в условии
в решении
в тексте к заданию
в атрибутах
Категория:
Атрибут:
Всего: 88 1–20 | 21–40 | 41–60 | 61–80 …
Добавить в вариант
Какой из процессов обеспечивает эукариотические клетки энергией наиболее эффективно?
1) фотосинтез
2) гликолиз
3) спиртовое брожение
4) окислительное фосфорилирование
Все приведённые ниже признаки, кроме двух, можно использовать для описания митохондрий. Определите два признака, «выпадающих» из общего списка, и запишите в ответ цифры, под которыми они указаны.
1) не делятся в течение жизни клетки
2) имеют собственный генетический материал
3) являются одномембранными
4) содержат ферменты окислительного фосфорилирования
5) имеют двойную мембрану
Источник: РЕШУ ЕГЭ
Установите соответствие между характеристиками и названиями функций живого вещества в биосфере (по В. И. Вернадскому): к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ХАРАКТЕРИСТИКА
А) выделение метана в атмосферу
в результате деятельности денитрифицирующих бактерий
Б) образование воды и углекислого газа в процессе дыхания аэробов
В) накопление солей кремния в клетках хвощей
Г) восстановление углекислого газа в процессе фотосинтеза
Д) образование известняка
ФУНКЦИЯ
1) окислительно-восстановительная
2) газовая
3) концентрационная
Запишите в таблицу выбранные цифры под соответствующими буквами.
| А | Б | В | Г | Д |
Установите соответствие между процессами и функциями вещества биосферы: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ПРОЦЕССЫ
А) переход двухвалентного железа в трёхвалентное под действием железобактерий
Б) преобразование сероводорода в серу серобактериями
В) выделение кислорода в атмосферу при фотосинтезе
Г) накопление кальция в костях человека
Д) выделение молекулярного азота в атмосферу при денитрификации
Е) накопление кремния в стеблях хвощей
ФУНКЦИИ ВЕЩЕСТВА
1) газовая
2) концентрационная
3) окислительно-восстановительная
Запишите в таблицу выбранные цифры под соответствующими буквами.
| А | Б | В | Г | Д | Е |
К процессу окислительного фосфорилирования относится процесс
1) расщепления глюкозы ферментами
3) синтеза глюкозы из неорганических соединений
4) синтеза белков из аминокислот
Источник: Яндекс: Тренировочная работа ЕГЭ по биологии. Вариант 2.
Окислительное фосфорилирование – это этап
1) фотосинтеза
2) гликолиза
3) пластического обмена
4) энергетического обмена
Выберите процессы, относящиеся к энергетическому обмену веществ.
1) выделение кислорода в атмосферу
2) образование углекислого газа, воды, мочевины
3) окислительное фосфорилирование
4) синтез глюкозы
5) гликолиз
6) фотолиз воды
Найдите ошибки в приведённом тексте. Укажите номера предложений, в которых они допущены, исправьте их.
1. При дыхании синтезируется глюкоза через ряд последовательных этапов. 2.На некоторых этапах энергия химических связей глюкозы используется для синтеза АТФ. 3. Дыхание начинается с соединения двух молекул пировиноградной кислоты. 4. Первичный процесс бескислородного дыхания происходит в цитоплазме. 5. В результате этого дыхания образуются две молекулы АТФ. 6. Конечным этапом цикла является окислительное фосфорилирование, на которое расходуется энергия АТФ.
Все приведённые ниже признаки, кроме двух, можно использовать для описания общих свойств характерных для митохондрий и пластид. Определите два признака, «выпадающих» из общего списка, и запишите в ответ цифры, под которыми они указаны.
1) не делятся в течение жизни клетки
2) имеют собственный генетический материал
3) содержат ферменты окислительного фосфорилирования
4) имеют двойную мембрану
5) участвуют в синтезе АТФ
Источник: РЕШУ ЕГЭ
Установите соответствие между характеристиками и органоидами клетки: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ХАРАКТЕРИСТИКИ
А) замкнутая молекула ДНК
Б) окислительные ферменты на кристах
B) внутреннее содержимое — кариоплазма
Г) линейные хромосомы
Д) наличие хроматина в интерфазе
Е) складчатая внутренняя мембрана
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| A | Б | В | Г | Д | Е |
Источник: ЕГЭ по биологии 2017. Досрочная волна
Установите соответствие между особенностями строения органоидов клетки и органоидами: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ОСОБЕННОСТИ СТРОЕНИЯ ОРГАНОИДОВ
А) основу составляет липидный бислой
Б) имеет двумембранную пористую оболочку
В) содержит кариоплазму
Г) в органоиде множество ферментов окислительного цикла
Д) содержит кольцевую хромосому
Е) осуществляет фаго- и пиноцитоз у животных
ОРГАНОИДЫ
1) клеточная мембрана
2) ядро
3) митохондрия
Запишите в таблицу выбранные цифры под соответствующими буквами
| A | Б | В | Г | Д | Е |
Установите соответствие между характеристикой процессов, происходящих при энергетическом обмене, и этапами этого процесса: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ХАРАКТЕРИСТИКА
А) идёт с образованием ПВК
Б) происходит в митохондриях
В) процесс анаэробный
Г) в ходе процесса образуется 36 молей АТФ
Д) образуются углекислый газ, вода, мочевина
Е) происходит в цитоплазме
ПРОЦЕСС
1) гликолиз
2) окислительное
фосфорилирование
Запишите в таблицу выбранные цифры под соответствующими буквами
| A | Б | В | Г | Д | Е |
Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны.
Окислительно-восстановительная функция живого вещества в биосфере проявляется в
1) выделении кислорода в процессе фотосинтеза
2) образовании углекислого газа и воды из глюкозы в процессе дыхания
3) образовании отложений фосфора на дне водоёма
4) восстановлении углекислого газа до глюкозы в процессе фотосинтеза
5) превращении сероводорода в кристаллическую серу бактериями
6) выделении азота денитрифицирующими бактериями
Установите соответствие между процессами и стадиями клеточного дыхания: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ПРОЦЕССЫ
А) окислительное фосфорилирование
Б) транспорт электронов по цепи переносчиков
В) образование пировиноградной кислоты
Г) расщепление шестиуглеродного сахара
Д) активация глюкозы с затратой АТФ
Е) цикл трикарбоновых кислот
СТАДИИ КЛЕТОЧНОГО ДЫХАНИЯ
1) бескислородный этап
2) кислородный этап
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| А | Б | В | Г | Д | Е |
Все перечисленные ниже понятия и процессы, кроме трёх, используют для описания световой стадии фотосинтеза в клетке растения. Определите три понятия, «выпадающих» из общего списка, и запишите в таблицу цифры, под которыми они указаны
1) перемещение электронов
2) фотолиз воды
3) окисление НАДФ·Н
4) восстановление углерода водородом
5) фотофосфорилирование
6) окислительное фосфорилирование
Окислительно-восстановительная функция живого вещества планеты связана с
2) климатическими условиями
3) обменом веществ и энергии
4) освоением организмами новых мест обитания
Какая функция живого вещества проявляется при поглощении бактериями молекулярного азота из воздуха
3) окислительно-восстановительная
4) биохимическая
В чем заключается биологический смысл окислительного фосфорилирования?
Установите соответствие между характеристикой обмена и его видом.
ХАРАКТЕРИСТИКА
А) окисление органических веществ
Б) образование полимеров из мономеров
В) расщепление АТФ
Г) запасание энергии в клетке
Д) репликация ДНК
Е) окислительное фосфорилирование
ВИД ОБМЕНА
1) пластический
2) энергетический
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| A | Б | В | Г | Д | Е |
Источник: Демонстрационная версия ЕГЭ—2013 по биологии
Какой из процессов относится к диссимиляции?
1) окислительное фосфорилирование
4) синтез липидов
Всего: 88 1–20 | 21–40 | 41–60 | 61–80 …
Обмен веществ
Обмен веществ (метаболизм) складывается из процессов расщепления и синтеза — диссимиляции и ассимиляции, постоянно
протекающих в организме. Чтобы жизнь продолжалась, количество поступающей энергии должно превышать (или как минимум равняться)
количеству расходуемой энергии, поэтому диссимиляция и ассимиляция поддерживают определенный баланс друг с другом.

Энергетический обмен
Энергетический обмен (диссимиляция — от лат. dissimilis ‒ несходный) — обратная ассимиляции сторона обмена веществ, совокупность реакций, которые приводят к высвобождению энергии химических связей. Это реакции расщепления жиров,
белков, углеводов, нуклеиновых кислот до простых веществ.
Возможно три этапа диссимиляции: подготовительный, анаэробный и аэробный. Среда обитания определяет количество
этапов диссимиляции. Их может быть три, если организм обитает в кислородной среде, и два, если речь идет об
организме, обитающем в бескислородной среде (к примеру, в кишечнике).
