитаем продолжительность Периодов, — Триасовый 230-35=195 млн лет назад;
минус Юрский 195-58=137 млн лет назад. Получается что Археоптерикс жил в Юрском периоде.
Предка определяем по 4 колонке (или пользуемся своими знаниями).
- Эра – Мезозойская; Период – Юрский;
- к рептилиям животное можно отнести на основании наличия челюсти с зубами, длинного хвоста и развитых пальцев;
- к птицам животное можно отнести на основании наличия перьевого покрова и крыльев
Ответ: см. решение
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB21942
Рассмотрите рисунок с изображением животных, обитающих на разных материках, и определите,
(А) какие виды изображены, (Б) какую группу доказательств эволюции они иллюстрируют и (В) тип изоляции, который привел к формированию таких видов.

Для каждой буквы выберите соответствующий термин из предложенного списка
- биологическая
- виды-эндемики
- аллопатрические виды
- географическая
- сравнительно-анатомические
- биогеографические
- реликтовые виды
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
Виды, изображенные на рисунке — реликтовые. 7)
Они иллюстрируют биогеографические доказательства эволюции. 6)
В задании сказано, что животные располагаются на разных материках, тип изоляции — географическая изоляция. 4)
Ответ: 764
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB12389
Установите правильную последовательность эр в истории Земли.
- Протерозойская
- Кайнозойская
- Архейская
- Палеозойская
- Мезозойская
| № | Эпоха |
| 1 | Архей |
| 2 | Протерозой |
| 3 | Палеозой |
| 4 | Мезозой |
| 5 | Кайнозой |
Ответ: 31452
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB10833
Расположите животных в порядке повышения у них интенсивности обмена веществ в процессе эволюции.
- Млекопитающие
- Хрящевые рыбы
- Земноводные
- Костные рыбы
- Пресмыкающиеся
Интенсивность обмена веществ повышается вместе с повышением уровня организации.
Ответ: 24351
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB10834
Установите, в какой последовательности должны располагаться растения в соответствии с их усложнением в процессе эволюции.
- полевой хвощ
- береза бородавчатая
- ель обыкновенная
- кукушкин лен
Кукушкин лен — мох.
Ель — голосеменное растение.
Береза — покрытосеменное растение
Ответ: 4132
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB19050
Установите соответствие между характеристикой эволюционного процесса и процессом, которому соответствует данная характеристика
| ХАРАКТЕРИСТИКА | ЭВОЛЮЦИОННЫЙ ПРОЦЕСС |
|
А) ведёт к видообразованию Б) ведёт к формированию надвидовых таксонов В) происходит в популяциях Г) сопровождается мутационным процессом Д) происходит в течение длительного исторического периода (миллионы лет) Е) характеризуется биологическим прогрессом или регрессом |
1) микроэволюция 2) макроэволюция |
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
Сразу видны приставки, которые и характеризуют данные виды эволюции. Макро — большой, микро — маленький
Если сравнивать виды и надвидовые таксоны, то надвидовые таксоны будут больше, значит, к формированию надвидовых таксонов относится макроэволюция, а к видообразованию — микроэволюция
Популяции меньше, чем вид, а за видообразование отвечает микроэволюция, значит, в популяциях происходит микроэволюция.
Сопровождается мутационным процессов микроэволюция.
Длительное историческое время — макроэволюция.
Биологический прогресс или регресс — макроэволюция.
Ответ: 121122
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB14138
Установите соответствие между примером и типом доказательств эволюции животного мира, который он иллюстрирует.
| ПРИМЕР | ТИП ДОКАЗАТЕЛЬСТВ |
|
A) филогенетический ряд лошади Б) наличие копчика в скелете человека B) перо птицы и чешуя ящерицы Г) отпечатки археоптерикса Д) многососковость у человека |
1) сравнительно-анатомические 2) палеонтологические |
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
Филогенетический ряд — палеонтология. 2)
Копчик — анатомия. 1)
Перо птицы и чешуя ящерицы — анатомия. 1)
Отпечаток — палеонтология. 2)
Многососковость – анатомия. 1)
Ответ: 21121
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB23017
Установите соответствие между примерами и методами изучения эволюции: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
| ПРИМЕР | МЕТОД |
|
А) отпечаток семенного папоротника в пластах каменного угля Б) сходство зародышевого развития хордовых на ранних этапах развития В) рудимент тазового пояса питона Г) появление развитого хвостового отдела позвоночника у человека Д) филогенетический ряд моллюсков Е) окаменелость белемнита |
1) сравнительно-анатомический 2) эмбриологический 3) палеонтологический |
Запишите в таблицу выбранные цифры под соответствующими буквами.
Посмотрим на методы:
Сравнительно-анатомический. Сравнение и изучение строения.
