В предыдущей статье мы обсудили гипоталамус и гипофиз, которые теснейшим образом связаны друг с другом. Гипоталамус выделяет
либерины и статины, регулирующие деятельность гипофиза. Сейчас мы подробнее познакомимся со строением гипофиза и гормонами,
которые он выделяет.
Гипофиз
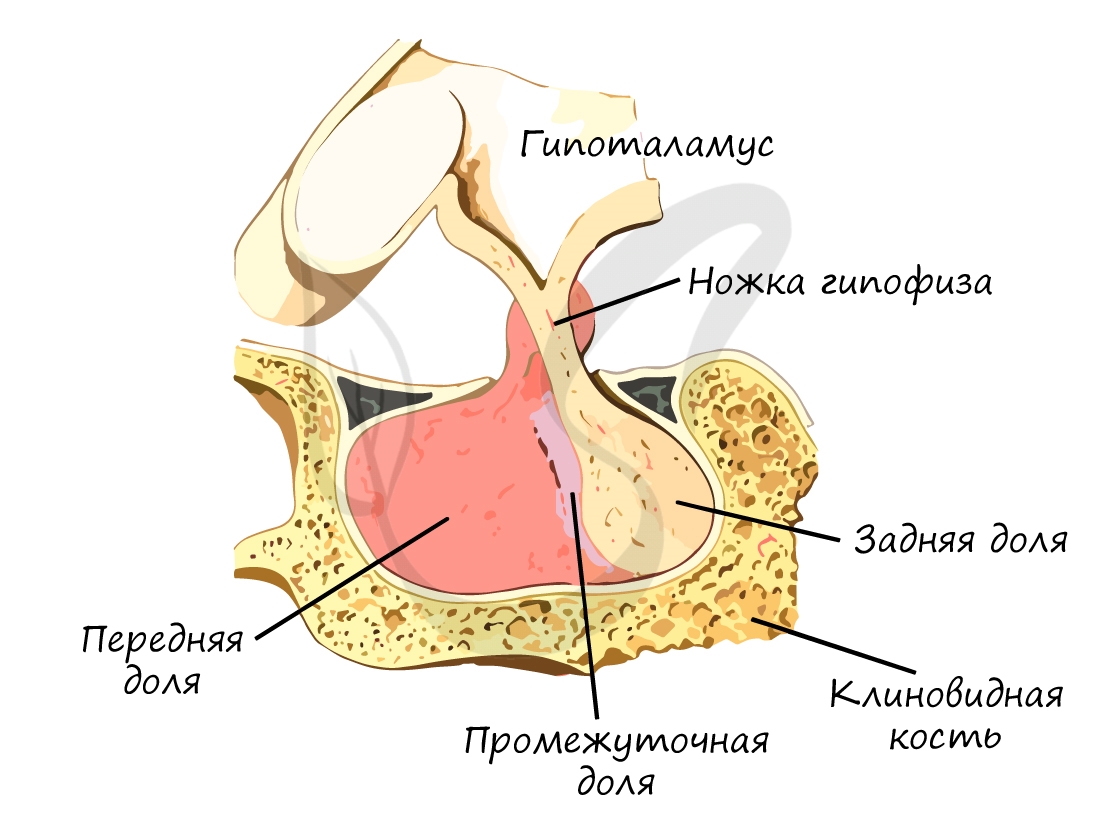
Гипофиз (нижний мозговой придаток, питуитарная железа) — железа внутренней секреции, расположенная в основании черепа. Состоит
из трех долей: передней, промежуточной (средней) и задней. Гипофиз называют «дирижером» желез внутренней секреции, так как его
гормоны влияют на их работу.
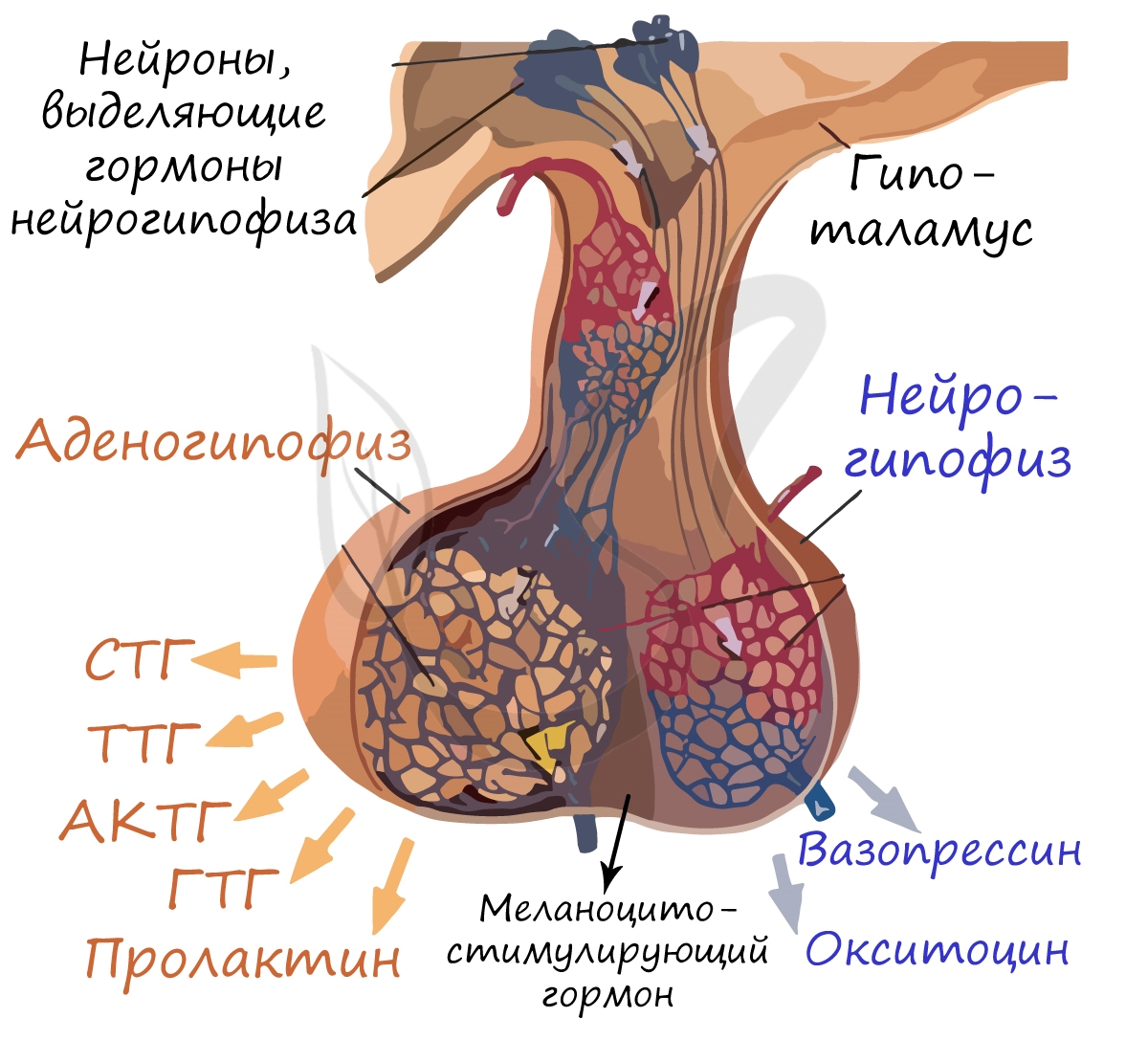
В передней части гипофиза (аденогипофиз) вырабатываются и выделяются в кровь тропные (от греч. tropos — направленность) гормоны:
- Тиреотропный гормон (ТТГ) — стимулирует выделение гормонов щитовидной железой (лат. glandula thyroidea — щитовидная железа)
- Адренокортикотропный (АКТГ) — стимулирует кору надпочечеников (от лат. adrenalis — надпочечный и лат. cortex — кора)
- Гонадотропный (ГТГ) — влияет на секрецию половыми железами половых гормонов и на созревание в половых железах яйцеклеток/сперматозоидов
(лат. gonas — половая железа) - Соматотропный (СТГ) — гормон роста, оказывает влияние на рост и развитие всех клеток организма (греч. soma — тело)
- Пролактин — стимулирует развитие молочных желез и образование в них молока у кормящих матерей
Особенное внимание обратим на гормон роста — СТГ. Нарушение его секреции приводит к тяжелым заболеваниям, так как он влияет на
рост и развитие организма. Секреция СТГ может быть повышена, в этом случае говорят о гиперфункции аденогипофиза (греч. hyper — над),
или снижена, в таком случае говорят о гипофункции аденогипофиза (греч. hypo — внизу). В детском и во взрослом возрасте последствия
гипо- и гиперфункции отличаются.