Обсудим этапы энергетического обмена более подробно:
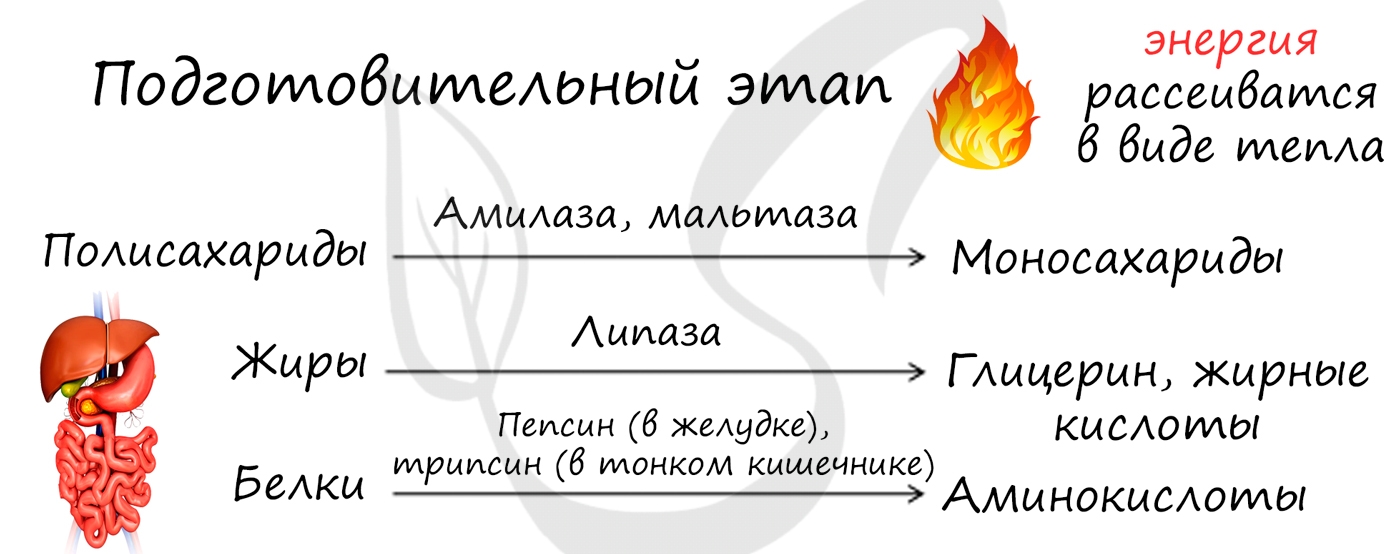
- Подготовительный этап
- Бескислородный этап (анаэробный) — гликолиз
- Кислородный этап (аэробный)
Подготовительный этап осуществляется ферментами в ЖКТ. В результате действия ферментов сложные вещества превращаются в более простые: полимеры распадаются на мономеры. Это сопровождается разрывом химических связей и выделением энергии, большая часть
которой рассеивается в виде тепла.
Под действием ферментов белки расщепляются на аминокислоты, жиры — на глицерин и жирные кислоты, сложные углеводы — до простых сахаров.
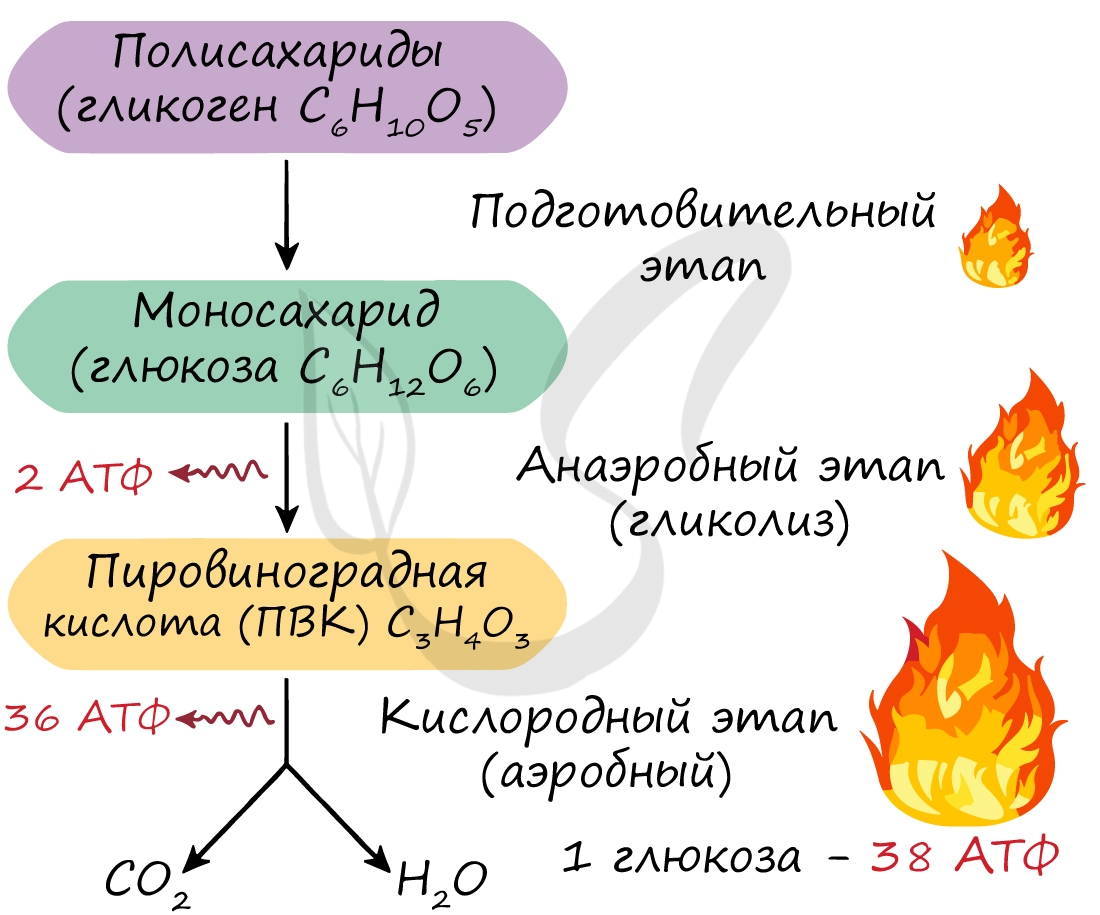
Этот этап является последним для организмов-анаэробов, обитающих в условиях, где кислород отсутствует. На этапе гликолиза
происходит расщепление молекулы глюкозы: образуется 2 молекулы АТФ и 2 молекулы пировиноградной кислоты (ПВК).
Происходит данный этап в цитоплазме клеток.
Этот этап доступен только для аэробов — организмов, живущих в кислородной среде. Из каждой молекулы ПВК, образовавшейся на
этапе гликолиза, синтезируется 18 молекул АТФ — в сумме с двух ПВК выход составляет 36 молекул АТФ.
Таким образом, суммарно с одной молекулы глюкозы можно получить 38 АТФ (гликолиз + кислородный этап).
Кислородный этап протекает на кристах митохондрий (складках, выпячиваниях внутренней мембраны), где наибольшая концентрация окислительных ферментов. Главную роль в этом процессе играет так называемый цикл Кребса, который подробно изучает биохимия.
АТФ — аденозинтрифосфорная кислота
Трудно переоценить роль в клетке АТФ — универсального источника энергии. Молекула АТФ состоит из азотистого основания —
аденина, углевода — рибозы и трех остатков фосфорной кислоты.
Между остатками фосфорной кислоты находятся макроэргические связи — ковалентные связи, которые гидролизуются с выделением
большого количества энергии. Их принято обозначать типографическим знаком тильда «∽».
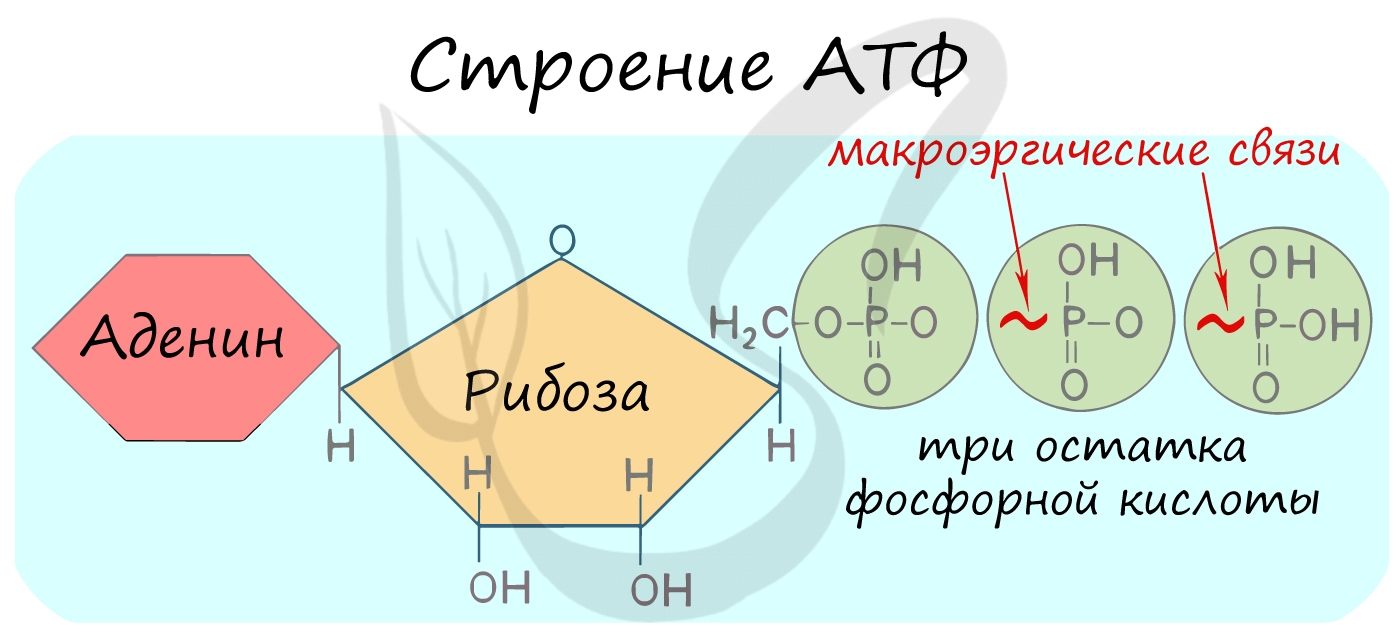
АТФ гидролизуется до АДФ (аденозиндифосфорная кислота), а затем и до АМФ (аденозинмонофосфорная кислота).