Эмбриологический — сравнение и изучение эмбрионов.
Палеонтология — изучение ископаемых остатков и их отпечатков.
А) отпечаток — палеонтология. 3)
Б) Зародыш — эмбриология. 2)
В) Рудимент таза — анатомия. 1)
Г) Хвостовой отдел позвоночника — анатомия. 1)
Д) Филогенетический ряд-палеонтология. 3)
Филогенетические ряды − ряды видов, последовательно сменявших друг друга в процессе эволюции различных групп животных и растений.
Е) Окаменелость — палеонтология. 3)
Ответ: 321133
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB10330
Результатом эволюции является
- появление новых засухоустойчивых сортов растений
- возникновение новых видов в изменившихся условиях среды
- выведение высокопродуктивных пород крупного рогатого скота
- формирование новых приспособлений к жизни в изменившихся условиях
- сохранение старых видов в стабильных условиях обитания
- получение высокопродуктивных бройлерных кур
Разберем варианты ответа и просто подумаем.
Засухоустойчивые растения не появляются в результате эволюции, их выводят селекционеры.
Возникновение новых видов в изменившихся условиях среды — эволюция
Выведение порог скота — точно селекция.
Приспособление — часть эволюции.
Сохранение старых видов в стабильных условиях — тоже эволюция.
Разведением бройлерных кур эволюция не занимается.
Ответ: 245
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Задание EB10333
К палеонтологическим доказательствам эволюции относят
- остаток третьего века у человека
- отпечатки растений на пластах каменного угля
- окаменевшие остатки папоротников
- рождение людей с густым волосяным покровом на теле
- копчик в скелете человека
- филогенетический ряд лошади
Палеонтология занимается изучением ископаемых остатков животных организмов, их следов, отпечатков.
К данному описанию подходят варианты 2,3 и 6
Ответ: 236
pазбирался: Ксения Алексеевна | обсудить разбор | оценить
Ксения Алексеевна | Просмотров: 4.7k
| † Ихтиостега |
| Часть скелета в Палеонтологическом музее им. Ю. А. Орлова, Москва |
| Модель в Государственном музее естествознания Штутгарта |
| Без ранга: | Двусторонне-симметричные |
| Семейство: | † Ichthyostegidae |
Международное научное название Ichthyostega
Säve-Söderbergh, 1932
Ихтиосте́га
[2] (лат. Ichthyostega, от др.-греч. ἰχθυο- + στέγη — рыбья кровля) — род вымерших челюстноротых из семейства Ichthyostegidae клады тетраподоморф (Tetrapodomorpha), живших в девонском периоде[3] и представляющий собой первое промежуточное звено между рыбами и амфибиями. Этот род не является прямым предком современных видов, предки которых — лепоспондилы (Lepospondyli) — появились в каменноугольном периоде.
У ихтиостег были лапы, но они, возможно, не использовались для ходьбы по суше. Ихтиостеги имели хвостовой плавник и некоторые органы чувств, функционирующие только в воде. Тело их было покрыто мелкими чешуйками.
По мнению некоторых учёных, ихтиостеги могут считаться переходными формами между кистепёрыми рыбами и наземными позвоночными.
История изучения
В 1932 году Сёве-Седерберг описал четыре вида ихтиостег из верхнего девона, найденных в восточной части Гренландии. Эти виды могли быть синонимичными, потому что их морфологические различия не были резко выражены. Виды различались в пропорциях и строении черепа. Сравнение было проведено на четырнадцати экземплярах, собранных в 1931 году Датской Восточной Гренландской экспедицией. Дополнительные экземпляры были собраны между 1933 и 1955 годами.
Род ихтиостег находится в родстве с акантостегой (Acanthostega gunnari
), также обнаруженной в восточной Гренландии. По сравнению с акантостегой череп ихтиостеги выглядит более рыбообразным, однако её пояс передних конечностей сильнее и лучше адаптирован к передвижению.
Описание
Ихтиостеги были около 1,5 метров длиной и имели по семь пальцев на задних лапах. Точное количество пальцев на передних лапах пока не установлено, но вероятно, что их было тоже семь. На хвосте у них был плавник «рыбьего» типа, поддерживаемый невральными и гемальными дугами.
Ноздри располагались у нижнего края челюстей. Слёзная кость примыкает к ноздре, но не к глазнице. Межвисочная кость отсутствует (это обстоятельство исключает возможность происхождения большинства темноспондилов от этого рода). Заднетеменная кость непарная. Челюстная кость соприкасается с квадратноскуловой. Сохраняются подкрышечные и предкрышечные кости. Носовые кости широкие. Глазницы овальные и располагаются в центральной части черепа. Хорда через отико-окципитальную часть мозговой коробки доходит до гипофизарной ямы.