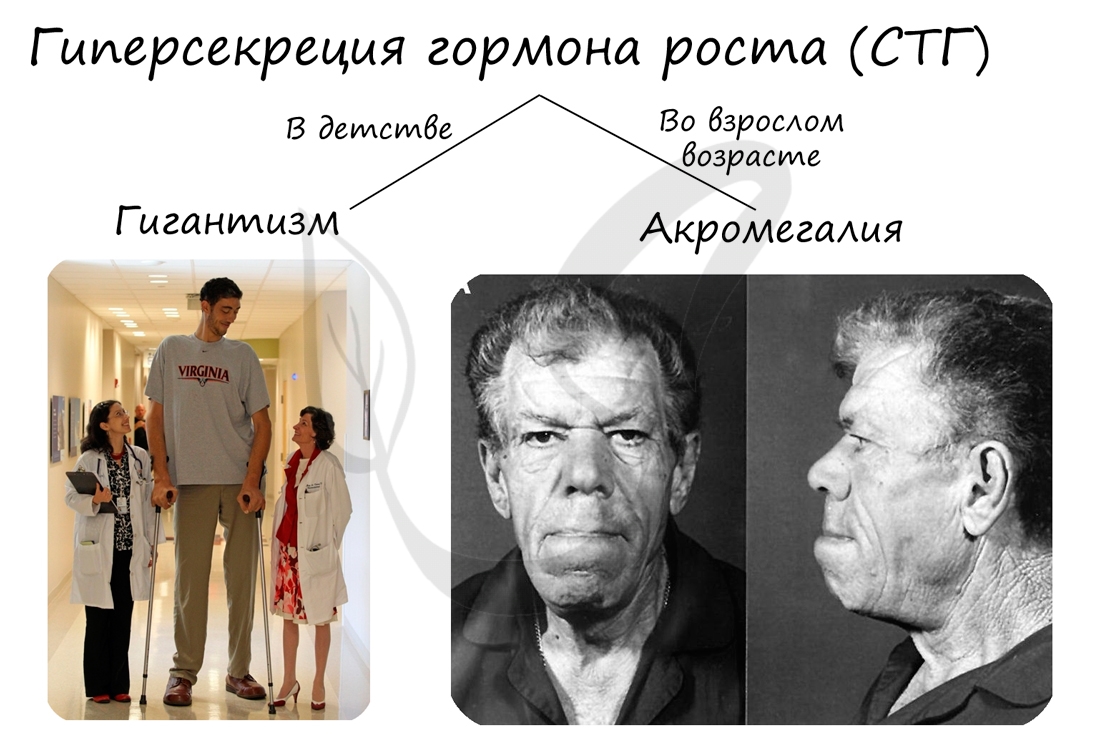
При гиперфункции аденогипофиза (СТГ повышен) в детском возрасте происходит избыточный рост костей и развивается гигантизм, пропорции тела при этом сохраняются. При гигантизме рост человека может достигать 2 и более метров. При такой патологии наиболее предрасположены к заболеваниям половые железы, суставы, нередко нарушается психика.
Во взрослом возрасте гиперфункция аденогипофиза не сопровождается увеличением роста, так как рост большей части костей окончен. Однако, начинают избыточно расти те кости, в которых есть хрящевая прослойка: фаланги пальцев, нижняя челюсть. Утолщаются губы и нос, увеличиваются
внутренние органы. Такое состояние во взрослом возрасте называется акромегалия (греч. akron — конечность и megas — большой).
При гипофункции аденогипофиза (снижена секреция СТГ) в детском возрасте развивается карликовость — задержка роста. При
карликовости тело имеет правильные пропорции, рост не более 1 метра, психика в норме. Это состояние может корректировать
врач, вовремя (в детстве!) назначив гормон роста в виде лекарства.
При гипофункции аденогипофиза во взрослом возрасте развивается изменение обмена веществ, что может привести как к
истощению, так и к ожирению.

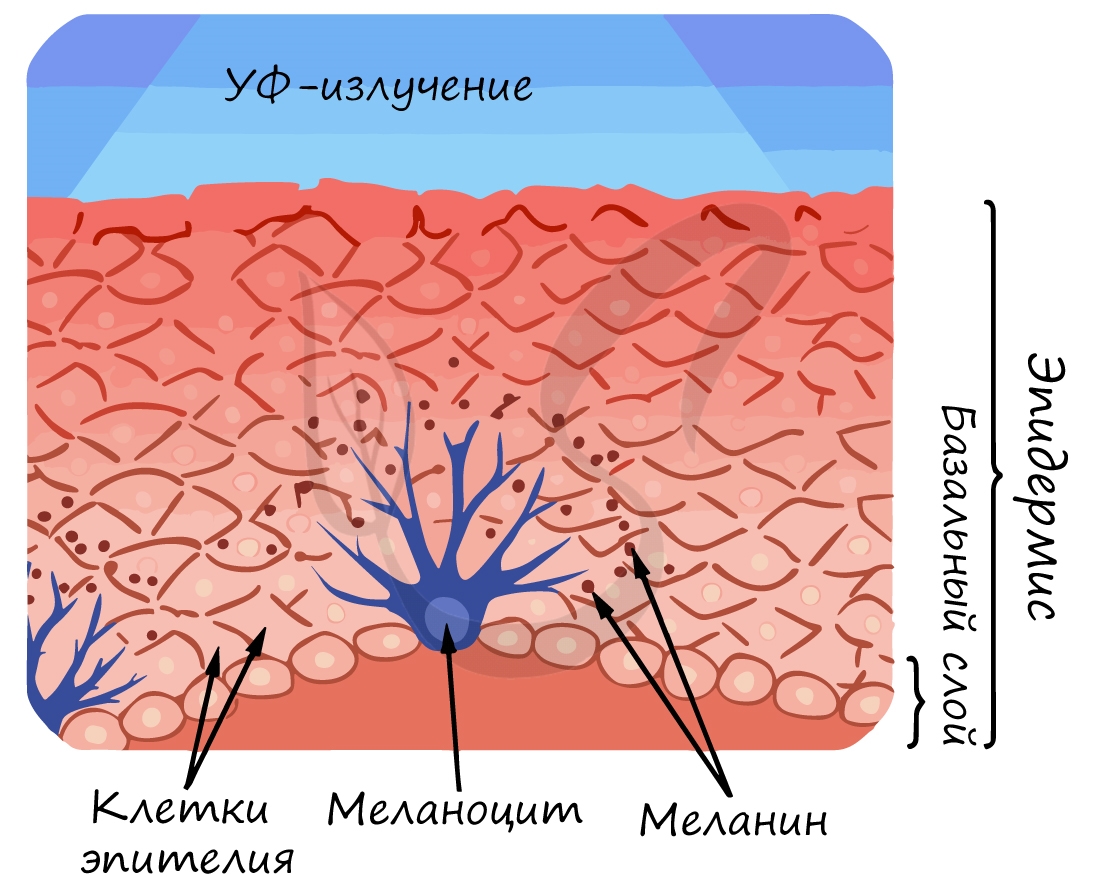
Промежуточная доля гипофиза синтезирует и выделяет меланотропный (меланоцитостимулирующий гормон). Вам уже известно, что меланоциты
располагаются в базальном слое эпидермиса, их пигмент — меланин, придает темную окраску коже. Меланотропный гормон стимулирует активность меланоцитов: они синтезируют меланин, пигментация кожи усиливается.
Задняя доля гипофиза — нейрогипофиз — не синтезирует (!), а только выделяет в кровь два гормона: вазопрессин (антидиуретический
гормон — АДГ) и окситоцин. Эти гормоны синтезируются нейронами гипоталамуса и по отросткам нейронов спускаются в нейрогипофиз,
где попадают в кровь.
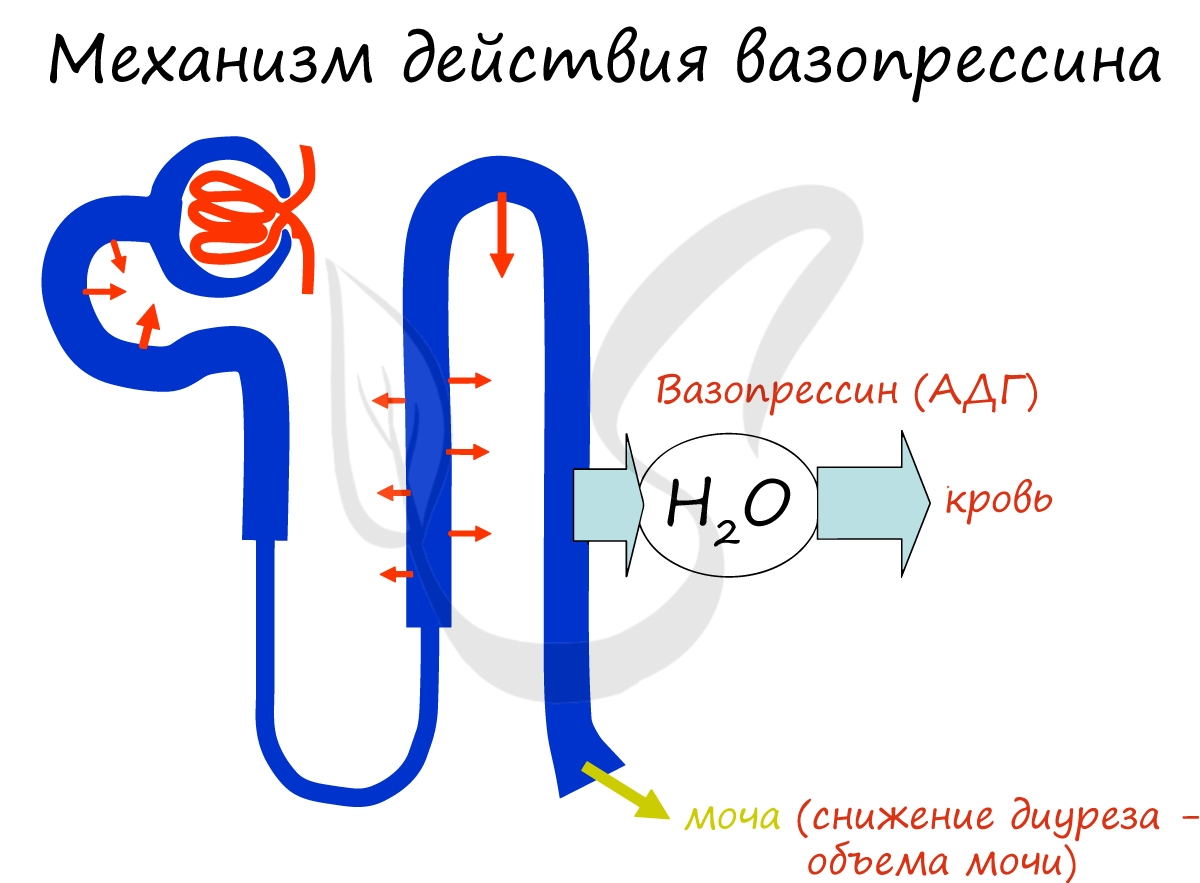
Вазопрессин усиливает реабсорбцию (всасывание) воды в канальцах нефрона, тем самым снижая выведение ее с мочой. При
нарушении секреции АДГ объем мочи может возрастать до 20 литров в сутки! Такое состояние носит название несахарный
диабет, так как подобно диабету характеризуется увеличением диуреза (объема мочи) и сильной жаждой.