Гидролиз АТФ сопровождается выделением энергии (E) на каждом этапе и может быть представлен такой схемой:
- АТФ + H2O = АДФ + H3PO4 + E
- АДФ + H2O = АМФ + H3PO4 + E
- АМФ + H2O = аденин + рибоза + H3PO4 + E
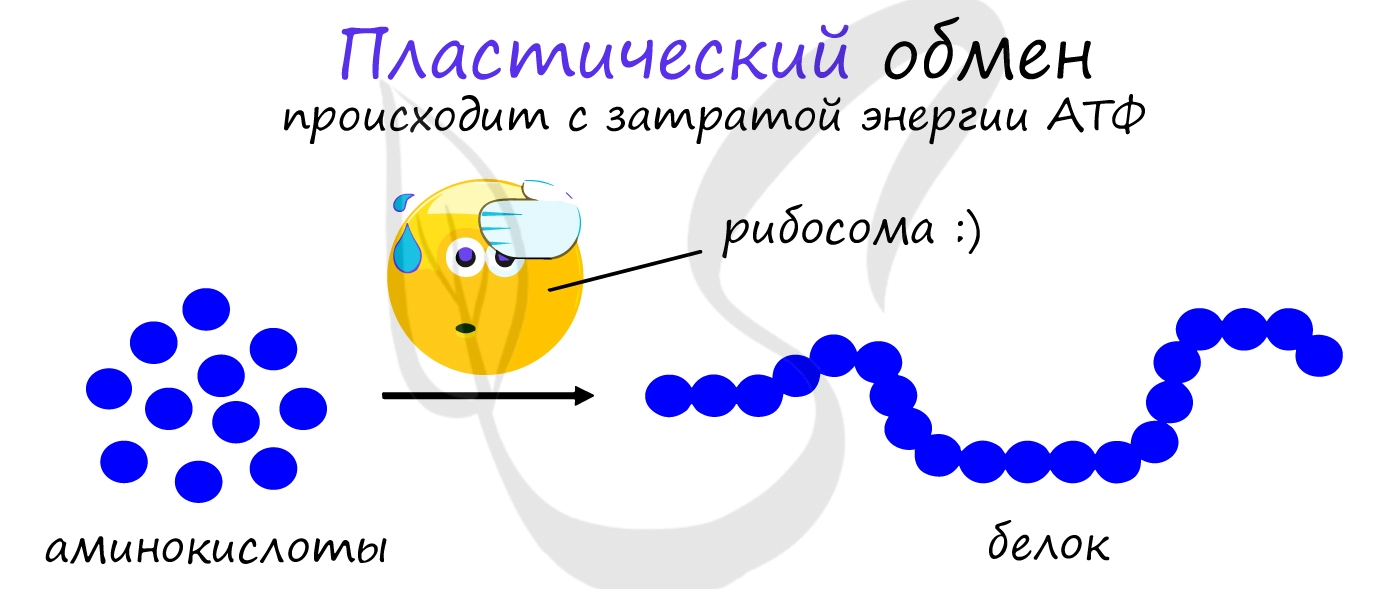
Пластический обмен
АТФ является универсальным источником энергии в клетке: энергия макроэргических связей АТФ используется для реакций
пластического обмена (ассимиляции), протекающих с затратой энергии: синтеза белка на рибосоме (трансляции),
удвоению ДНК (репликации) и т.д.
В результате пластического обмена в нашем организме происходит синтез белков, жиров и углеводов.
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Понятие метаболизма
Метаболизм — совокупность всех химических реакций, протекающих в живом организме. Значение метаболизма состоит в создании необходимых организму веществ и обеспечении его энергией.
Выделяют две составные части метаболизма — катаболизм и анаболизм.
Составные части метаболизма
| Часть | Характеристика | Примеры | Затраты энергии |
| Катаболизм (энергетический обмен, диссимиляция) | Совокупность химических реакций, приводящих к образованию простых веществ из более сложных | Гидролиз полимеров до мономеров и расщепление последних до низкомолекулярных соединений углекислого газа, воды, аммиака и других веществ | Энергия выделяется |
| Анаболизм (пластический обмен, ассимиляция) | Совокупность химических реакций синтеза сложных веществ из более простых | Образование углеводов из углекислого газа и воды в процессе фотосинтеза, реакции матричного синтеза | Энергия поглощается |
Процессы пластического и энергетического обмена неразрывно связаны между собой. Все синтетические (анаболические) процессы нуждаются в энергии, поставляемой в ходе реакций диссимиляции. Сами же реакции расщепления (катаболизма) протекают лишь при участии ферментов, синтезируемых в процессе ассимиляции.
Роль ФТФ в метаболизме
Энергия, высвобождающаяся при распаде органических веществ, не сразу используется клеткой, а запасается в форме высокоэнергетических соединений, как правило, в форме аденозинтрифосфата (АТФ). По своей химической природе АТФ относится к мононуклеотидам.
АТФ (аденозинтрифосфорная кислота) — мононуклеотид, состоящий из аденина, рибозы и трёх остатков фосфорной кислоты, соединяющихся между собой макроэргическими связями.
В этих связях запасена энергия, которая высвобождается при их разрыве:
АТФ + H2O → АДФ + H3PO4 + Q1
АДФ + H2O → АМФ + H3PO4 + Q2
АМФ + H2O → аденин + рибоза + H3PO4 + Q3,
где АТФ — аденозинтрифосфорная кислота; АДФ — аденозиндифосфорная кислота; АМФ — аденозинмонофосфорная кислота; Q1 = Q2 = 30,6 кДж; Q3 = 13,8 кДж.
Запас АТФ в клетке ограничен и пополняется благодаря процессу фосфорилирования. Фосфорилирование — присоединение остатка фосфорной кислоты к АДФ (АДФ + Ф → АТФ). Он происходит с разной интенсивностью при дыхании, брожении и фотосинтезе. АТФ обновляется чрезвычайно быстро (у человека продолжительность жизни одной молекулы АТФ менее 1 мин).
Энергия, накопленная в молекулах АТФ, используется организмом в анаболических реакциях (реакциях биосинтеза). Молекула АТФ является универсальным хранителем и переносчиком энергии для всех живых существ.
Энергетический обмен
Энергию, необходимую для жизнедеятельности, большинство организмов получают в результате процессов окисления органических веществ, то есть в результате катаболических реакций. Важнейшим соединением, выступающим в роли топлива, является глюкоза.
По отношению к свободному кислороду организмы делятся на три группы.
Классификация организмов по отношению к свободному кислороду
| Группа | Характеристика | Организмы |
| Аэробы (облигатные аэробы) | Организмы, способные жить только в кислородной среде | Животные, растения, некоторые бактерии и грибы |
| Анаэробы (облигатные анаэробы) | Организмы, неспособные жить в кислородной среде | Некоторые бактерии |
| Факультативные формы (факультативные анаэробы) | Организмы, способные жить как в присутствии кислорода, так и без него | Некоторые бактерии и грибы |
У облигатных аэробов и факультативных анаэробов в присутствии кислорода катаболизм протекает в три этапа: подготовительный, бес- кислородный и кислородный. В результате органические вещества распадаются до неорганических соединений. У облигатных анаэробов и факультативных анаэробов при недостатке кислорода катаболизм протекает в два первых этапа: подготовительный и бескислородный. В результате образуются промежуточные органические соединения, еще богатые энергией.
Этапы катаболизма
1. Первый этап — подготовительный — заключается в ферментативном расщеплении сложных органических соединений на более простые. Белки расщепляются до аминокислот, жиры — до глицерина и жирных кислот, полисахариды — до моносахаридов, нуклеиновые кислоты — до нуклеотидов. У многоклеточных организмов это происходит в желудочно-кишечном тракте, у одноклеточных — в лизосомах под действием гидролитических ферментов. Высвобождающаяся при этом энергия рассеивается в виде теплоты. Образовавшиеся органические соединения либо подвергаются дальнейшему окислению, либо используются клеткой для синтеза собственных органических соединений.
2. Второй этап — неполное окисление (бескислородный) — заключается в дальнейшем расщеплении органических веществ, осуществляется в цитоплазме клетки без участия кислорода. Главным источником энергии в клетке является глюкоза. Бескислородное, неполное окисление глюкозы называется гликолизом. В результате гликолиза одной молекулы глюкозы образуется по две молекулы пировиноградной кислоты (ПВК, пируват) CH3COCOOH, АТФ и воды, а также атомы водорода, которые связываются молекулой-переносчиком НАД+ и запасаются в виде НАД·Н.
Суммарная формула гликолиза имеет следующий вид:
C6H12O6 + 2H3PO4 + 2АДФ + 2НАД+ → 2C3Н4O3 + 2H2O + 2АТФ + 2НАД·Н.
Далее при отсутствии в среде кислорода продукты гликолиза (ПВК и НАД·Н) перерабатываются либо в этиловый спирт — спиртовое брожение (в клетках дрожжей и растений при недостатке кислорода)
CH3COCOOH → СО2 + СН3СОН
СН3СОН + 2НАД·Н → С2Н5ОН + 2НАД+,
либо в молочную кислоту — молочнокислое брожение (в клетках животных при недостатке кислорода)
CH3COCOOH + 2НАД·Н → C3Н6O3 + 2НАД+.