РАННИЕ ТЕТРАПОДЫ
КЛАСС ЧЕТВЕРОНОГООБРАЗНЫЕ (Tetrapodomorpha или Stegocephali)
Возникли, возможно, на границе среднего и позднего девона, 380 млн. лет назад. Окаменелости четвероногообразных известны из позднего девона Гренландии, России, Австралии. Обнаруженные следы тетрапода также известны из раннего–среднего девона Австралии и среднего–позднего девона Бразилии, правда, пальцы на этих отпечатках не видны.
Развивались в сторону приспособления к дыханию атмосферным воздухом (внутренние ноздри, легкие) и преобразования парных плавников в пятипалые конечности. Поначалу имели конечности с разным количеством пальцев (6–8), которое при выходе животных на сушу сократилось до пяти лишь в карбоне. Дали начало земноводным и, просуществовав около 100 млн. лет, вымерли в карбоне. Тетраподы произошли от древнейших среднедевонских остеолепиформов, которые еще сохранилив своем строении некоторые поролепиформовые признаки.
Предки тетрапод обитали в мелководных, густо заросших растительностью водоемах непостоянной конфигурации. Судя по всему, конечности развились для того, чтобы передвигаться по дну водоемов (это особенно важно, когда водоем обмелел настолько, что спина уже начинает торчать наружу) и пробираться сквозь плотные заросли водных растений; особенно же полезными конечности оказались для того, чтобы при пересыхании водоема переползти посуху в другой, соседний.
Прежде термин стегоцефалы означал всех позвоночных, имеющих конечности с пальцами и суставами (chiridii), кроме лиссамфибий и амниот. Теперь (Laurin, 1998), этот таксон определен, как все хоанаты, более близко связанные с темноспондилиями, чем с Panderichthys (самый близкий известный родственник четвероногов, еще сохранивший парные плавники). Поэтому Stegocephali включают весь таксон имеющих пальцы и некоторые единицы (Elginerpeton, Metaxygnathus, Ventastega, и Hynerpeton), которые могли сохранить парные плавники (наличие пальцев у них неизвестно из-за фрагментарности находок).

Стегоцефалы (Stegocephali), или панцирноголовые. Характерным для стегоцефалов был сплошной панцирь из кожных костей, прикрывавший голову сверху и с боков (стегальный череп), так что оставались отверстия только для ноздрей, глаз и теменного глаза, который у них был, очевидно, хорошо развит. Кроме того, у многие виды имели и брюшной панцирь, состоявший из налегающих друг на друга костных чешуек. Почти все стегоцефалы вымерли в каменноугольный период, 250 млн. лет назад и в течение триаса доживали свой век лишь немногочисленные, крайне специализированные лабиринтодонты. Однако несколько разновидностей продержались в изолированной части Австралии, на ее юго-востоке еще 130 миллионов лет, до раннего мела.
Никто из самых ранних четвероногов не имел кисти, способной сгибаться, потому что у них отсутствует поверхность для сгибающей мышцы на фалангах. Из-за этого им было трудно передвигаться по твердым (камни) поверхностям. Недостаток гибкости удерживал четвероногов ближе к мягкой почве, сдерживая их проникновение вглубь суши.
Ранние тераподоморфы унаследовали S-образное изгибание тела при передвижении и дыхание воздухом от своих предков — саркоптеригий. Большинство из них были крупными (1–2 м) и имели характерные рыбоядные зубы. Грудные и тазовые плавники превратились в конечности, костные элементы плавников развили более сложные схемы сочленения. Спинные срединные и анальные плавники были утеряны. Плечевой пояс потерял связь с задней частью черепа, что дало большую подвижность пояса и головы. Тазовый пояс стал более крепко присоединенным к позвоночному столбу, для поддержки задних конечностей. Мускулы конечностей увеличились в размере и сложности; вентральные мускулы (флексоры) поднимают тело от земли, спинные мускулы (экстензоры) перемещают конечности и тело вперед. Первые четвероноги уже имели легкие, которые иногда использовались как органы дыхания, унаследованные от рыб; они также вероятно имели жабры, так как сохранились следы жаберных створок.

Филогенетические изучения показали несколько развитых характеристик (синапоморфии) присущих стегоцефалам:
Утрата нескольких черепных костей. У пандерихтид (группа саркоптеригий, наиболее близко связанная со стегоцефалами), череп был твердо соединен с плечевым поясом несколькими костями, которые рано исчезли в эволюции наземных позвоночных. Потеря этих костей также привела к появлению подвижной шеи, которая позволяет голове перемещаться относительно туловища. Это разъединение позволяет голове оставаться относительно устойчивой при ходьбе.