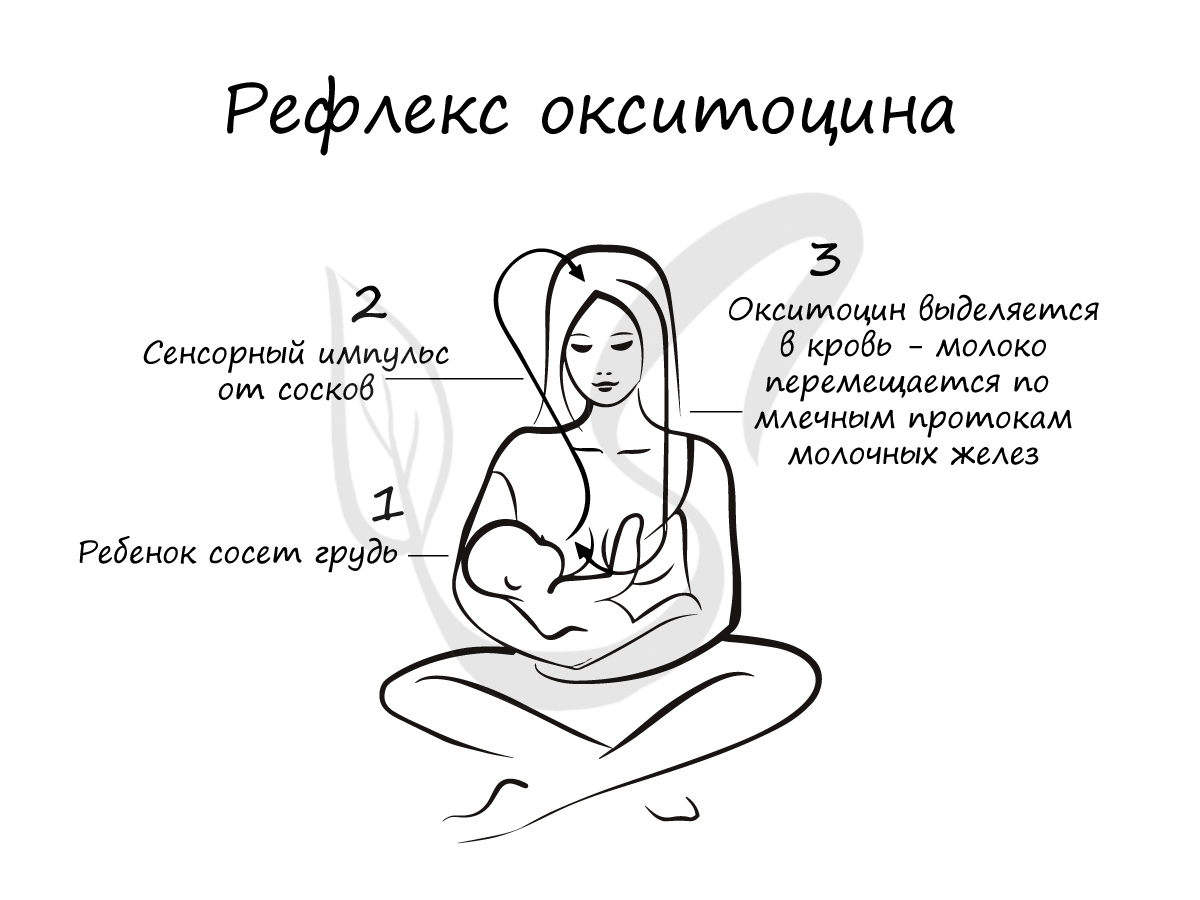
Окситоцин играет важную роль во время родов — он стимулирует сокращения матки, способствую продвижению плода по родовым
путям. У кормящих матерей окситоцин способствует лактации (секреции молока) в молочных железах при кормлении.
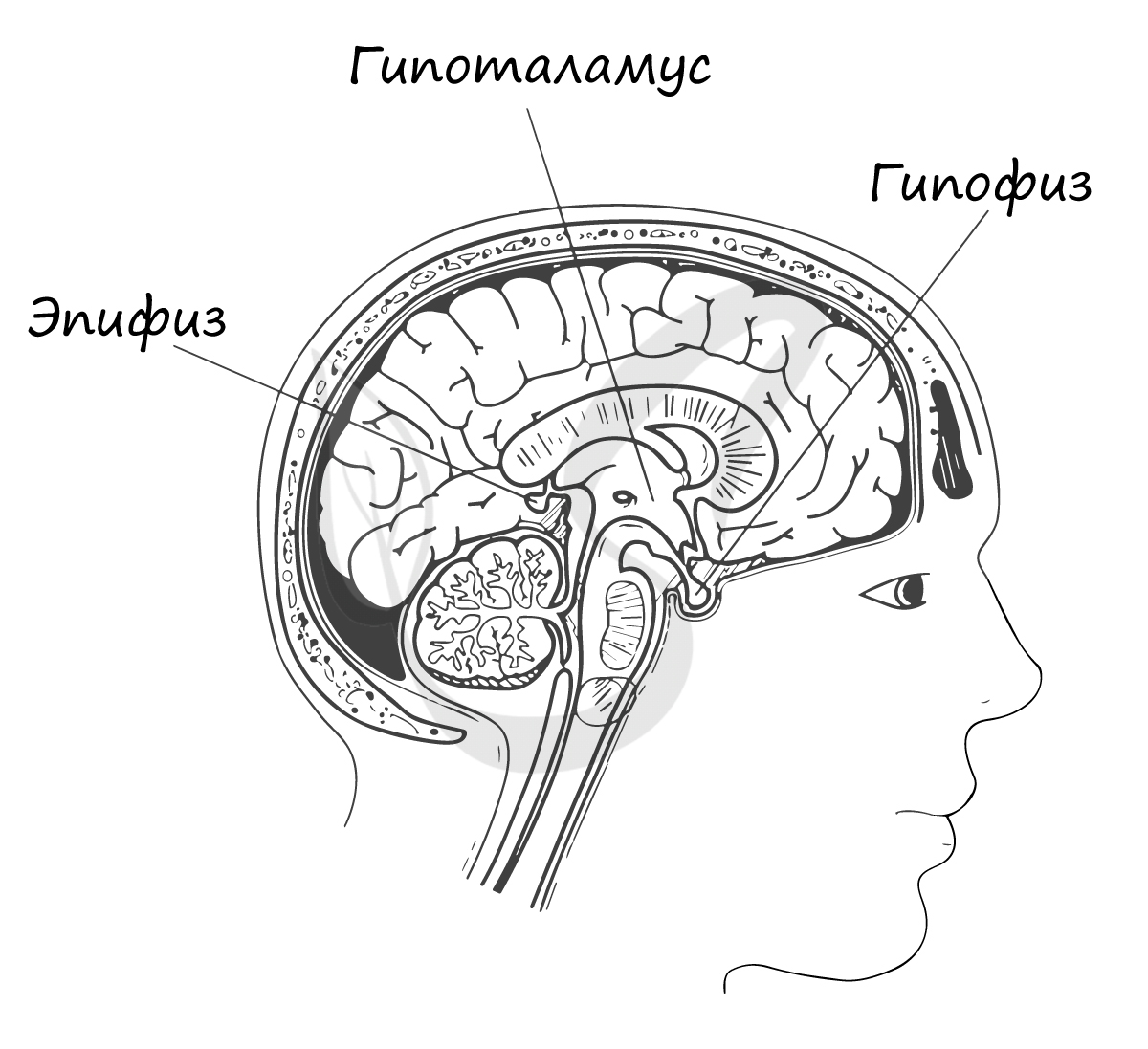
Эпифиз
Эпифиз (шишковидное тело) — эндокринная железа внутренней секреции, анатомически относящаяся к промежуточному мозгу.