При наличии в среде кислорода продукты гликолиза претерпевают дальнейшее расщепление до конечных продуктов.
3. Третий этап — полное окисление (дыхание) — заключается в окислении ПВК до углекислого газа и воды, осуществляется в митохондриях при обязательном участии кислорода.
Он состоит из трёх стадий:
А) образование ацетилкоэнзима А;
Б) окисление ацетилкоэнзима А в цикле Кребса;
В) окислительное фосфорилирование в электронотранспортной цепи.
А. На первой стадии ПВК переносится из цитоплазмы в митохондрии, где взаимодействует с ферментами матрикса и образует 1) диоксид углерода, который выводится из клетки; 2) атомы водорода, которые молекулами-переносчиками доставляются к внутренней мембране митохондрии; 3) ацетилкофермент А (ацетил-КоА).
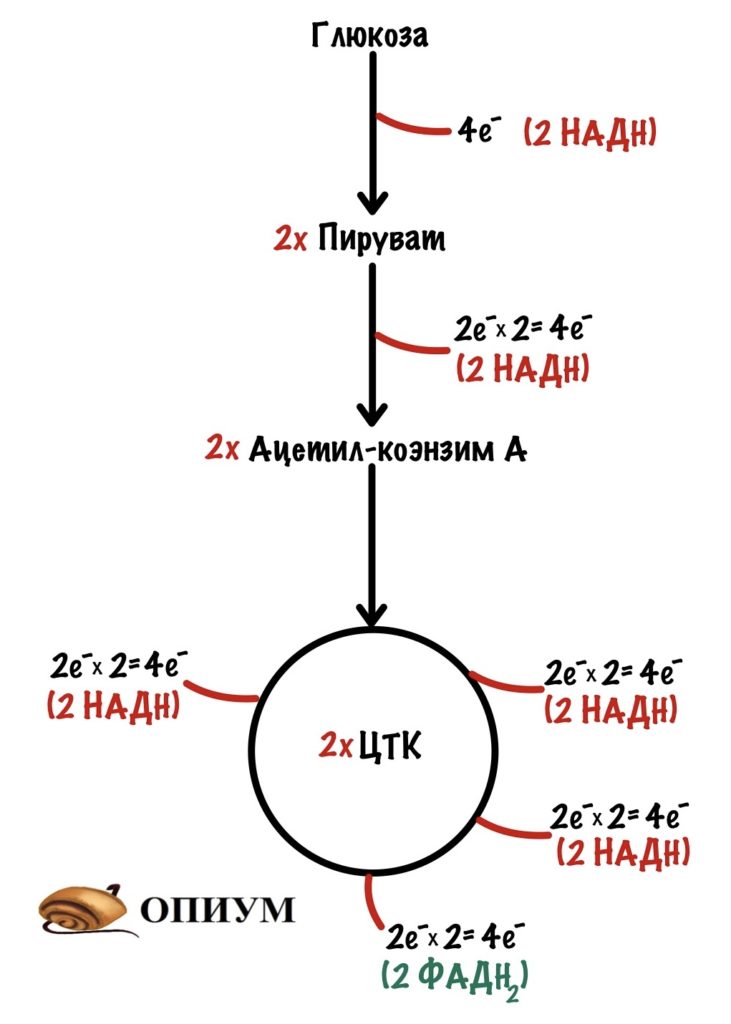
Б. На второй стадии происходит окисление ацетилкоэнзима А в цикле Кребса. Цикл Кребса (цикл трикарбоновых кислот, цикл лимонной кислоты) — это цепь последовательных реакций, в ходе которых из одной молекулы ацетил-КоА образуются 1) две молекулы диоксида углерода, 2) молекула АТФ и 3) четыре пары атомов водорода, передаваемые на молекулы-переносчики — НАД и ФАД. Таким образом, в результате гликолиза и цикла Кребса молекула глюкозы расщепляется до СО2, а высвободившаяся при этом энергия расходуется на синтез 4 АТФ и накапливается в 10 НАД·Н и 4 ФАД·Н2.
В. На третьей стадии атомы водорода с НАД·Н и ФАД·Н2 окисляются молекулярным кислородом О2 с образованием воды. Один НАД·Н способен образовывать 3 АТФ, а один ФАД·Н2–2 АТФ. Таким образом, выделяющаяся при этом энергия запасается в виде ещё 34 АТФ.
Этот процесс протекает следующим образом. Атомы водорода концентрируются около наружной стороны внутренней мембраны митохондрии. Они теряют электроны, которые по цепи молекул-переносчиков (цитохромов) электронотранспортной цепи (ЭТЦ) переносятся на внутреннюю сторону внутренней мембраны, где соединяются с молекулами кислорода:
О2 + е— → О2—.
В результате деятельности ферментов цепи переноса электронов внутренняя мембрана митохондрий изнутри заряжается отрицательно (за счёт О2—), а снаружи — положительно (за счёт Н+), так что между её поверхностями создаётся разность потенциалов. Во внутреннюю мембрану митохондрий встроены молекулы фермента АТФ- синтетазы, обладающие ионным каналом. Когда разность потенциалов на мембране достигает критического уровня, положительно заряженные частицы H+ силой электрического поля начинают проталкиваться через канал АТФазы и, оказавшись на внутренней поверхности мембраны, взаимодействуют с кислородом, образуя воду:
1/2О2— +2H+ → Н2О.
Энергия ионов водорода H+, транспортирующихся через ионный канал внутренней мембраны митохондрии, используется для фосфорилирования АДФ в АТФ:
АДФ + Ф → АТФ.
Такое образование АТФ в митохондриях при участии кислорода называется окислительным фосфорилированием.
Суммарное уравнение расщепления глюкозы в процессе клеточного дыхания:
C6H12O6 + 6O2 + 38H3PO4 + 38АДФ → 6CO2 + 44H2O + 38АТФ.
Таким образом, в ходе гликолиза образуются 2 молекулы АТФ, в ходе клеточного дыхания — ещё 36 молекул АТФ, в целом при пол- ном окислении глюкозы — 38 молекул АТФ.
Пластический обмен
Пластический обмен, или ассимиляция, представляет собой совокупность реакций, обеспечивающих синтез сложных органических соединений из более простых (фотосинтез, хемосинтез, биосинтез белка и др.).
Гетеротрофные организмы строят собственные органические вещества из органических компонентов пищи. Гетеротрофная ассимиляция сводится, по существу, к перестройке молекул:
органические вещества пищи (белки, жиры, углеводы) → простые органические молекулы (аминокислоты, жирные кислоты, моносахариды) → макромолекулы тела (белки, жиры, углеводы).
Автотрофные организмы способны полностью самостоятельно синтезировать органические вещества из неорганических молекул, потребляемых из внешней среды. В процессе фото- и хемосинтеза происходит образование простых органических соединений, из которых в дальнейшем синтезируются макромолекулы:
неорганические вещества (СО2, Н2О) → простые органические молекулы (аминокислоты, жирные кислоты, моносахариды) → макромолекулы тела (белки, жиры, углеводы).
Фотосинтез
Фотосинтез — синтез органических соединений из неорганических за счёт энергии света. Суммарное уравнение фотосинтеза:
Фотосинтез протекает при участии фотосинтезирующих пигментов, обладающих уникальным свойством преобразования энергии солнечного света в энергию химической связи в виде АТФ. Фотосинтезирующие пигменты представляют собой белковоподобные вещества. Наиболее важным является пигмент хлорофилл. У эукариот фотосинтезирующие пигменты встроены во внутреннюю мембрану пластид, у прокариот — во впячивания цитоплазматической мембраны.
Строение хлоропласта очень похоже на строение митохондрии. Во внутренней мембране тилакоидов гран содержатся фотосинтетические пигменты, а также белки цепи переноса электронов и молекулы фермента АТФ-синтетазы.
Процесс фотосинтеза состоит из двух фаз: световой и темновой.
1. Световая фаза фотосинтеза протекает только на свету в мембране тилакоидов граны.
К ней относятся поглощение хлорофиллом квантов света, образование молекулы АТФ и фотолиз воды.
Под действием кванта света (hv) хлорофилл теряет электроны, переходя в возбуждённое состояние:
Эти электроны передаются переносчиками на наружную, то есть обращенную к матриксу поверхность мембраны тилакоидов, где накапливаются.
Одновременно внутри тилакоидов происходит фотолиз воды, то есть её разложение под действием света:
Образующиеся электроны передаются переносчиками к молекулам хлорофилла и восстанавливают их. Молекулы хлорофилла возвращаются в стабильное состояние.
Протоны водорода, образовавшиеся при фотолизе воды, накапливаются внутри тилакоида, создавая Н+-резервуар. В результате внутренняя поверхность мембраны тилакоида заряжается положительно (за счёт Н+), а наружная — отрицательно (за счёт е—). По мере накопления по обе стороны мембраны противоположно заряженных частиц нарастает разность потенциалов. При достижении критической величины разности потенциалов сила электрического поля начинает проталкивать протоны через канал АТФ-синтетазы. Выделяющаяся при этом энергия используется для фосфорилирования молекул АДФ:
АДФ + Ф → АТФ.
Образование АТФ в процессе фотосинтеза под действием энергии света называется фотофосфорилированием.
Ионы водорода, оказавшись на наружной поверхности мембраны тилакоида, встречаются там с электронами и образуют атомарный водород, который связывается с молекулой-переносчиком водорода НАДФ (никотинамидадениндинуклеотидфосфат):
2Н+ + 4е– + НАДФ+ → НАДФ·Н2.