Потеря костей жаберной крышки, прикрывавших жаберную камеру у костных рыб. Жаберная крышка не была нужна ранним хоанатам, потому что они утратили внутренние жабры своих предков. Однако, жаберная крышка, возможно, исчезла раньше внутренних жабр. Увеличение эффективности дыхания воздухом вело к сокращению или модификации внутреннего аппарата жабры. Одна из этих костей, подъязычно-челюстная, стала стременем, костью, которая, в конечном счете, играть центральную роль для слуха на суше. Освобождение плечевого пояса от тыла черепа, возможно, также было следствием сдвига от дыхания жабры и связано с множеством изменений в структуре плечевого пояса непосредственно.
Сокращение нотохорды. Тело позвонка у остеолепиформ тонкое и окружает нотохорду не сжимая ее сильно. У стегоцефалов тело позвонка толстое, и позвонки сжимают нотохорду. Специальные соединительные поверхности (zygapophyses) связывают невральные дуги друг с другом. Нотохорда остеолепиформ простиралась до области, близкой к гипофизу. Стегоцефалы имеют более короткую нотохорду, которая не проходит в черепную крышку.
Таз кистеперых рыб состоит из двух изолированных наборов костей, которые не входят в контакт со спинным хребтом или друг другом. Напротив, две половины тазового пояса четвероногов соединяются вентрально, образуя лонное сращение и соединяются со спинным хребтом через крестцовые ребра. Это позволяет четвероногам передавать вес тела на задние конечности. У остеолепиформ не было никакой костной связи между тазовым поясом и их осевым скелетом.
Утрата кожных плавниковых лучей (измененная чешуя, которая поддерживает плавники). Это просто представляет собой устранение структуры, которая больше не была необходима и, возможно, даже была вредна на земле.
Эти особенности не появились одновременно и внезапно. Самые старые известные стегоцефалы, типа ихтиостеги и акантостеги, обладают промежуточными состояниями некоторых из этих особенностей и испытывают недостаток других. Например, ихтиостега сохранила относящуюся к жаберной крышке кость, которая была частью комплекса жаберной крышки, покрывавшей камеру жабры у остеолепиформ. Акантостега сохранила косточку, которая является одним из элементов, связывавших плечевой пояс с черепом у остеолепиформ.
Нотохорда ихтиостеги и акантостеги еще простиралась глубоко в черепную крышку, и большинство их хвостовых позвонков испытывало недостаток зигапофисов. Связь между крестцовым ребром и тазовым поясом акантостеги все еще плохо определялась. Наконец, и ихтиостега и акантостега сохраняют элементы в хвосте, указывающие, что этот таксон все еще имел хвостовой плавник.
У современных четвероногов найдены еще некоторые мягкие анатомические особенности, не присущие никаким другим живущим позвоночным:
- Наличие слоя мертвых роговых клеток, уменьшающих потерю испаряемой воды.
- Большинство четвероногов развило мускульный язык с железами, за исключением некоторых лиссамфибий, которые имеют только первичный язык, подобно рыбам. Первичный язык — просто мясистый сгиб на дне рта, который испытывает недостаток соответствующих мускулов и имеет ограниченную подвижность.
- Околощитовидная железа, вовлеченная в управление уровнем кальция в крови.
- Железа, расположенная спереди глаза и выделяющая маслянистую жидкость для смазывания глаза.
- Орган Якобсона, расположенный на небе и предназначенный для обоняния продуктов питания во рту.
- Потеря внутренних жабр. Внешние жабры, существующие у многих водных и личиночных лиссамфибий — новые структуры и — не гомологичны с внутренними жабрами рыб.
Трудно определить точно, когда эти особенности появились, потому что они не сохранились в окаменелостях. Однако эти особенности не найдены у двоякодышащих рыб (самые близкие к четвероногам ныне живущие формы). Акантостега, девонский стегоцефал, все еще имела внутренние жабры, но ни у каких других стегоцефалов их, как известно, не было. Поэтому, внутренние жабры были, вероятно, потеряны рано в эволюции стегоцефалов, в девоне или миссиссиппии (приблизительно 360 миллионов лет назад), и никакой четвероног не имел когда-либо внутренние жабры.

Четвероноги девона и раннего миссиссипия вели водный и полуводный образ жизни, а первые полностью наземные четвероноги развились лишь в позднем пенсильвании, через 100 млн. лет назад после возникновения первых четвероногов. Однако имеется несколько пробелов в ископаемой находке четвероногов, наибольший из них, пробел Ромера, относится к раннему карбону (Tournasian) и длится 20 миллионов лет — между последними девонскими четвероногами и известными с середины Visean.
Примечания
- Из-за ограничений MediaWiki в качестве вышестоящего таксона тетраподоморф указан инфратип челюстноротых (Gnathostomata) вместо клады дипнотетраподоморф (Dipnotetrapodomorpha), приведённого в источнике: Betancur-R. R. et al.