В зависимости от освещенности, нейроны эпифиза синтезируют и выделяют гормон мелатонин, участвующий в регуляции
суточных и сезонных ритмов организма. Свет тормозит выработку мелатонина.
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Эндокринная система вместе с нервной составляет единое целое. Объединяющим и координирующим центром нейроэндокринной системы является область промежуточного мозга — гипоталамус, который не только проводит нервные импульсы, но и секретирует различные физиологические вещества [1].
Гипоталамус расположен на основании мозга и ограничен спереди перекрестом зрительных нервов, сзади — маммилярными телами, по бокам — расходящимися трактами зрительных нервов. Сверху в гипоталамическую область внедряется третий желудочек мозга, превращая гипоталамус в парный орган с симметричным расположением клеточных образований. В основании гипоталамуса располагается срединное возвышение, переходящее в ножку гипофиза.
Эндокринные функции гипоталамуса осуществляют преимущественно его передняя и медиальная части. Нервные клетки гипоталамуса группируются в многочисленные ядра, часть которых обладает секреторными свойствами. Крупноклеточные ядра переднего гипоталамуса (супраоптические и паравентрикулярные) связаны нервными волокнами с задней долей гипофиза. Многочисленные ядра медиобазального и части заднего гипоталамуса, часть паравентрикулярных, вентромедиальные и аркуатные — продуцируют нейросекрет в систему воротных вен гипофиза. Многочисленные комиссуральные пути гипоталамуса обеспечивают взаимодействие между нейроэндокринными ядрами. Нервные клетки паравентрикулярных и аркуатных ядер имеют многочисленные контакты с вышележащими структурами ЦНС.
Роль гипоталамуса в регуляции эндокринных функций была выяснена в 1955 г. Гиллемин впервые показал, что гипоталамический экстракт при введении его животным стимулирует секрецию гипофизом АКТГ. В 60-х годах XX века интенсивно исследовали влияние экстрактов гипоталамуса и срединного возвышения на секрецию гипофизом гормона роста (соматотропин, СТГ) и тиреотропного гормона (тиреотропин, ТТГ). Благодаря достижениям аналитической химии пептидов, разработке радиоиммунологических методов определения гормонов, развитию техники культивирования клеток сделан революционный скачок в исследованиях гипоталамической регуляции функций эндокринной системы.
Наличие в гипоталамусе специфических рецепторов к периферическим гормонам эндокринных желез и тропным гипофизарным гормонам определяет его особое положение в системах обратной связи. В гипоталамусе происходит переключение информации с нервно-медиаторного на нейрогормональный путь, которой составляют либерины и статины: кортиколиберин (АКТГ-рилизинг гормон, КРГ), люлиберин (гонадолиберин, ЛГ/ФСГ-рилизинг-гормон, ГнРГ), тиролиберин (ТРГ), соматолиберин (РГГР), соматостатин, меланолиберин, а также дофамин (ДА) и энкефалины, оказывающие влияние на функции гипофиза.
Гипоталамус является высшим вегетативным центром, осуществляющим сложную интеграцию и различных внутренних систем. В организме нет ни одной функции, к регуляции которой гипоталамус не имел бы отношения: он играет существенную роль в поддержании оптимального уровня обмена веществ (белкового, углеводного, жирового, водного и минерального) и энергии, в регуляции температурного баланса организма, деятельности пищеварительной, сердечно-сосудистой, выделительной, дыхательной и эндокринной систем.
Нейроэндокринологи имеют полное право рассматривать гипоталамус как главный нейроэндокринный орган, который осуществляет взаимосвязь между ЦНС и железами внутренней секреции. Он является интегрирующим звеном, пультом управления, на котором замыкаются все пути между ЦНС и эндокринной системой. Естественно, возникает вопрос, каким образом маленькая структура промежуточного мозга справляется со столь многочисленными функциями.
Для эндокринологов, в первую очередь, важен вопрос о регуляции гормональной системы.
Связь между гипофизом, который является основным продуцентом всех тропных гормонов (АКТГ, ТТГ, ЛГ, ФСГ, СТГ, пролактин) и гипоталамусом осуществляется через портальную систему сосудов, а также через гипоталамо-гипофизарный тракт — самый короткий, но хорошо очерченный пучок аксонов нейронов гипоталамуса. Волокна их берут начало в супраоптическом и паравентрикулярных ядрах и проходят через гипофизарную ножку к нейрогипофизу.
Биохимиками было показано существование в гипоталамусе специфических веществ, которые синтезируются нервными клетками и освобождаются в капилляры портальной системы, достигая передней доли гипофиза. На сегодняшний день выделено и охарактеризовано более 10 гипоталамических нейрогормонов, каждый из которых обладает специфическим влиянием (стимуляцией или торможением) секреции того или иного тропного гормона гипофиза. Такие соединения получили название ингибирующих и рилизинг-гормонов (либерины и статины), обладающие высокой специфичностью действия и чрезвычайно высокой биологической активностью. Установлена и подтверждена химическим синтезом их структура.
Основной точкой приложения гипоталамических рилизинг-гормонов являются клетки гипофиза, и некоторое время считалось, что каждый гормон стимулирует один тип клеток аденогипофиза, который в свою очередь синтезирует лишь один тропный гормон.
В дальнейшем было показано, что в ряде случаев гипоталамические гормоны стимулируют секрецию нескольких гипофизарных гормонов. Например, ТРГ усиливает выделение и ТТГ, и ПРЛ. ГнРГ способствует секреции ЛГ и ФСГ; соматостатин угнетает секрецию и гормона роста, и ТТГ. Каждый клеточный элемент аденогипофиза подвергается действию многих регуляторных факторов, и клетки в аденогипофизе не строго специализированы: в одной клетке могут синтезироваться несколько гормонов, например ЛГ и ФСГ, АКТГ и β-липотропин, СТГ и ПРЛ, но не ТТГ или ЛГ. Бифункциональность некоторых гормонов объясняется по-разному. Способность ГнРГ вызывать секрецию как ЛГ, так и ФСГ, причем несовпадающую по времени, определяется разной чувствительностью гонадотрофов, секретирующих ЛГ или ФСГ, а также длительностью воздействия с учетом влияния половых стероидов на гипофизарные клетки в разные стадии полового цикла. Большие концентрации эстрогенов в плазме, предшествующие овуляторному выбросу ЛГ, увеличивают чувствительность гипофиза к ГнРГ, а минимальные количества эстрогенов и прогестерона, секретируемого после овуляции, напротив, снижают эту чувствительность. Секреция ФСГ более чувствительна к ингибирующему эффекту эстрогенов. Нельзя исключить наличие в аденогипофизе и двух видов рецепторов. Одни из них характеризуются высоким сродством к ГнРГ и низкой связывающей способностью; за их счет осуществляется быстрая секреция ЛГ; вторые — более низким сродством к ГнРГ, но большей силой связывания, что и определяет более медленное освобождение ФСГ. Возможно существование двух различных нейрогормонов — ЛГ-РГ и ФСГ-РГ.
Анатомия и физиология гипофиза
Гипофиз расположен в турецком седле в основании черепа. Сверху он отделен от мозга выростом твердой мозговой оболочки. Гипофиз состоит из трех долей: передней, задней и средней (промежуточной). Последняя, как правило, анатомически не обособлена и вместе с передней входит в состав аденогипофиза, который составляет около 80% массы железы; задняя доля — нейрогипофиз — анатомически и функционально объединена со срединным возвышением, являющимся частью гипоталамуса. Аденогипофиз образуется из задней стенки ротовой полости зародыша (карман Ратке), задняя доля — из дивертикула дна третьего желудочка мозга.
Структурно-функциональная связь гипофиза с гипоталамусом осуществляется нервным и гуморальным путем. Нервные волокна из крупноклеточных гипофизотропных ядер гипоталамуса (паравентрикулярного и супраоптического) через срединное возвышение и ножку гипофиза достигают задней его доли. По аксонам этих клеточных ядер в заднюю долю гипофиза поступают окситоцин и вазопрессин. На стенках капилляров срединного возвышения оканчиваются терминали пептидергических волокон. Аксоны гипоталамических нейронов, идущие к аденогипофизу, прерываются на уровне срединного возвышения, где и происходит секреция гипоталамических статинов и либеринов.
Нейрогормоны попадают в переднюю долю гипофиза с кровотоком по системе воротных вен гипофиза. Стенки каппиляров, образуемых ветвями верхних гипофизарных артерий в срединном возвышении, проницаемы для рилизинг-гормонов. Концентрация последних в крови воротных вен гипофиза во много раз выше, чем в периферической крови.
Кровоток через переднюю долю гипофиза значительнее, чем через любой другой орган, что, наряду с высоким содержанием нейрогормонов в крови сосудов гипофиза, создает оптимальные условия для регуляции функций аденогипофиза и зависимых от него эндокринных желез. Задняя доля гипофиза снабжается кровью нижних гипофизарных артерий.