Таким образом, во время световой фазы фотосинтеза происходят три процесса: образование кислорода вследствие разложения воды, синтез АТФ и образование атомов водорода в форме НАДФ·Н2. Кислород диффундирует в атмосферу, а АТФ и НАДФ·Н2 участвуют в процессах темновой фазы.
2. Темновая фаза фотосинтеза протекает в матриксе хлоропласта как на свету, так и в темноте и представляет собой ряд последовательных преобразований СО2, поступающего из воздуха, в цикле Кальвина. Осуществляются реакции темновой фазы за счёт энергии АТФ. В цикле Кальвина СО2 связывается с водородом из НАДФ·Н2 с образованием глюкозы.
В процессе фотосинтеза кроме моносахаридов (глюкоза и др.) синтезируются мономеры других органических соединений — аминокислоты, глицерин и жирные кислоты. Таким образом, благодаря фотосинтезу растения обеспечивают себя и всё живое на Земле необходимыми органическими веществами и кислородом.
Сравнительная характеристика фотосинтеза и дыхания эукариот представлена в таблице.
Сравнительная характеристика фотосинтеза и дыхания эукариот
| Признак | Фотосинтез | Дыхание |
| Уравнение реакции | 6СО2 + 6Н2О + энергия света → C6H12O6 + 6O2 | C6H12O6 + 6O2 → 6СО2 + 6Н2О + энергия (АТФ) |
| Исходные вещества | Углекислый газ, вода | Органические вещества, кислород |
| Продукты реакции | Органические вещества, кислород | Углекислый газ, вода |
| Значение в круговороте веществ | Синтез органических веществ из неорганических | Разложение органических веществ до неорганических |
| Превращение энергии | Превращение энергии света в энергию химических связей органических веществ | Превращение энергии химических связей органических веществ в энергию макроэргических связей АТФ |
| Важнейшие этапы | Световая и темновая фаза (включая цикл Кальвина) | Неполное окисление (гликолиз) и полное окисление (включая цикл Кребса) |
| Место протекания процесса | Хлоропласты | Гиалоплазма (неполное окисление) и митохондрии (полное окисление) |
Генетическая информация у всех организмов хранится в виде определённой последовательности нуклеотидов ДНК (или РНК у РНК-содержащих вирусов). Прокариоты содержат генетическую информацию в виде одной молекулы ДНК. В эукариотических клетках генетический материал распределён в нескольких молекулах ДНК, организованных в хромосомы.
ДНК состоит из кодирующих и некодирующих участков. Кодирующие участки кодируют РНК. Некодирующие области ДНК выполняют структурную функцию, позволяя участкам генетического материала упаковываться определённым образом, или регуляторную функцию, участвуя во включении генов, направляющих синтез белка.
Кодирующими участками ДНК являются гены. Ген — участок молекулы ДНК, кодирующей синтез одной мРНК (и соответственно полипептида), рРНК или тРНК.
Участок хромосомы, где расположен ген называется локусом. Совокупность генов клеточного ядра представляет собой генотип, совокупность генов гаплоидного набора хромосом — гено́м, совокупность генов внеядерных ДНК (митохондрий, пластид, цитоплазмы) — плазмон.
Реализация информации, записанной в генах, через синтез белков называется экспрессией (проявлением) генов. Генетическая информация хранится в виде определённой последовательности нуклеотидов ДНК, а реализуется в виде последовательности аминокислот в белке. Посредниками, переносчиками информации выступают РНК. То есть реализация генетической информации происходит следующим образом:
ДНК → РНК → белок.
Этот процесс осуществляется в два этапа:
1) транскрипция;
2) трансляция.
Транскрипция (от лат. transcriptio — переписывание) — синтез РНК с использованием ДНК в качестве матрицы. В результате образуются мРНК, тРНК и рРНК. Процесс транскрипции требует больших затрат энергии в виде АТФ и осуществляется ферментом РНК-полимеразой.
Одновременно транскрибируется не вся молекула ДНК, а лишь отдельные её отрезки. Такой отрезок (транскриптон) начинается промотором — участком ДНК, куда присоединяется РНК-полимераза и откуда начинается транскрипция, а заканчивается терминатором — участком ДНК, содержащим сигнал окончания транскрипции. Транскриптон — это ген с точки зрения молекулярной биологии.
Транскрипция, как и репликация, основана на способности азотистых оснований нуклеотидов к комплементарному связыванию. На время транскрипции двойная цепь ДНК разрывается, и синтез РНК осуществляется по одной цепи ДНК.
В процессе транскрипции последовательность нуклеотидов ДНК переписывается на синтезирующуюся молекулу мРНК, которая выступает в качестве матрицы в процессе биосинтеза белка.
Гены прокариот состоят только из кодирующих нуклеотидных последовательностей.
Гены эукариот состоят из чередующихся кодирующих (экзонов) и некодирующих (интронов) участков.
После транскрипции участки мРНК, соответствующие интронам, удаляются в ходе сплайсинга, являющегося составной частью процессинга.
Процессинг — процесс формирования зрелой мРНК из её предшественника пре-мРНК. Он включает два основных события. 1.Присоединение к концам мРНК коротких последовательностей нуклеотидов, обозначающих место начала и место конца трансляции. Сплайсинг — удаление неинформативных последовательностей мРНК, соответствующих интронам ДНК. В результате сплайсинга молекулярная масса мРНК уменьшается в 10 раз.
Трансляция (от лат. translatio — перевод) — синтез полипептидной цепи с использованием мРНК в роли матрицы.
В трансляции участвуют все три типа РНК: мРНК является информационной матрицей; тРНК доставляют аминокислоты и узнают кодоны; рРНК вместе с белками образуют рибосомы, которые удерживают мРНК, тРНК и белок и осуществляют синтез полипептидной цепи.
Этапы трансляции
| Этап | Характеристика |
| Инициация | Сборка комплекса, участвующего в синтезе полипептидной цепи. Малая субчастица рибосомы соединяется с инициаторной мет-трнк, а затем с мрнк, после чего происходит образование целой рибосомы, состоящей из малой и большой субчастиц. |
| Элонгация | Удлинение полипептидной цепи. Рибосома перемещается вдоль мрнк, что сопровождается многократным повторением цикла присоединения очередной аминокислоты к растущей полипептидной цепи. |
| Терминация | Завершение синтеза полипептидной молекулы. Рибосома достигает одного из трёх стоп-кодонов мрнк, а так как не существует трнк с антикодонами, комплементарными стоп-кодонам, синтез полипептидной цепи прекращается. Она высвобождается и отделяется от рибосомы. Рибосомные субчастицы диссоциируют, отделяются от мрнк и могут принять участие в синтезе следующей полипептидной цепи. |
Реакции матричного синтеза. К реакциям матричного синтеза относятся
- самоудвоение ДНК (репликация);
- образование мРНК, тРНК и рРНК на молекуле ДНК (транскрипция);
- биосинтез белка на мРНК (трансляция).
Все эти реакции объединяет то, что молекула ДНК в одном случае или молекула мРНК в другом выступают в роли матрицы, на которой происходит образование одинаковых молекул. Реакции матричного синтеза являются основой способности живых организмов к воспроизведению себе подобных.
Регуляция экспрессии генов. Тело многоклеточного организма построено из разнообразных клеточных типов. Они отличаются структурой и функциями, то есть дифференцированы. Различия проявляются в том, что помимо белков, необходимых любой клетке организма, клетки каждого типа синтезируют ещё и специализированные белки: в эпидермисе образуется кератин, в эритроцитах — гемоглобин и т. д. Клеточная дифференцировка обусловлена изменением набора экспрессируемых генов и не сопровождается какими-либо необратимыми изменениями в структуре самих последовательностей ДНК.
1. Матрикс митохондрии
содержит все ферменты цикла Кребса. В митохондриях ПВК превращается в ацетил-кофермент А (за счет присоединения КоА). В КоА входит адениловый нуклеотид, а также пантотеновая кислота (синтезируется в кишечнике человека бактериями). Надо помнить, что окисляться могут и жирные кислоты, и аминокислоты — но в результате всегда создается именно ацетил-КоА. При этом идет восстановление HAДН из НАД+ и выделяется СО2.
2. Ацетил-КоА объединяется с молекулой щавелеускусной кислоты, происходит образование лимонной кислоты.
3. Дальше лимонная кислота продолжает окисляться в процессе ферментных реакций.
1) Восстанавливаются 3 молекулы НАДН, одна молекула ФАДН2, и ГТФ (гуанозинтрифосфат).
2) ГТФ используется для фосфорилирования АДФ и образования АТФ.
3) Лимонная кислота утрачивает 2 углеродных атома, в результате чего возникают две молекулы СО2.
4. Пройдя одну за другой семь реакций, лимонная кислота вновь превращается в щавелеуксусную, а та, в свою очередь, вновь соединяется с ацетил-КоА. Цикл замыкается.
1) В лимонной кислоте словно бы сгорает присоединившийся ранее остаток ацетил-КоА.
2) Протоны водорода и электроны переносятся на акцепторы — НАД+ и ФАД.
3) В итоге энергия органических соединений аккумулируется в молекулах НАДН, ФАДН2, АТФ.
4) К тому же, подчеркнем, в цикле Кребса идет выделение CO2.