The tree of life and a new classification of bony fishes // PLOS Currents Tree of Life. — 2013. — doi:10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288. - Ихтиостега // Италия — Кваркуш. — М. : Советская энциклопедия, 1973. — (Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 11).
- ↑ 12Ichthyostega
(англ.) информация на сайте Paleobiology Database. (Проверено 16 мая 2020).
Недостающие звенья[править | править код]
| 1850 |
| 1900 |
| 1950 |
| 2002 |
| Прогресс в изучении эволюции человека. Увеличение числа известных науке видов гоминин по времени. Каждый вид изображен в виде прямоугольника, показывающего границы, в которых варьировал объём черепа, и место вида в палеонтологической летописи. Видно постепенное заполнение промежутков между видами. |
Многие переходные формы между живыми организмами пока не найдены. Обычно их называют недостающими звеньями. Редкость переходных форм объясняется неполнотой палеонтологической летописи. Сторонники теории прерывистого равновесия также указывают на быстроту эволюционных преобразований. Однако палеонтологи постоянно находят новые ископаемые, заполняя пробелы (например, в 2004 году был найден тиктаалик — ещё одно переходное звено между рыбами и земноводными).
Неполнота палеонтологической летописи[править | править код]
Далеко не каждая переходная форма существует в виде окаменелости из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фоссилизации, то есть перехода в окаменелое состояние. Для образования окаменелости необходимо, чтобы погибший организм был погребён под большим слоем осадочных пород. Из-за крайне медленной скорости осадконакопления на суше, чисто сухопутные виды редко переходят в окаменелое состояние и сохраняются. Кроме того, редко удаётся обнаружить виды, живущие в глубинах океана, из-за редкости поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, либо наземные виды, ведущие полуводный образ жизни, либо обитающие недалеко от береговой линии. Закономерности процессов захоронения останков живых существ изучает особая отрасль палеонтологии — тафономия.
Ископаемые переходные формы — это формы организмов, сочетающие признаки более древних и молодых групп организмов.
Семенные папоротники сочетали признаки папоротников и семенных растений (семена, сидящие на безлистых осях или листоподобных органах, имели мегаспорангий — семязачаток; прототип стробилы; эустелу; окаймленные поры на трахеидах).

Ихтиостеги — рыболягушки, подотряд вымерших земноводных (лабиринтодонтов), представляющие собой переходную форму между кистеперыми рыбами и амфибиями. Известны из верхнедевонских отложений Гренландии и Австралии. Ихтиостеги имели такие признаки рыб:
- Тело обтекаемой формы.
- Имелся рудиментарный хвостовой плавник.
- Органы боковой линии на черепе, как у рыб, заключены в костные каналы.
- Сходное строение позвонков.
- В черепе сохранились рудиментарные кости жаберных крышек.
- Ноздри, как у двоякодышаших рыб, смещены к краю рта.
Признаки амфибий: имеются плечевой, тазовый пояса и грудная клетка, в конечностях имеются сходные кости.
Зверообразные рептилии — звероящеры, вымершие рептилии (синапсиды, тероморфы), сочетали признаки рептилий и зверей. Они были широко распространены в перми и триасе. Обособились от котилозавров в карбоне. В верхнем карбоне и перми преобладали примитивные звероящеры (пеликозавры). В верхней перми — триасе — преобладали терапсиды.
Звероящеры имели такие признаки млекопитающих:
- одна височная ямка, ограниченная снизу скуловой дугой
- нёбо вторичное костное
- клыки хорошо развиты
- нёбные зубы
- развита большая зубная кость
- нижняя челюсть подвешена непосредственно к черепу, а не при помощи квадратной кости, как у рептилий.
Это были хищники. Их потомки — териодонты. Обнаружены в Европе, Северной Америке, Африке.
Археоптерикс — промежуточная форма между птицами и рептилиями. Имела такие признаки птиц: тело покрыто перьями; маховые крылья крупные, асимметричные; кости конечностей частично полые; передние конечности — крылья, но с тремя пальцами.

Главными палеонтологическими доказательствами
эволюции считаются:
1) ископаемые остатки существ и растений, а также их отпечатки;
2) переходные формы;
3) филогенетические ряды.
Если с окаменевшими остатками и отпечатками (а также с редкими находками целых организмов, например, мамонтов, вмерзших в лед) все достаточно понятно, то переходные формы и филогенетические ряды следует рассмотреть подробнее.
Переходные формы
Переходными формами называют такие группы организмов, которые имеют одновременно признаки более древнего и более современного вида.
1. Псилофиты (риниофиты) — промежуточная форма между водорослями и папоротникообразными.