Являясь анатомически целостным органом, гипофиз образован двумя онтогенетически разными образованиями: передним (железистым) и задним (нервным). Клетки аденогипофиза разделяются по своей гистологической окраске на базофилы, ацидофилы (эозинофилы), хромофобы. Однако выявление специализированных клеток—лактотрофов, тиреотрофов, соматотрофов, кортикотрофов, гонадотрофов, продуцирующих соответствующие гормоны, осуществляется иммуноцитохимическим методом, выявляющим специализацию клеток с помощью высокоспецифических антител к конкретному гормону. Большую часть популяции аденогипофизоцитов составляют лактотрофы и соматотрофы. Обнаруживаются и камбиальные элементы, способные при необходимости дифференцироваться в те или иные специализированные клетки.
Гипофиз как центральный орган эндокринной системы получает афферентную и эфферентную информацию. Из срединного возвышения по сосудам портальной системы в аденогипофиз поступает кровь, содержащая гипоталамические нейрогормоны. К гипофизу поступает также большое количество гормонов периферических эндокринных желез и биологически активных метаболитов. Переработка этой информации позволяет гипофизу осуществлять тонкую интегральную функцию и посылать периферическим эндокринным железам адекватные, строго дозированные регуляторные сигналы путем выделения в кровь тропных гормонов. Периферические эндокринные железы, реагируя на тропные гормоны гипофиза, секретируют в кровь собственные гормоны, регулирующие биохимические процессы в тканях-мишенях. Циркулирующие в крови гормоны и биологически активные метаболиты контактируют со всеми структурами нейроэндокринной системы: ЦНС, гипоталамусом, гипофизом, другими железами внутренней секреции, клетками АПУД-системы (которые присутствуют в различных органах и тканях и продуцируют полипептидные гормоны за счет поглощения и декарбоксилирования предшественников аминов).
Главным в механизме действия гипоталамических гормонов является их избирательное взаимодействие с рецепторами, локализованными на плазматических мембранах аденогипофизарных клеток; конформационные изменения гормон-рецепторных комплексов приводят к изменению активности ферментных систем клеточных мембран. Посредником в дальнейшей передаче гормонального сигнала, направленного на секрецию и синтез аденогипофизарных гормонов, является цАМФ, которым активирует цАМФ-зависимую протеинкиназу. Изменение проницаемости плазматических мембран, происходящее под влиянием рилизинг-гормонов, способствует проникновению в клетку экзогенного Са2+, необходимого для секреции тропных гормонов.
Вопрос и о локализации биосинтеза гипоталамических гормонов окончательно не решен. Имеются данные об их образовании в нервных окончаниях — синаптосомах, так как именно в них обнаруживается скопление этих гормонов и биогенных аминов. В настоящее время биогенным аминам отводится главная роль в регуляции секреции и синтеза гипоталамических гормонов. В гипоталамусе присутствуют специфические рецепторы гормонов и периферических аминорецепторов.
Воспринимая регуляторные сигналы сверху, железа ускоряет или тормозит выделение собственного гормона и, более того, она получает информацию о результатах действия ее гормонов на периферии. Механизм такой организации эндокринной железы, называемой в литературе принципом обратной связи (положительной и отрицательной), был впервые сформулирован М.М. Завадовским (1935). Он обеспечивает надежность работы нейроэндокринной системы в организме. Классическим примером может служить функционирование щитовидной железы, когда ТТГ стимулирует секрецию тиреоидных гормонов, которые в свою очередь, достигнув высоких концентраций в крови, тормозят секрецию гипофизом ТТГ. Обратная связь реализуется на четырех уровнях (ЦНС, гипоталамус, гипофиз, сама железа), и в ней различают длинные, короткие и ультракороткие звенья. Эндокринные регуляторы — гормоны дистантного действия — достигают своих целевых объектов с кровотоком.
Концентрация и ритм секреции гормонов определяют направленность действия и точку приложения в организме. Так, эстрогены, стимулируя секрецию пролактина гипофизом, увеличивают выделение ДА гипоталамусом, что приводит к торможению лактотропной функции гипофиза. На телах нейронов, синтезирующих ДА, обнаружены рецепторы эстрадиола. В этом процессе участвуют и метаболические регуляторы, такие как глюкоза [10].
В одной и той же железе имеется возможность обмена химической информацией между клетками, в которой принимают участие и гормоны. Показано, в частности, наличие рецепторов инсулина на альфа-клетках, секретирующих глюкогон (паракринная регуляция). Биологически активные вещества, в том числе гормоны, могут выделяться и связываться рецепторами той же клетки (аутокринная регуляция).
Чувствительность тканей к конкретному гормону определяется количеством специфических рецепторов, связывающих этот гормон. Высокие концентрации гормона, связывая большую часть рецепторов, снижают их биосинтез и уровень активных форм рецепторов, что предохраняет клетки-мишени от чрезмерного гормонального воздействия. Возможна и ситуация, когда гормон увеличивает концентрации собственных рецепторов.
Гормон может влиять не только на свои рецепторы, но и на рецепторы других гормонов. Так, ПРЛ стимулирует образование рецепторов ЛГ и ФСГ в гонадах, тиреоидные гормоны влияют на уровень эстрогенных и катехоламиновых рецепторов и т.д. Обратная связь реализуется довольно быстро, что обеспечивает постоянный относительный гомеостаз в организме; замедление или ускорение этого процесса свидетельствует о развитии патологического процесса.
Нарушения гормональной рецепции ведут к развитию патологического процесса (например, инсулинорезистентность при ожирении и сахарном диабете 2-го типа).
В срединном возвышении обнаружены многочисленные синаптические и аксональные контакты нейронов, вырабатывающих нейропептиды, а также анастомозы между капиллярами сосудов, идущими к аденогипофизу и нейрогипофизу, что и обеспечивает тесное взаимодействие различных эндокринных подсистем (гипоталамус—гипофиз—кора надпочечников, гипоталамус—гипофиз—щитовидная железа, гипоталамус—гипофиз—гонады и др.).
Главным достижением нейроэндокринологии последних лет является доказательство причастности нейромедиаторов к нейроэндокринной интеграции действия нейросекреторных нейронов, регулирующих секрецию тропных гормонов гипофиза. К ним относятся норадреналин (НА), адреналин (А), ДА, серотонин, гистамин, гамма-аминомасляная кислота (ГАМК), ацетилхолин, соматостатин, ТРГ, КРГ, соматолиберин и многие другие соединения. Все эти вещества несут нейрогенную информацию, необходимую для нормального функционирования нейросекреторных нейронов, регулирующих секрецию тропных гормонов гипофиза. Прежде всего это люлиберинпродуцирующие нейроны, регулирующие выделение ЛГ и ФСГ, нейроны, секретирующие пролактинингибирующий гормон, ТРГ, вазоактивный интестинальный полипептид, а также ДА. Полное понимание нейроэндокринного контроля требует точного знания топографии нейросекреторных и обычных проводниковых нейронов. Необходимо определить главные проекции нейронов и идентифицировать нейромедиаторы, содержащиеся в них. Обнаружены синапсы между норадренергическими нейронами и ГАМК-содержащими нейронами в этой области. Следует отметить также возможность несинаптического взаимодействия катехолсодержащих нейронов в ЦНС.
Дофаминергические нейроны, иннервирующие гипоталамус, имеют клеточные тела в трех областях мозга: во-первых, это тубероинфундибулярные нейроны, которые составляют 3—5% клеточных тел нейронов аркуатной области и обозначаются как А12. Терминали этих нейронов обнаружены в срединном возвышении в тесном контакте с первичными капиллярами гипофизарного портального сплетения. Эти нейроны выделяют большое количество ДА в портальную кровь и играют ведущую роль в регуляции секреции ПРЛ, а также ГнРГ. Две другие группы дофаминергических нейронов расположены в каудальном таламусе, заднем гипоталамусе и перивентрикулярных ядрах. Диффузное распределение дофаминергических нейронов в гипоталамусе затрудняет изучение их роли в регуляции секреции гонадотропинов и других тропных гормонов.
Изложение контрольных функций адренергических нейронов в секреции гонадотропинов целесообразно начать с их роли в регуляции пульсирующего выделения ЛГ, которая определяется квантовым выделением ГнРГ. Большинство исследователей, занимающихся этой проблемой, приходят к выводу, что норадренергические нейроны в физиологических условиях включаются в этот процесс, хотя на отдельных нейронах обнаружены адренергические рецепторы, активация которых вызывает торможение пульсирующего выделения ЛГ. Направленность действия ДА-нейронов зависит, скорее всего, от уровня половых гормонов.
При изучении роли биогенных аминов в регуляции овуляторного выброса гонадотропинов особое внимание обращается на значение половых гормонов в активации или торможении катехолсодержащих нейронов в системе обратной связи. На модели овариэктомированных крыс с имплантацией эстрадиола показано, что выброс ЛГ каждые 24 ч сопровождается увеличением скорости обмена НА в преоптической и аркуатной областях и супрахиазматических ядрах, а также срединном возвышении. Аналогичные данные получены и в опытах на интактных животных, у которых отмечено увеличение скорости обмена НА в преоптической области во 2-й половине проэструса, т.е. во время овуляторного выброса ЛГ, тогда как в другие стадии цикла подобных изменений в обмене НА в этой области не зарегистрировано. О негативном действии эстрадиола на обмен НА свидетельствуют эксперименты на овариэктомированных животных, у которых скорость обмена НА в преоптической области увеличивалась через 3 сут, а введение эстрадиола приводило к снижению скорости его обмена в этой области и срединном возвышении уже через 3 ч. Действие эстрогенов на обмен НА может осуществляться как непосредственно на уровне норадренергических нейронов, содержащих эстрогенные рецепторы, так и через опиоидные или ГАМК-содержащие интернейроны, которые также содержат рецепторы стероидных гормонов. Скорость обмена адреналина в некоторых областях гипоталамуса также увеличивается одновременно с овуляторным выбросом ЛГ. Это касается, главным образом, медиобазального гипоталамуса и преоптической области. Возникает вопрос о степени сродства адренергических рецепторов нейронов к половым гормонам в ходе менструального цикла.