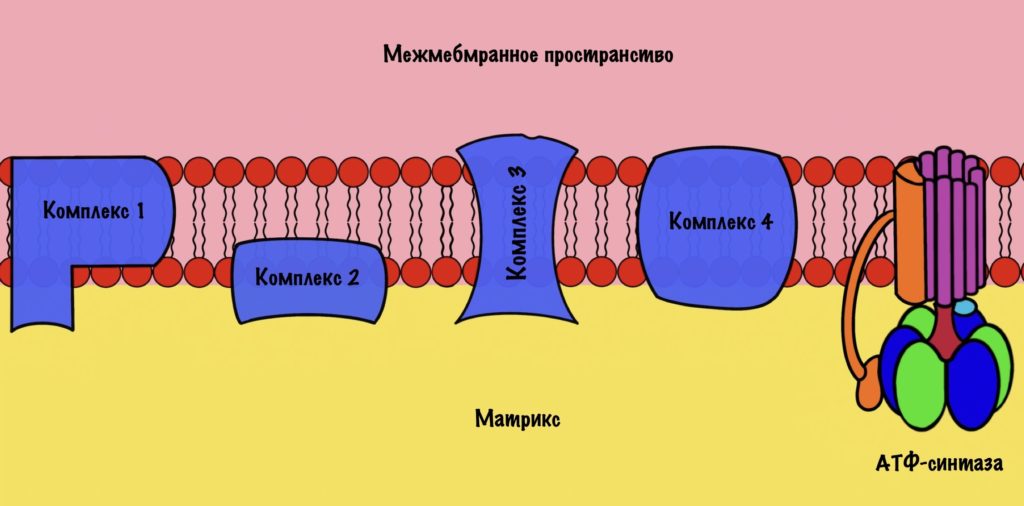
Цепь переноса электронов. Окислительное фосфорилирование
1. Электроны от НАДН и ФАДН2 переносятся по цепи переноса электронов (которая имеет много звеньев) к конечному акцептору — кислороду. В ходе этого процесса освобождается энергия электронов НАДН и ФАДН2, которая идет на синтез АТФ из АДФ. Именно потому данный процесс и назван фосфорилированием. При этом электроны концентрируются с внутренней стороны крист, а протоны с внешней (противоположно тому, как это было в хлоропластах).
2. Перенос электронов идет по цепи, расположенной с внутренней стороны внутренней мембраны митохондрий, где находятся ряд переносчиков. Самый сильный акцептор электронов — в конце цепи — кислород. Промежуточные переносчики: убихинон, ФМН, НАДН-дегидрогеназа, цитохромы b, с1, с, цитохромоксидаза — комплекс цитохромов а и а3 (в них есть атомы меди).
3. Куда идут атомы водорода и электроны? Атомы водорода, электроны от НАДН и ФАДН2 отправляются на внутреннюю сторону мембраны митохондрии. Протоны идут в межмембранное пространство, лежащее между двумя мембранами митохондрий, наружной и внутренней, формируя протонный резервуар. А электроны атома водорода остаются на внутренней стороне мембраны, где они и концентрируются. Так создается разность потенциалов.
4. Протоны двигаются через канал в молекуле фермента АТФ-синтетазы
при достижении разности потенциалов определенной величины. АТФ-синтетазы встроены во внутренние мембраны митохондрий. Они образуют АТФ из АДФ. Энергия протонов при движении их через канал фермента расходуется на синтез АТФ. По мере образования АТФ протонный резервуар теряет свою энергию.
5. Внутри митохондрий катионы водорода Н+, соединяясь с кислородом и электронами, образуют воду: 2Н+ + 0,5О2 = Н2О.
6. Подведем итог. В цикле трикарбоновых кислот образуются НАДН, ФАДН2, СО2. При окислительном фосфорилировании образуются 34 АТФ, 6Н2О. В результате окисление одной молекулы глюкозы дает 38 АТФ, 6 СО2, 6Н2О.
1) КПД кислородного этапа — 55 процентов.
2) 45 процентов энергии теряется в виде тепла.
3) АТФ идет на химическую работу (биосинтез), механическую (мышцы), осмотическую (накопление и вывод веществ), электрическую (нервная ткань).
Метаболизм состоит из двух взаимно противоположных, но взаимосвязанных процессов пластического и энергетического обмена.
Энергетический обмен необходим организму для образования энергии, которая, в свою очередь, будет израсходована на важные биологические процессы, происходящие в клетках, тканях, органах, в том числе и на пластический обмен.
Все наши движения, мыслительные и физиологические процессы (пищеварение, кровообращение, выделение), любое проявление жизнедеятельности требуют затрат энергии.
Энергетический обмен также называют катаболизм или диссимиляцией. Это достаточно длительный процесс, который происходит вплоть до того момента, пока все питательные вещества, поступившие в организм, не расщепятся до углекислого газа, воды или других простых соединений, которые организм уже не сможет использовать.
Этот процесс аналогичен горению, при котором выделяется вода, углекислый газ и огромное количество энергии.
Катаболизм- это прежде всего многоступенчатый процесс, он не нуждается в высоких температурах, а выделившаяся энергия по большей части не переходит в тепловую, чтобы безвозвратно рассеяться, а запасается для дальнейших нужд в виде молекул АТФ.
Все это делает этот процесс невероятно эффективным и уникальным!
Эта информация доступна зарегистрированным пользователям
Энергетический обмен— это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ.
Каким же образом энергия реакции расщепления используется клеткой?
Ученые обнаружили, что любая деятельность клетки всегда точно совпадает во времени с распадом молекул АТФ.
К примеру, при синтезе белков, углеводов, жиров в клетке идет активный распад АТФ.
В результате опытов было обнаружено, что любая работа мышц сопровождается активным расщеплением АТФ в их клетках.
Ученые сделали вывод, что именно АТФ является непосредственным источником энергии, необходимой для сокращения мышц и для синтеза сложных соединений.
Известно, что в среднем содержание АТФ в клетках составляет от 0,05% до 0,5% ее массы, то есть запас молекул АТФ в организме ограничен и после распада АТФ должно произойти его восстановление.
Многоуровневый процесс энергетического обмена- это последовательные реакции восстановления молекул АТФ, которые происходят при участии ферментов.
Это можно сравнить с аккумулятором для телефона: когда его заряд садится, то устройство необходимо вновь зарядить.
Если в клетке постоянно измерять содержание АТФ, то его количество существенно не изменяется, но количество углеводов, белков, жиров будет уменьшаться. Это объясняется тем, что реакции расщепления углеводов, белков, жиров и других веществ обеспечивают быстрое и полное восстановление израсходованной АТФ.
В каждой клетке нашего организма в течение суток АТФ примерно 10 тысяч раз распадается и вновь заново образуется.
Таким образом, АТФ- это единый и универсальный источник энергии для функциональной деятельности клетки.
Следует отметить, что возможна передача энергии из одних частей клетки в другие.
Синтез АТФ может происходить в одном месте и в одно время, а использоваться может в другом месте и в другое время.
Синтез АТФ в основном происходит в митохондриях. Образовавшаяся там АТФ по каналам эндоплазматической сети направляется в те места клетки, где возникает потребность в энергии.
Эта информация доступна зарегистрированным пользователям
Это одно из проявлений высочайшей организованности и упорядоченности всех химических реакций, протекающих в клетке.
Растения могут преобразовывать энергию солнечных лучей в АТФ на первом этапе фотосинтеза. Хемосинтезирующие бактерии способны запасать энергию в форме АТФ, получаемую при реакциях окисления различных неорганических соединений.
Следует отметить, что фотосинтезирующие и хемосинтезирующие организмы также способны получать энергию благодаря окислению органических веществ, синтезированных в собственных клетках из неорганических соединений.
У гетеротрофов (животных, грибов) образование АТФ идет в клетках при помощи реакций окисления органических веществ, поступающих вместе с пищей.
В клетках растений:
Крахмал →глюкоза → АТФ
В клетках животных:
гликоген → глюкоза → АТФ
Энергетический обмен делится на три последовательных этапа:
- подготовительный этап
- бескислородный этап
- кислородный этап
Подготовительный этап
Вся пища, которая поступает в наш организм, подвергается ферментативному расщеплению, при котором:
- белки расщепляются до аминокислот
- липиды расщепляются до глицерина и жирных кислот
- сложные углеводы (крахмал) расщепляются до глюкозы
На этом этапе вся выделившаяся при расщеплении веществ энергия рассеивается в виде тепла.
Эта информация доступна зарегистрированным пользователям
У одноклеточных животных подготовительный этап протекает в клетках, где и происходит расщепление сложных органических веществ на простые вещества под действием ферментов лизосом.
У многоклеточных организмов расщепление веществ начинает происходить в пищеварительном канале, а далее в клетках под действием лизосом.
Эта информация доступна зарегистрированным пользователям
Образовавшиеся небольшие органические молекулы могут быть использованы в качестве «строительного материала» или могут подвергаться дальнейшему расщеплению (гликолизу).
Вывод: на первом этапе энергетического обмена происходит распад сложных органических веществ на простые, с выделением энергии, которая вся рассеивается в виде тепла.
Эта информация доступна зарегистрированным пользователям
Ключевое место в метаболизме всех типов клеток занимают реакции с участием сахаров, например, глюкозы, потому что процесс расщепления глюкозы идет наиболее быстро и легче, ведь организму необходимо достаточно быстро восстанавливать энергетические затраты.
Аминокислоты и белки использовать для образования энергии слишком не выгодно, так как большая их часть является структурными компонентами клеток. В этом случае организм разрушал бы сам себя.
Жиры могут использоваться для получения энергии, но главным образом после того, как израсходовались запасы углеводов, ведь жиры из-за своей гидрофобности очень медленно окисляются и малоподвижны в клетках. При этом из жиров в отсутствие кислорода АТФ получить нельзя, а из глюкозы можно.