2. Семенные папоротники — между голосеменными и папоротниками.
3. Ихтиостега — между пресноводными земноводными и кистеперыми рыбами.
4. Сеймурия — между земноводными и рептилиями.
5. Группа зверозубых ящеров — переходная форма от рептилий к млекопитающим (найдена в районе бассейна Северной Двины). Эти ящеры имеют значительное сходство с млекопитающими в строении черепа, позвоночника, конечностей, их зубы делятся на клыки, резцы и коренные.
6. Археоптерикс — совмещает признаки птиц и рептилий. Его нередко до сих пор называют переходной формой, и даже в тестах ЕГЭ требуется выбирать именно археоптерикса.
7. Однако, по мнению многих специалистов, переходной формой между рептилиями и птицами является протоавис. Он имел киль и поэтому летал. Для него характерны полые кости.
Признаки птиц у археоптерикса:
1) задние конечности с цевкой;
2) наличие перьев;
3) общий вид.
Признаки пресмыкающихся у археоптерикса:
1) длинный ряд хвостовых позвонков;
2) брюшные ребра;
3) наличие зубов;
4) грудная кость без киля, слабо развитые грудные мышцы и мышцы крыльев.
Филогенетические ряды
Филогенетическими рядами называются ряды видов, последовательно сменяющихся от одного к другому в процессе эволюции. Например, «отец» эволюционной палеонтологии В.О. Ковалевский в своих научных трудах первым воссоздал филогенетический ряд лошади.
1. Из-за быстрого бега на длинные расстояния в высоком темпе у лошадей удлинялись конечности, уменьшалось число пальцев на них, постепенно формировалось роговое копыто.
2. Значительно увеличивались размеры животных (изначально предки лошадей предположительно были не больше лисы).
3. Указанные изменения были связаны с тем, что лошади поменяли ареал обитания — из тропических лесов, где жили их самые древние предки, лошади переместились в степи и вынуждены были пробегать длинные дистанции в поисках пищи.
4. Кроме перечисленных признаков для лошадей в процессе эволюции были характерны постепенный переход к стадному образу жизни, череп стал более крупным, челюсти массивными. Произошли изменения в строении зубов (выросли их размеры и поверхность). Увеличилась длина кишечника.
Вопрос в ЕГЭ по биологии, относящиеся к данной теме. Что представляет собой ряд предков современной лошади? Укажите не менее трех изменений в конечности лошади, произошедших в ходе эволюции.
1. Последовательный ряд предков — филогенетический ряд лошади.
2. В процессе эволюции у лошадей постепенно уменьшалось количество пальцев.
3. Удлинились конечности лошади, сформировалось роговое копыто.
На сегодняшний день выделяют три основных эмбриологических доказательства эволюции: закон зародышевого сходства, биогенетический закон, развитие всех организмов из одной клетки — зиготы. В ЕГЭ по биологии был вопрос, где надо было пояснить эти три доказательства.
Лондонский экземпляр археоптерикса, переходного звена между рептилиями и птицами.
Переходная форма
— организм с промежуточным состоянием, обязательно существующим при состоявшемся постепенном переходе от одного биологического типа строения к другому. Переходные формы характеризуются наличием более древних и примитивных (в значении первичных) черт, чем их более поздние родственники, но, в то же время, наличием более прогрессивных (в значении более поздних) черт, чем их предки. Как правило, говоря о промежуточных формах, имеют в виду ископаемые виды, хотя промежуточные виды вовсе не должны непременно вымирать. Известно множество переходных форм, иллюстрирующих происхождение четвероногих от рыб, рептилий от земноводных, птиц от динозавров, млекопитающих от териодонтов, китообразных от наземных млекопитающих, лошадей от пятипалого предка и человека от древних гоминид[1].
Теория эволюции
Ко времени написания Чарльзом Дарвином «Происхождения видов» науке не были известны переходные формы жизни. Этот факт Дарвин считал одним из самых слабых мест своей теории. Однако уже через два года был обнаружен археоптерикс — переходное звено между рептилиями и птицами.
Переходные формы между двумя группами организмов не обязательно являются потомками одной группы и предками другой. По ископаемым останкам, как правило, невозможно достоверно установить, является ли один организм прямым предком другого. Кроме того, вероятность найти в палеонтологической летописи чьего-нибудь прямого предка очень мала, и по умолчанию среди биологов принято такую возможность вообще не рассматривать. Гораздо выше вероятность обнаружить относительно близких родственников этого предка, схожих с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не «участок ствола».[2]
История изучения[ | ]
В 1932 году Сёве-Седерберг описал четыре вида ихтиостег из верхнего девона, найденных в восточной части Гренландии. Эти виды могли быть синонимичными, потому что их морфологические различия не были резко выражены. Виды различались в пропорциях и строении черепа. Сравнение было проведено на четырнадцати экземплярах, собранных в 1931 году Датской Восточной Гренландской экспедицией. Дополнительные экземпляры были собраны между 1933 и 1955 годами.