Влияние половых гормонов на активность дофаминергических нейронов проявляется менее отчетливо. Дело в том, что изменение состояния тубероинфундибулярных дофамин-содержащих нейронов теснее связано с секрецией ПРЛ. Введение эстрогенов повышает уровень ПРЛ, который в свою очередь усиливает обмен ДА в этих нейронах. Показано также наличие рецепторов к эстрадиолу в дофаминергических клеточных телах нейронов аркуатного ядра и повышение скорости обмена ДА в этом ядре и срединном возвышении во второй половине стадии проэструса. В преоптической области изменения дофаминергической активности в это время не наблюдается. Овариэктомия незначительно влияет на обмен ДА в некоторых областях гипоталамуса, тогда как введение эстрадиола вызывает снижение обмена ДА в преоптической области и срединном возвышении и увеличение в медиобазальном гипоталамусе.
Имеется ряд сообщений о влиянии катехоламинергических агентов на овуляторный выброс ЛГ в проэструсе и вызванный эстрогенами выброс ЛГ. Начало этим исследованиям было положено в 1950 г. Сойером, который показал, что α-адренергические антагонисты блокируют овуляцию у крыс. В дальнейшем были исследованы более специфические фармакологические агенты, такие как α-метил-р-тирозин и диэтилдитиокарбомат, блокирующие синтез НА и А соответственно. Они тормозили овуляторный выброс ЛГ. Блокада α-адренергических рецепторов также прерывает овуляцию, причем наиболее эффективным является препарат празозин, избирательно блокирующий подтип α1-рецепторов, тогда как блокада другого подтипа рецепторов (α2) пипероксаном неэффективна.
Приведенные данные позволяют утверждать, что норадренергические и адренергические нейроны включаются в стимуляцию овуляторного выброса ЛГ, и действие их опосредуется через α1-адренергические рецепторы, локализованные в основном в медиобазальной преоптической области гипоталамуса. Роль дофаминсодержащих нейронов в регуляции овуляторного выброса ЛГ спорна: введение относительно специфического антагониста ДА пимозина в утренние часы проэструса снижало выброс ЛГ у крыс во второй половине дня. Этот же препарат, введенный женщинам за 2 ч до ожидаемого выброса ЛГ в середине менструального цикла, снижает величину выброса. В другом случае внутрижелудочковое введение ДА в проэструсе у крыс способствует увеличению уровня ЛГ в крови. Исследования in vitro также дают спорные результаты. Так, перфузия ДА фрагментов медиобазального гипоталамуса самцов крыс увеличивает выделение ГнРГ, которое блокируется введением антагониста α-адренергических рецепторов (фентоламина), но не антагониста ДА (пимозина). ДА обладает способностью ускорять выделение НА из терминалей срединного возвышения, но не влияет на обратный захват и обмен НА. Противоречивые данные затрудняют интерпретацию роли и значения ДА в решении системы ГнРГ—ЛГ.
Серотонинергическая система ЦНС представляет собой популяцию нейронов ствола мозга, которые берут начало в ядрах шва среднего мозга. Аркуатные ядра получают плотную серотонинергическую иннервацию, тесно контактирующую с дофаминергическими клеточными телами аркуатного ядра и медиальной зоны инсепта. Описаны контакты между нейрональными элементами, содержащими ГнРГ и серотонин, в преоптической области, концевой пластинке и срединном возвышении. Серотонин обнаружен в нервных окончаниях, локализованных вокруг больших кровеносных сосудов аденогипофиза, а также в секреторных гранулах гонадотрофов. Причастность серотонина к регуляции гонадотропной функции гипофиза доказана многочисленными исследованиями. Он может как стимулировать, так и тормозить эту функцию в зависимости от гормонального фона. Действие серотонина на секрецию ЛГ и овуляцию опосредовано регуляцией секреции ГнРГ нейросекреторными нейронами. Серотонинергическая система является одной из многих нейросекреторных систем, тесно связанных с гонадотропинрегулирующей системой преоптикопереднего и медиобазального отделов гипоталамуса и срединного возвышения. Половые гормоны модулируют активность этой нейротрансмиттерной системы. Анализ роли нейромедиаторов адренергического ряда в регуляции гонадотропной функции гипофиза позволяет утверждать, что НА следует рассматривать в качестве ведущего агента в регуляции секреции ЛГ, основной точкой приложения которого является преоптическая область. ДА реализует свой эффект через аркуатную область гипоталамуса, а серотонин играет роль синхронизирующего агента.
Новый класс соединений, выделяемых в группу пептидных нейромедиаторов, вносит свой вклад в наши представления об участии нервной системы в регуляции эндокринных функций гипофиза. Обнаружение регуляторных пептидов, общих как для нервной, так и для эндокринной систем, вызвало революцию в наших представлениях. Возникло новое направление исследований — психонейроэндокринология.
Практически каждый из перечисленных выше нейропептидов причастен к регуляции секреции гонадотропинов, ПРА, АКТГ, ТТГ. Большинство из них обнаружено в медиобазальном гипоталамусе. Нейропептиды, выделяясь в гипофизарную портальную систему, действуют на несколько типов аденогипофизарных клеток и, кроме того, взаимодействуют с многими нейромедиаторами непептидной природы, поступающими в медиобазальный гипоталамус из других структур, что в свою очередь обеспечивает их прямое или опосредованное действие на гормональный контроль.
Гипоталамические нейроны, продуцирующие нейропептиды, составляют общую систему. Большинство их клеточных тел локализовано в паравентрикулярных ядрах, медиальной перивентрикулярной области и аркуатных ядрах. Незначительная часть таких нейронов обнаружена в преоптической области и супрахиазматических ядрах. Бóльшая часть всех этих нейронов заканчивается в области срединного возвышения. Аксональные коллатерали иннервируют ряд гипоталамических и экстрагипоталамических структур. Нервные клетки паравентрикулярных и аркуатных ядер в свою очередь имеют многочисленные контакты с большим числом других структур ЦНС, что и определяет их стратегическую позицию в координации гормональной регуляции, а также ряда поведенческих реакций. Особенно это касается полового поведения, требующего синхронизации поведенческих компонентов с процессами менструального цикла. Окончания нейронов, синтезирующих нейропептиды, тесно контактируют не только с перикарионами, вырабатывающими нейротрансмиттеры, но и с нейронами, синтезирующими нейропептиды. Например, ГнРГ-продуцирующие нейроны и их волокна могут оканчиваться на аналогичных клеточных телах; имеют место также аксо-аксональные контакты.
Учитывая большое число нейропептидов, принимающих участие в регуляции репродуктивной системы, целесообразно сгруппировать их по месту синтеза. Такими группами могут быть: 1) нейропептиды, продуцируемые нейронами, локализованными, главным образом, в преоптической области (соматостатин, ГнРГ, тахикинины, нейротензин, предсердный натрийуретический гормон); 2) нейропептиды, продуцируемые нейронами аркуатной области (панкреатический нейропептид, пептид YY, нейропептид Y); 3) нейропептиды, синтезирующиеся в перикарионах паравентрикулярных или супраоптических ядер (ТРГ, КРГ, холецистокинин, вазопрессин, окситоцин, ангиотензин, брадикинин, бомбезин); 4) гастроинтестинальные пептиды (ВИП, гастроинтестинальный пептид); 5) семейство опиоидных пептидов.
Механизм действия этих нейропептидов как на гипоталамическом, так и на гипофизарном уровне можно проследить на примере модуляции секреции гонадотропинов, ПРЛ, окситоцина, т.е. тех гормонов, которые имеют отношение к репродуктивной функции. Например, соматостатин, который обнаружен во многих структурах ЦНС и главным действием которого является угнетение продукции гормона роста и частично ТТГ и ПРЛ, способен изменять активность нейромедиаторных систем мозга, вызывая блокаду секреции ГнРГ.
Группа классических пептидов (тахикинины А и Б, субстанция Р) стимулирует выделение ПРЛ и тормозит выделение гормона роста. Пептид нейротензин широко распространен в ЦНС и проявляет свой эффект за счет стимуляции секреции инсулина, ингибиции секреции глюкагона и модуляции гастроинтестинальной перистальтики. Внутрижелудочковое введение нейротензина снижает уровень ЛГ в крови, что делает возможным его участие в контроле гонадотропной функции гипофиза. Предсердный натрийуретический гормон угнетает выделение вазопрессина и может стимулировать выделение ЛГ и пролонгировать действие ГнРГ. Семейство панкреатических пептидов также причастно к функционированию репродуктивной системы, они чаще всего блокируют секрецию ЛГ и снижают частоту пульсации ГнРГ в срединном возвышении.