Поэтому организм выбирает наиболее выгодный путь получения энергии в виде молекул АТФ за счет расщепления, в первую очередь, глюкозы.
Второй этап энергетического обмена называют бескислородным, так как процесс расщепления глюкозы и образования молекул АТФ идет без участия кислорода.
Гликолиз (от греч. «гликос» сладкий, «лизис»- расщепление) — последовательное расщепление глюкозы.
Гликолиз идет в цитоплазме клеток без участия кислорода. Он состоит из последовательных реакций, каждая из которых катализируется общим ферментом.
В ходе реакций гликолиза молекула глюкозы С6Н12О6 распадается на две трехуглеродные молекулы пировиноградной кислоты (ПВК)— С3Н4О3, при этом суммарно образуются две молекулы АТФ и вода.
Акцептором (лат. accipio- «я принимаю, получаю») водорода в реакции гликолиза служит кофермент НАД+.
НАД+ (никотинамидадениндинуклеотид) — кофермент, имеющийся во всех живых клетках.
НАД+ переносит электроны из одной реакции в другую.
НАД+ является окислителем и забирает электрон от другой молекулы и один водород, восстанавливаясь в НАД H, который далее служит восстановителем и уже отдаёт электроны.
Уравнение реакции гликолиза:
Эта информация доступна зарегистрированным пользователям
Дальнейшая судьба ПВК может быть различной и зависит от того, какой тип извлечения энергии предпочитают организмы: анаэробный (бескислородный) или аэробный (кислородный).
Например, паразитические черви, живущие в кишечнике организмов хозяев, выбирают бескислородный путь преобразования ПВК, так как они мало подвижны и их клеткам хватает энергии, которая образуется при гликолизе глюкозы.
Эти виды паразитов выбирают именно такой путь преобразования энергии еще и потому, что при распаде глюкозы образуются ядовитые вещества (ацетон, уксусная кислота и этиловый спирт), которые действуют угнетающе на организм хозяина и ослабляют его иммунитет, что, в свою очередь, помогает паразиту существовать в агрессивной для него среде.
Эта информация доступна зарегистрированным пользователям
Образовавшаяся в результате гликолиза пировиноградная кислота подвергается дальнейшему преобразованию уже на внутренней мембране митохондрий, то есть переходит на третий этап энергетического обмена.
Вывод: на втором этапе энергетического обмена, гликолизе, из 1 молекулы глюкозы образуется 2 молекулы ПВК и 2 молекулы АТФ.
Эта информация доступна зарегистрированным пользователям
Если в клетку прекратилась подача кислорода, то ПВК подвергается брожению, к примеру, в клетках растений, которые были затоплены во время весенних паводков.
В зависимости от того, какие конечные продукты образуются, выделяют несколько видов брожения.
Рассмотрим основные виды:
1. Спиртовое брожение
Встречается в основном у дрожжей и растений.
Конечными продуктами являются этанол и углекислый газ.
Дрожжи:
Эта информация доступна зарегистрированным пользователям
При доступе кислорода процесс брожения ослабевает, на смену ему приходит дыхание.
Подавление спиртового брожения кислородом называется эффектом Пастера.
Спиртовое брожение используется в пищевой промышленности: хлебопечении, виноделии.
При этом типе брожения сначала происходит образование уксусного альдегида, а затем этилового спирта:
Эта информация доступна зарегистрированным пользователям
2. Молочнокислое брожение
Осуществляется с помощью лактобактерий, бифидобактерий, стрептококков.
Из ПВК они образуют молочную кислоту, ацетон, янтарную и уксусную кислоту.
Молочнокислые бактерии широко используются в молочной промышленности для получения молочнокислых продуктов, а также в создании пробиотиков.
Эта информация доступна зарегистрированным пользователям
У животных и человека при недостатке кислорода также может происходить молочнокислое брожение с образованием молочной кислоты.
В мышцах есть запасы углеводов в виде гликогена. При долгой и усиленной работе кровь не успевает снабдить мышцы достаточным количеством кислорода, в результате чего мышечные клетки вынуждены переходить на бескислородный способ получения АТФ.
При этом образуется молочная кислота, вызывающая боли в мышцах.
Эта информация доступна зарегистрированным пользователям
Квашение- разновидность молочнокислого брожения, в процессе которого образуется молочная кислота, оказывающая на продукты (наряду с добавляемой поваренной солью) консервирующее и размягчающее действие.
Квашение применяется при консервировании овощей и в кожевенном производстве.
Эта информация доступна зарегистрированным пользователям
3. Маслянокислое брожение
Масляная кислота, бутанол, ацетон, уксусная и ряд других органических кислот являются продуктами сбраживания углеводов бактериями- сахаролитическими анаэробами.
Благодаря определению наличия тех или иных кислот в клетке можно установить, какие бактерии образовали эти кислоты.
Знание механизмов брожения имеет большое практическое значение не только для живых организмов, но и для человека:
- для разработки методов диагностики инфекционных заболеваний, по набору ферментов или кислот, которые образовались во время брожения
- для создания современных биотехнологий молочнокислых продуктов, сыра, хлеба, вина и многих других продуктов питания
Недостатком процессов брожения является извлечение незначительной доли той энергии, которая заключена в связях органических молекул.
Для бактерий, паразитических видов, живущих в бескислородной среде, энергии, образующейся в результате брожения или гликолиза, достаточно для существования, поэтому они, в отличие от человека, не нуждаются в кислороде.
Также брожение является жизненно важным процессом для хвойных растений. В зимний период устьица хвои закупориваются смолой и газообмен с окружающей средой практически прекращается, в этом случае для получения энергии в клетках активно идет процесс спиртового брожения.
Эта информация доступна зарегистрированным пользователям
Эта информация доступна зарегистрированным пользователям
Окислительное фосфорилирование (дыхание)- процесс синтеза АТФ с обязательным участием комплекса ферментов, идет на внутренних мембранах митохондрий в присутствии кислорода.
Эта информация доступна зарегистрированным пользователям
Для нормального течения кислородного процесса необходима целостность мембран митохондрий.
В процессе такого клеточного дыхания энергия может переходить из химической в тепловую.
Кислородное дыхание гораздо эффективнее гликолиза, так как полное окисление органических веществ приводит к выделению большого количества энергии, причем примерно 60% ее запасается в молекулах АТФ, а 40% рассеивается в виде тепла.
Образовавшаяся в процессе гликолиза пировиноградная кислота поступает в митохондрии. Здесь она превращается в богатое энергией вещество ацетилкофермент А (Ацетил-КоА).
Ацетил-КоА взаимодействует с молекулой щавелевоуксусной кислоты, образуя лимонную кислоту, которая подвергается дальнейшим превращениям, заканчивающимся образованием щавелевоуксусной кислоты.
Эта кислота вновь взаимодействует с Ацетил-КоА, и цикл превращений повторяется.
Этот сложный процесс получил название цикл Кребса или цикл трикарбоновых кислот, еще его называют «циклом лимонной кислоты».
Цикл превращения трикарбоновых кислот открыл немецкий биохимик Ханс Кребс (1900- 1981), за что в 1953 году совместно с другими исследователями он получил Нобелевскую премию.
В цикле Кребса образуются молекулы СО2 (выводятся из клетки) и атомы водорода Н.
Специальные молекулы-переносчики НАД доставляют атомы Н к внутренней мембране митохондрий, имеющей внутреннюю и наружную поверхности.
Здесь атомы Н теряют электрон и превращаются в протоны Н+
Н — ē → Н+
Свободные электроны перемещаются по цепи переноса электронов на внутреннюю поверхность внутренней мембраны и присоединяются к атому кислорода О2:
О2 + ē= О2—
Далее 4 протона Н+соединяются с О2—в результате образуются две молекулы воды, которую еще называют метаболической водой, то есть выработанной организмом, а не полученной извне.
4Н+ + О2— = 2 Н2О + Q(т)
Обратите внимание, что в данной реакции образуется тепловая энергия (Qт)
Эта информация доступна зарегистрированным пользователям
Сиреневой стрелочкой показано, как белки дыхательной цепи забирают электроны у НАД Н.
Оказывается, белки дыхательной цепи за счет разницы энергии входящего электрона и выходящего могут протаскивать сквозь мембрану протоны Н+ в пространство между мембранами, где формируется положительный заряд.
Таким образом, между внутренней и наружной поверхностями внутренней мембраны митохондрий возникает разность потенциалов и формируется большая потенциальная энергия, за счет которой происходит образование АТФ.
Красная стрелка показывает, как белки дыхательной цепи переносят протоны Н+ в пространство между мембранами.
АТФ- синтаза (фермент, синтезируюший АТФ) пропускает протоны Н+ через свой активный центр и за счет этой энергии из АДФ и фосфорной кислоты образуется молекула АТФ.
В матриксе митохондрий протоны Н+соединяются с анионами О-2, образуя воду (Н2О).
АДФ + Ф → АТФ+ Н2О
Таким образом, в процессе дыхания в митохондриях образуются бедные энергией вещества: СО2 и Н2О, и освобождается большое количество энергии.
При полном расщеплении двух молекул ПВК, (синтезированных из одной молекулы глюкозы в процессе гликолиза), образуется 36 молекул АТФ.
Для большинства животных процессы дыхания являются основным путем получения энергии, необходимой для их жизнедеятельности.
ПВК + О2 → СО2 + Н2О + Qт (энергия)
Эта информация доступна зарегистрированным пользователям
Организм человека функционирует в аэробных условиях: 90% энергии он получает при участии кислорода.