Род ихтиостег находится в родстве с акантостегой (Acanthostega gunnari
), также обнаруженной в восточной Гренландии. По сравнению с акантостегой череп ихтиостеги выглядит более рыбообразным, однако её пояс передних конечностей сильнее и лучше адаптирован к передвижению.
Примеры

Переходные формы в эволюции человека
В настоящее время известно множество переходных форм между человеком разумным и его обезьяноподобными предками. Так, например, открыты останки сахелантропа, ардипитека, австралопитеков (африканского, афарского и других), человека умелого, человека работающего, человека прямоходящего, человека-предшественника, гейдельбергского человека, неандертальца и раннего человека разумного.
Переходные формы в эволюции человека
В наше время найдено большое количество ископаемых останков, которые раскрывают эволюционный путь человека разумного от ее человекообразных предков. К формам, которые в большей или меньшей степени можно отнести к переходным, относятся: сахелантропа, ардипитека, австралопитеки (африканский, афарский и другие), человек умелый, человек работающий, человек прямоходящий, человек-предшественник, гейдельбергский человек и кроманьонцы.
Среди упомянутых форм значительное внимание заслуживают австралопитеки. Австралопитек афарский с точки зрения эволюции находится между современными двуногими людьми и их четвероногими древними предками. Большое количество рис скелета этого австралопитека четко отражают двуногость, причем до такой степени, что некоторые исследователи считают, что это свойство возникла задолго до появления австралопитека афарского. Среди общих черт анатомии, его таз гораздо больше похож на этих костей у человека, чем у обезьян. Края подвздошных костей короче и шире, крестцовая кость широкая и расположена непосредственно позади тазобедренного сустава. Существует явное свидетельство о существовании мест крепления для мышц-разгибателей колена, предусматривает вертикальное положение этого организма. В то время, как таз австралопитека не совсем как у человека (заметно шире, с ориентацией края подвздошных костей наружу), эти особенности указывают на принципиальную перестройку, связанную с хождением на двух ногах. Бедренная кость образует угол в направлении колена. Эта черта позволяет ноге размещаться ближе к средней линии тела и является явным свидетельством привычный характер передвижения на двух ногах. В наше время человек разумный, орангутаны и коаты имеют такие же черты. Ноги австралопитека имели большие пальцы, что делает практически невозможным захват стопой ветвей деревьев. Кроме особенностей локомоции, в австралопитека был также значительно больше мозг, чем у современных шимпанзе и зубы были значительно больше подобными зубов современного человека, чем к обезьянам.
Спор о постепенности эволюционных преобразований
Переходные формы являются объектом исследований в споре между градуалистами и пунктуалистами. Градуалисты считают, что эволюция является постепенным, непрерывным процессом. Пунктуалисты, или сторонники теории прерывистого равновесия считают, что виды в течение долгого времени остаются неизменными, а изменения происходят в очень короткий промежуток времени. Переходные формы являются аргументом обеих сторон. Пунктуалисты утверждают, что большинство видов появляются внезапно и на протяжении всей своей истории меняются слабо. Градуалисты отмечают, что среди позвоночных нет ни одного бесспорного примера очень быстрого перехода от одного вида к другому, большинство переходов вообще плохо задокументированы ископаемыми. Также много хорошо описанных внутривидовых изменений. Например, описаны изменения среднекайнозойских кроликов, миоценовых грызунов, гиппарионов, эоценовых парнокопытных, переход между родами эоценовых приматов.[3]
Недостающие звенья
| 1850 |
| 1900 |
| 1950 |
| 2002 |
| Прогресс в изучении эволюции человека. Увеличение числа известных науке видов гоминин по времени. Каждый вид изображен в виде прямоугольника, показывающего границы, в которых варьировал объём черепа, и место вида в палеонтологической летописи. Видно постепенное заполнение промежутков между видами. |
Многие переходные формы между живыми организмами пока не найдены. Обычно их называют недостающими звеньями. Редкость переходных форм объясняется неполнотой палеонтологической летописи. Сторонники теории прерывистого равновесия также указывают на быстроту эволюционных преобразований. Однако палеонтологи постоянно находят новые ископаемые, заполняя пробелы (например, в 2004 году был найден тиктаалик — ещё одно переходное звено между рыбами и земноводными).