Пептиды, синтезирующиеся в паравентрикулярных и супрахиазматических ядрах, основными из которых являются вазопрессин и окситоцин, наиболее активно влияют на репродуктивную систему. Основная гормональная функция вазопрессина на уровне аденогипофиза — стимуляция выделения АКТГ за счет усиления действия КРГ. Имеются данные о тормозном влиянии вазопрессина на секрецию ПРЛ за счет повышения обмена ДА в тубероинфундибулярных нейронах. Стимулирующий эффект вазопрессина на выделение ПРЛ может опосредоваться опиатными нейронами. ТРГ, синтезирующийся в паравентрикулярных ядрах, помимо своего прямого действия на секрецию ТТГ, способен стимулировать выделение ПРЛ и гормона роста, и его эффект усиливается под влиянием эстрогенов. ТРГ также оказывает влияние на обмен нейромедиаторов в мозге. Из группы гастроинтестинальных пептидов следует выделить вазоактивный интестинальный полипептид (ВИП), основное нейроэндокринное действие которого сводится к стимуляции выделения ПРЛ, а также гормона роста и ТТГ. Этот эффект является эстрогензависимым.
Особое внимание в регуляции гонадотропной функции гипофиза в последние годы уделяется опиоидным пептидам. Главным действием опиоидов является их способность повышать выделение ПРЛ и гормона роста и блокировать секрецию ЛГ, ФСГ и ТТГ. Они специфически ингибируют овуляторный выброс ЛГ и ФСГ. Опиаты способны регулировать амплитуду и частоту пульсации ЛГ за счет модуляции гипоталамического пейсмекера, контролирующего периодичность активации ГнРГ-продуцирующих нейронов. Влияние опиатов на секрецию ЛГ опосредовано повышением секреции ПРЛ. Опиаты блокируют тоническую секрецию ГнРГ на фоне высокого уровня ПРЛ. Регуляция процессов биосинтеза и секреции ПРЛ, как и ЛГ, опиоидными пептидами осуществляется через специфические рецепторы.
Приведенные выше данные о влиянии различных нейромедиаторов адренергической природы и нейропептидов ставят вопрос о том, каким образом разные вещества с различными механизмами действия координируют работу репродуктивной системы. В ряде случаев отмечен синергизм их действия как на гипоталамическом, так и на гипофизарном уровне, в других — антагонизм их влияния. Можно отметить параллелизм в действии различных медиаторов, вырабатываемых в одних и тех же структурах. В настоящее время наблюдается быстрый прогресс в изучении химии и нейроанатомии нейросекреторных нейронов, участвующих в регуляции репродуктивной функции.
В настоящее время цельное представление о функционировании гипоталамо-гипофизарной системы распалось на отдельные направления исследований с использованием новых специфических методических приемов. Интенсивно используются достижения смежных дисциплин, таких как иммунология. Показана тесная взаимосвязь нейроэндокринной и иммунной систем в различные периоды жизни и при различных физиологических состояниях. Например, с возрастом снижается реакция иммунной системы и параллельно снижается активность гормона роста. При беременности снижается секреция пролактина и окситоцина и повышается секреция половых гормонов — эстрогенов, прогестерона. Доказана взаимосвязь нервной, иммунной и эндокринной систем регуляции секреции инсулина. Разработка фундаментальных аспектов нейроэндокринологии открывает широкие перспективы лечения нарушений репродуктивной системы, заболеваний щитовидной железы или надпочечников гормональными средствами или препаратами, обладающими нейрогенными свойствами. Такими препаратами могут быть производные нейромедиаторов адренергической, холинергической природы, а также нейропептиды.
Начиная с конца 50-х годов прошлого столетия внимание исследователей всего мира все больше стало привлекать изучение воздействия гормональных факторов на функцию головного мозга. В результате многочисленных исследований родилась новая наука — психонейроэндокринология. Ее задачами является изучение вопросов, связанных с влиянием гормонов на высшие функции мозга, — от молекулярного уровня до различных сторон психической деятельности.
Проблема воздействия гормональных факторов на высшие интегративные функции мозга чрезвычайно сложна как по существу, так и в силу отсутствия объективной модели, отражающей зависимость между уровнем гормонов в организме и его влиянием на когнитивные функции.
Показано, что гормоны, взаимодействуя с нейромедиаторами, образуют на уровне ЦНС единую координирующую нейрогуморальную систему, контролирующую интегративную деятельность головного мозга. Нарушение биосинтеза или обмена нейромедиаторов, нарушения их взаимодействия с соответствующими рецепторами могут играть патогенетическую роль в развитии ряда нервно-психических состояний. Сложились устойчивые представления о том, что функция гормонов в ЦНС связана с контролем экспресии гормонзависимых генов и регуляции таких процессов, как развитие и старение мозга, регенерация, адаптация, память.
Различают геномный и негеномный механизм действия гормонов. Полипептидные гормоны действуют на специфические рецепторные зоны мембраны клетки. Связывание лиганда поверхностными рецепторами клетки индуцирует внутриклеточный сигнал (цАМФ и/или кальций) и приводит к быстрому результату (активации остеокластов, секреции кортизола и т.п.). Трансдукция сигнала полипептидных гормонов может вовлекать фосфатидилиновый путь или фосфорилирование рецепторов.
Стероидные и тиреоидные гормоны проявляют свое действие главным образом через специфические внутричерепные связывающие белки, которые взаимодействуют с элементами генов клеток-мишеней, что сопровождается изменением транскрипции специфических генов.
Гипофиз — главный регулятор функционирования эндокринных желез
В аденогипофизе секретируются следующие гормоны: ТТГ, АКТГ, ЛГ и ФСГ, которые оказывают стимулирующее влияние соответственно на клетки щитовидной железы, коры надпочечников и половые железы. Попадая в кровь, гормоны периферических желез активизируют деятельность всех органов и тканей организма, в том числе и аденогипофиза. Осуществляя обратную связь, они блокируют избыточное поступление тропных гормонов гипофиза. Наступает равновесие между мутацией гормонов гипофиза и потребностью организма в гормонах периферических желез. Несколько иной результат наблюдается в отношении ПРА и гормона роста, не имеющих специализированного целевого органа, секреция которого оказывала бы тормозной эффект на выделение этих гормонов. Помимо блокирующего эффекта гормонов периферических желез на синтез и секрецию тропных гормонов гипофиза, аналогичным эффектом обладают также ингибиторные факторы гипоталамического происхождения.
Синдромы эндокринной недостаточности, обусловленные полным или частичным выпадением функций передней и задней долей гипофиза, обозначаются как гипопитуаризм. Гипофункция гипофиза может быть следствием поражения самого гипофиза или гипоталамуса. Результат один — сниженная секреция гипофизарных гормонов с последующим дефицитом гормонов периферических желез.
Уровень гормона роста в крови в норме очень низок, но увеличивается при физической нагрузке, во время сна и при стрессе. Наиболее надежные стимулирующие тесты для данного гормона — индуцированная инсулином гипогликемия, инфузия аргинина или введение L-допа. Эти тесты сопровождаются увеличением секреции гипоталамического соматолиберина.
Секреция пролактина у здоровых людей обычно низкая в течение дня, увеличивается во сне, при стрессе и родах. В клинике используют провоцирующий тест с внутривенным введением ТРГ
Гипоталамус , состоящий из большого количества дискретных ядер , каждое из которых отвечает за специфические функции , таким образом, гипоталамус регулирует различные функции организма посредством сохранения гомеостаза и секрецией гормонов. Эти функции включают в себя контроль за : 1) расходом энергии — посредством регуляции потребления пищи, регуляции процесса пищеварения и уровня метаболизма; 2) уровнем артериального давления и водно — электролитный баланс посредством регуляции потребления воды и соли; 3) репродукцией — через гормональную регуляцию полового созревания , скрещивания ( спаривания ) , беременности и лактации; 4) над температурой тела — посредством регуляции различных процессов терморегуляции ; 5) быстрой реакции — за счет выделения гормонов «оси стресса» и активации симпатического отдела вегетативной неврной системы; 6) циркадианными ритмами и циклом «сон — бодрствование». Гипоталамуса представляет собой «коллектор параллельных сетей» , которые регулируют какие — либо функции организма. Некоторые гипоталамические ядра получают входящую информацию от сенсорных систем , предоставляющих необходимую ( релевантную ) информацию об окружающей среде , так , в частности. свет , воспринимаемый визуальной системой , позволяет контролировать циркадианные ритмы , или аромат феромонов — предупреждает об опасности со стороны хищника или , напротив , о возможности добычи. Многие гипоталамчиеские ядра получают интерорецептивную информацию от внутренних органов , а гипоталмические сети регулируют процессы гомеостаза. , которые координируют различные физиологические процессы , обеспечивающие поддержку постоянной внутренней среды организма, стабильное аретриальное давление , температуру тела или определенный уровень потребления пищи. Биологическая оценка этих путей активации специфическими гипоталамическими ядрами позволяет осуществлять контроль вегетативной нервной системы, поведения , регуляции активности эндокринных желез. Гипоталамус регулирует гомеостаз в сравнительно узком диапазоне анализа входящих сигналов, так , например, если в результате физических упражнений повышается температура тела , гипоталамус дает сигнал симпатическому отделу вегетативной нервной системы о повышении потоотделения и расширении сосудов. ( усиливает отдачу тепла посредством увеличения кровотока в сосудах кожи ).
Нейроны гипоталамуса оказывают прямое влияние за счет проекций своих аксонов на заднюю долю гипофиза (непосредственно поступая в кровеносное русло) , здесь вырабатываются два пептидных гормона : окситоцин и вазопрессин, первый регулирует материнское и социальное поведение , второй — водно — электроллитный баланс и также социальное поведение. Кроме того, окситоцин и вазопрессин могут также выделяться из синаптических терминалей в качестве пептидо — нейротрансмиттеров , которые передают сигналы на постсинаптические нейроны.
Гипоталамические нейроны оказывают стимулирующее действие на эндокринные клетки переднего гипофиза, выделяя так называемые «пре — гормоны». Кортикотропин — релизинг гормон ( CRH) стимулирует выделение адренокортикотропного гормона ( ACTH) , который в свою очередь стимулирует выделение глюкокортикоидов из надпочечников ( кортизол ) , вся цепочка регулирует реакции на стимулы стресса. Гонадотропин — релизиг гормон ( GnRH) стиммулирует выделение лютеинизирующего гормона (LH) и фолликулостимулирующего гормона ( FSH) , эти гормоны стимулируют выделение половых гормонов , половое созревание и сексуальное поведение. Гормон стимулирующий выделение гормона роста ( GHRH) способствует выделению гормона роста ( GH) . Тиротропин — релизиг гормон ( TRH) активирует выделение тироид стимулирующего гормона ( TSH) — тиротропина и пролактина. Первый стимулирует процессы метаболизма, второй — продукцию молока. Гипоталамические нейроны также оказывают тормозное ( подавляющее, ингибирующее) действие на эндокринные клетки переднего гипофиза : соматостатин — на гормон роста , дофамин — на пролактин.
Категория сообщения в блог:
 Гипофиз является очень важной железой внутренней секреции. Он располагается на нижней поверхности головного мозга, в гипофизарной ямке турецкого седла клиновидной кости. Отросток твёрдой мозговой оболочки – диафрагма седла – отделяет гипофиз от полости черепа. Воронка соединяет гипофиз с гипоталамусом.
Гипофиз является очень важной железой внутренней секреции. Он располагается на нижней поверхности головного мозга, в гипофизарной ямке турецкого седла клиновидной кости. Отросток твёрдой мозговой оболочки – диафрагма седла – отделяет гипофиз от полости черепа. Воронка соединяет гипофиз с гипоталамусом.
Снаружи тело гипофиза покрыто соединительнотканной капсулой. Размеры гипофиза достаточно индивидуальны (10-17) х 16 х (5-10) мм, масса у мужчин составляет около 0,5-0,6 г, у женщин – около 0,6-0,7 г. Будучи анатомически единым образованием, гипофиз делится на две доли. Передняя доля (аденогипофиз) крупнее, она занимает 70-80% всей массы гипофиза и включает три части: дистальную, бугорную и промежуточную. В задней доле (нейрогипофиз) различают нервную часть и воронку.
Строение гипофиза
Физиологическая роль гипофиза обусловливает особенности его кровоснабжения. Нижние гипофизарные артерии отходят от внутренних сонных артерий. Верхние гипофизарные артерии отходят от сосудов артериального круга, направляются к серому бугру и воронке, где образуют между собой анастомоз и распадаются на капилляры, проникающие в ткань. На этих капиллярах заканчиваются разветвления аксонов нейросекреторных клеток гипоталамуса, образуя синапсы. Здесь нейросекрет выделяется в кровь. Из петель этой сети формируются воротные венулы, идущие по бугорковой части к передней доле гипофиза, где переходят в широкие синусоидные капилляры. Эти капилляры образуют вторичную гемокапиллярную сеть, оплетающую группы секреторных клеток. Капилляры вторичной сети, сливаясь, образуют выносящие вены, по которым кровь (с гормонами передней доли) выносится из гипофиза. Задняя доля гипофиза снабжается кровью в основном за счёт нижних гипофизарных артерий. Между верхними и нижними артериями существуют анастомозы.
Клетки передней доля гипофиза называются аденоцитами. Существует 2 типа аденоцидов:
- хромофильные аденоциты – крупные клетки, которые хорошо окрашиваются красителями;
- хромофобные аденоциты – мелкие клетки, слабо окрашиваются.
Среди хромофильных аденоцитов различают ацидофильные – клетки овальной формы, окрашивающиеся кислыми красителями в розовый цвет, а также крупные базофильные клетки, окрашивающиеся основными красителями и богатые глюкопротеидными включениями. Задняя часть передней доли, расположенная в виде прослойки между ней и задней долей, рассматривается как промежуточная часть. Она образована многослойным эпителием, среди клеток которого находятся пузырьки (псевдофолликулы).
Задняя доля образована клетками эпендимы (питуицитами), мелкими многоотростчатыми клетками и нервными волокнами, аксонами клеток супраоптического и паравентрикулярного ядер гипоталамуса, разветвления которых оканчиваются на капиллярах задней доли. Воронка гипофиза, соединяясь с воронкой гипоталамуса, образует ножку гипофиза.
Функции гипофиза
Функции гипофиза обусловлены действием выделяемых им гормонов. Посредством выделения этих веществ осуществляются: регуляция синтеза и секреции гормонов гипофизозависимыми эндокринными железами (надпочечники, половые железы); синтез и секреция меланинов; регуляция роста и созревания органов; координация функций различных органов (почки, матка, молочные железы).
В передней доле гипофиза синтезируются так называемые тропные гормоны:
- соматотропин (соматотропный гормон, гормон роста);
- тиреотропин (тиреотропный гормон);
- адренокортикотропный гормон (АКТГ);
- гонадотропные гормоны (фолликулотропин, лютеотропин);
- лактогенный гормон (пролактин);
- меланоцитостимулирующий гормон (меланоцитогропин).
Эти гормоны регулируют синтез и секрецию гормонов гипофизозависимых желез по принципу обратной связи: при снижении концентрации определённого гормона в крови клетки аденогипофиза выделяют сигнальный гормон, который стимулирует образование гормона этой железой, а повышение его уровня в крови приводит к замедлению секреции сигнального гормона.
В промежуточной части вырабатываются липотропные факторы гипофиза, оказывающие влияние на мобилизацию и утилизацию жиров в организме. Нейросекреторные клетки ядер гипоталамуса вырабатывают вазопрессин и окситоцин, которые транспортируются в заднюю долю гипофиза, откуда разносятся кровью.
Соматотропин секретируется не постоянно, а периодически, 3-4 раза вдень. Секреция его увеличивается во время голодания, тяжёлого физического труда и во время сна. С возрастом выработка соматотропина заметно уменьшается, однако сохраняется в течение всей жизни. Гормон роста оказывает двойное воздействие на клетки организма: в клетках усиливается распад накопленных углеводов и жиров, а также их мобилизация для энергетического и пластического обмена, под влиянием выработанных печенью соматомединов усиливается рост костей, синтез белка и деление клеток.
Недостаточная секреция соматотропина приводит к карликовости при сохранении нормального телосложения. Избыточная секреция – к гигантизму. Если гиперсекреция начинается у человека уже в зрелом возрасте, то развивается акромегалия. При этом непропорционально удлиняются конечности, нос и подбородок, язык и пищеварительные органы.
Адренокортикотропный гормон стимулирует выделение кортикостероидов клетками коркового вещества надпочечников. Секреция этого гормона усиливается при некоторых эмоциональных состояниях (страх, гнев, хронический стресс).
Тиреотропный гормон регулирует выделение гормонов щитовидной железы. Он активирует аденилатциклазу и увеличивает поглощение йода клетками железы. Помимо этого тиреотропин оказывает влияние на белковый обмен – увеличивает синтез белков, нуклеиновых кислот, а также увеличивает количество и размер тиреоидных клеток.
Гонадотропные гормоны стимулируют функции половых желез. Фолликулотропин регулирует у женщин развитие фолликулов в яичниках, а у мужчин – образование сперматозоидов и развитие простаты. Лютеотропин стимулирует выработку андрогенов (андростендиол, тестостерон и пр.) и эстрогенов (эстрадиол, эстриол и др.).
Пролактин увеличивает продукцию прогестерона в жёлтом теле яичника и продукцию молока (лактацию). Физиологическая роль этого гормона окончательно не выяснена, но почти все его известные эффекты связаны с размножением.
Меланоцитотронин регулирует распределение пигмента меланина и обусловливает, таким образом, окраску волос и кожи. Пигментные пятна при беременности и усиленная пигментация кожи у пожилых людей возникают в результат гиперфункции промежуточной доли гипофиза.
Вазопрессин участвует в регуляции мочеобразования, усиливая обратное всасывание воды из первичной мочи, обеспечивает водно-солевой гомеостаз организма.
Окситоцин стимулирует гладкую мускулатуру матки во время родов и секрецию молока. Он вызывает сокращение миоэпителиальных клеток, окружающих альвеолы и протоки молочной железы, благодаря чему молоко выделяется из груди.
С нарушениями функции гипофиза обращайтесь к врачу-эндокринологу в Краснодаре.