Неполнота палеонтологической летописи
Далеко не каждая переходная форма существует в виде окаменелости из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фоссилизации, то есть перехода в окаменелое состояние. Для образования окаменелости необходимо, чтобы погибший организм был погребён под большим слоем осадочных пород. Из-за крайне медленной скорости осадконакопления на суше, чисто сухопутные виды редко переходят в окаменелое состояние и сохраняются. Кроме того, редко удаётся обнаружить виды, живущие в глубинах океана, из-за редкости поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, либо наземные виды, ведущие полуводный образ жизни, либо обитающие недалеко от береговой линии. Закономерности процессов захоронения останков живых существ изучает особая отрасль палеонтологии — тафономия.
Ихтиостега

Использовала ли ихтиостега свои лапы для ходьбы по суше, доподлинно неизвестно, но то, что она обладала свойствами, которые присущи как водным, так и наземным существам, неоспоримый факт. То есть, она была одновременно и рыбой, и земным животным.
Ученые сходятся во мнении, что именно ихтиостега является первым древним земноводным, которое смогло выйти на сушу и приспособиться к новым условиям жизни, в то время как большая часть амфибий просто вымерла, по причине изменения климатических условий, приведших к высыханию водоемов. Хотя, в конечном итоге, для нее самой этот природный эксперимент окончился весьма плачевно.
И так, что же собой представляла ихтиостега?
Она относилась к числу земноводных, которые большую часть времени проводили в воде, и лишь изредка выходили на сушу, по которой передвигались с помощью лап, так же, как это делают современные ящерицы. Хотя, при дальнейшем исследовании было установлено, что быстро двигаться она не могла. Причина лежала в ее позвоночнике, который по своему строению напоминал позвоночник животных, но никак не земноводных. Если посмотреть на задние лапы ихтиостеги, то они больше напоминали ласты тюленя. Вот вам и еще одно препятствие для перемещения по суше. Получается, что нормально ходить она не могла, и вместо этого просто ползла, выбрасывая вперед передние конечности, и затем подтягивала все остальное туловище. Вот по этой причине она просто не смогла стать промежуточным звеном между кистеперыми рыбами и наземными позвоночными, хотя, и пыталась это сделать.
Так что же стало причиной ее появления?
Ископаемые останки ихтиостеги ученые находят в Гренландии. Их возраст датируется примерно 370 миллионов лет. Именно в это время на Земле началось глобальное потепление, что вызвало высыхание водоемов и появление буйной земной растительности. Влажные тропические леса изобиловали папоротниками, хвощами, грибами, плаунами. Для жизни и развития земноводных природа создала идеальные условия. Этим и могла воспользоваться ихтиостега, которая попыталась перебраться с воды на сушу. Хотя, как мы теперь понимаем, без воды она обходиться не могла. Это была ее стихия. Во-первых, она должны была опасаться пересыхания тела, а во-вторых, именно в воде происходило ее размножение. Самка делала кладку яиц прямо в воду, после чего, самец их оплодотворял. Вылупившиеся личинки также развивались в воде. Они дышали жабрами, которые затем трансформировались в легкие. Вот так происходило их превращение в амфибий.
Да, истиостега могла перемещаться по суше, что позволяло ей переходить из одного водоема в другой. Но делала она это довольно неуклюже, а поэтому далеко от воды не отходила. Хотя, высокая влажность тропического леса позволяла ей долго оставаться на суше, без риска погибнуть от пересыхания тела. Все это время она спокойно дышала воздухом.
Пищу, ихтиостега также добывала себе в воде. Она опускалась на дно, зарывалась в ил, оставляя на поверхности лишь свои глаза, и дожидалась, когда мимо нее будет проплывать жертва. Это могли быть: рыбы, ракообразные, моллюски, черви и т д. В окружающем пространстве ихтиостега ориентировалась с помощью своей боковой линии.
Эти материалы будут вам интересны:
Гибод
Геликоприон
Плакодерм
Амбулоцетус
Филогенетические ряды

Впервые были открыты В. О. Ковалевским, который показал, что современные однопалые копытные происходят от древних пятипалых мелких всеядных животных.
Характеристики
Ихтиостеги были около 1,5 метров в длину и имели по семь пальцев на задних ногах. Точное количество пальцев на передних лапах пока не установлено, но вероятно, что их было тоже семь. На хвосте у них был плавник «рыбьего» типа поддерживаемый невральная и гемальнимы дугами. Ноздри располагались у нижнего края челюстей. Слезная кость примыкает к ноздре, но не до глазницы. Мижвисочна кость отсутствует (это обстоятельство исключает возможность происхождения большинства темноспондилив от этого рода). Задньотимьяна кость нечетная. Челюстная кость сталкивается с квадратновилицевою. Сохраняются под крышкой и передкришкови кости. Носовые кости широкие. Глазницы овальные и расположены в центральной части черепа. Хорда через Словно-окципитальну часть мозговой коробки доходит до гипофизарной ямы.