Итак, повторим этапы энергетического обмена:
|
Подготовительный этап |
Бескислородный этап (гликолиз) |
Кислородный этап |
|
|
Где происходит |
В органах пищеварения, в клетках под действием ферментов |
Внутри клетки, в цитоплазме |
На внутренних мембранах митохондрий |
|
На какие вещества расщепляются соединения клетки |
Белки до аминокислот. Жиры до глицерина. Жирные кислоты и углеводы до глюкозы |
Глюкоза (С6Н12О6) до: ПВК (С3Н4О3) Энергия (Qт) |
ПВК (С3Н4О3) до: СО2 Н2О |
|
Сколько выделяется энергии |
Мало, рассеивается в виде тепла. |
40% энергии идет на синтез АТФ. 60% рассеивается в виде тепла |
Более 60% энергии запасается в виде АТФ |
|
Сколько синтезируется энергии в виде АТФ |
Не образуется |
2 молекулы АТФ |
36 молекул АТФ |
Общий вывод: объединяя анаэробные и аэробные процессы образования АТФ при расщеплении глюкозы, можно записать следующее суммарное уравнение:
Эта информация доступна зарегистрированным пользователям
Эта информация доступна зарегистрированным пользователям
Задача 1
В процессе гликолиза в клетках эукариот образовались 120 молекул пировиноградной кислоты (ПВК).
Какое количество молекул глюкозы подверглось расщеплению и сколько молекул АТФ образуется при полном ее окислении? Ответ поясните.
Решение:
1) В процессе гликолиза при расщеплении 1 молекулы глюкозы образуются 2 молекулы пировиноградной кислоты и выделяется энергия, которой хватает на синтез двух молекул АТФ
2) Так как образовалось 120 молекул пировиноградной кислоты, то расщеплению подверглось 120 : 2 = 60 молекул глюкозы
3) При полном окислении в расчете на одну молекулу глюкозы образуется 38 молекул АТФ
4) Следовательно, при полном окислении 60 молекул глюкозы образуются
38 х 60 = 2280 молекул АТФ
Задача 2
Гликолизу подверглись две молекулы глюкозы, окислению только одна.
Определите количество образовавшихся в результате этого процесса молекул АТФ и углекислого газа.
Решение:
Для решения используем уравнения 2 этапа (гликолиза) и 3 этапа (кислородного) энергетического обмена.
1) При гликолизе одной молекулы глюкозы образуются 2 молекулы АТФ
2) По условию задачи гликолизу подверглось 2 молекулы глюкозы:
2х2= 4 молекулы АТФ образовалось
3) При окислении (кислородный этап) из одной молекулы глюкозы образуются 36 молекул АТФ
4) По условию задачи окислению подверглась только одна молекула глюкозы:
1 х 36= 36 молекул АТФ образовалось
5) Таким образом образовалось 4 + 36 = 40 молекул АТФ
6) Углекислый газ образуется только на 3 этапе, при полном окислении одной молекулы глюкозы образуется 6 молекул СО2
Ответ: образовалось 40 молекул АТФ и 6 молекул СО2
Задача 3
В процессе окислительного фосфорилирования образовалось 1728 молекул АТФ.
Определите, какое количество молекул глюкозы подверглось расщеплению и сколько молекул АТФ образовалось в результате подготовительного и бескислородного этапов катаболизма.
Решение:
1) При окислительном фосфорилировании (третий этап энергетического обмена) из одной молекулы глюкозы образуется 36 молекул АТФ, следовательно, гидролизу подверглось:
1728 : 36 = 48 молекул глюкозы
2) При гликолизе она молекула глюкозы расщепляется до двух молекул пировиноградной кислоты (ПВК) с образованием двух молекул АТФ, следовательно, образовалось:
48 х 2 = 96 молекул АТФ
3) При полном окислении одной молекулы глюкозы образуется 38 молекул АТФ, следовательно, при полном окислении 48 молекул глюкозы образовалось:
48 х 38 = 1824 молекулы АТФ (1728 АТФ третьего этапа + 96 АТФ второго этапа = 1824)
Задача 4
Сколько молекул АТФ будет синтезировано в клетках в процессе гликолиза, если происходит окисление участка молекулы крахмала, содержащего 50 остатков глюкозы?
Решение:
1) В процессе гликолиза при окислении одной молекулы глюкозы образуется 2молекулы АТФ
2) Из фрагмента молекулы крахмала, содержащего 50 остатков глюкозы в результате гликолиза образуется 100 молекул АТФ (50 х 2 = 100)
Задача 5
В процессе полного расщепления глюкозы образовалось 342 молекулы АТФ.
Сколько молекул глюкозы поверглось расщеплению?
Сколько молекул АТФ образовалось в результате гликолиза?
Объясните полученный результат.
Решение:
1) В результате полного расщепления одной молекулы глюкозы образуется 38 молекул АТФ
2) Расщеплению поверглись 342 : 38 = 9 молекул глюкозы
3) Гликолиз– бескислородный этап катаболизма, в результате которого из одной молекулы глюкозы образуется 2 молекулы АТФ
4) Следовательно, в результате гликолиза образовалось 9 х 2 = 18 молекул АТФ
Задача 6
При беге со средней скоростью мышцы ног расходуют за 1 минуту 24 кДж энергии.
Определите сколько граммов глюкозы израсходуют мышцы ног за 25 минут бега, если кислород доставляется кровью к мышцам в достаточном количестве?
Для сведения: один моль содержит 180 грамм глюкозы и из этого количества образуется 38 молекул АТФ.
Решение:
1) Определим сколько всего энергии было израсходовано за 25 минут бега:
25 минут х 24 кДж = 600 кДж энергии
2) Определим сколько молекул АТФ образовалось, учитывая, что 1 молекула АТФ образует 40 кДж энергии:
600 : 40 = 15 молекул АТФ
3) Определим сколько глюкозы было израсходовано. Из условия ясно, что 1 моль глюкозы образует 38 молекул АТФ:
С6Н12О6 + 6 О2→ 6 СО2 + 6 Н2О + 38 АТФ
Составляем пропорцию:
1 моль (C6H12O6) — 38 (АТФ)
Х моль (C6H12O6) — 15 (АТФ)
Х=1 х 15/38
Х= 0,4 моль (C6H12O6)
4) Определяем, сколько грамм глюкозы содержится в 0,4 молях, для этого опять составляем пропорцию:
1 моль (C6H12O6) = 180 гр
0,4 моль (C6H12O6) = Х гр
Х=180 х 0,4/1
Х = 72 гр (C6H12O6)
Ответ:71 гр глюкозы израсходуют мышцы при беге продолжительность 25 минут
Эта информация доступна зарегистрированным пользователям
Читайте также
Цикл окислительного фосфорилирования
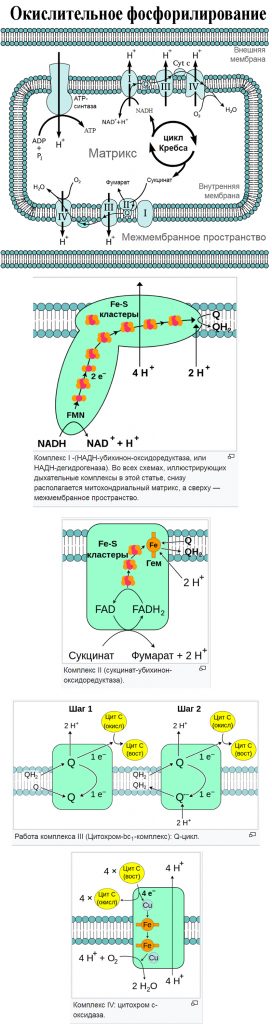 Окислительное фосфорилирование — метаболический путь, при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях клеток в виде АТФ. Хотя различные формы жизни на Земле используют разные питательные вещества, АТФ является универсальным соединением, в котором запасается энергия, необходимая для других метаболических процессов. Почти все аэробные организмы осуществляют окислительное фосфорилирование. Вероятно, широкому распространению этого метаболического пути способствовала его высокая энергетическая эффективность по сравнению с анаэробным брожением.
Окислительное фосфорилирование — метаболический путь, при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях клеток в виде АТФ. Хотя различные формы жизни на Земле используют разные питательные вещества, АТФ является универсальным соединением, в котором запасается энергия, необходимая для других метаболических процессов. Почти все аэробные организмы осуществляют окислительное фосфорилирование. Вероятно, широкому распространению этого метаболического пути способствовала его высокая энергетическая эффективность по сравнению с анаэробным брожением.
При окислительном фосфорилировании происходит перенос электронов от соединений-доноров к соединениям-акцепторам в ходе окислительно-восстановительных реакций. В ходе этих реакций выделяется энергия, которая далее запасается в виде АТФ. У эукариот эти окислительно-восстановительные реакции осуществляются несколькими белковыми комплексами, локализованными во внутренней митохондриальной мембране, а у прокариот они располагаются в межмембранном пространстве клетки. Этот набор связанных белков составляет электроно-транспортную цепь (ЭТЦ). У эукариот в состав ЭТЦ входит пять белковых комплексов, в то время как у прокариот её составляют множество различных белков, работающих с различными донорами и акцепторами электронов.
Электронно – транспортная цепь митохондрий является местом окислительного фосфорилирования у эукариот. НАДФ и сукцинат, образовавшиеся в ходе цикла Кребса окисляются, и их энергия передается АТФ-синтетазе, которая за счет этой энергии синтезирует АТФ.
Эта тема входит в мой курс по биохимии
Окислительное фосфорилирование — это заключительный этап общего катаболизма. Напомню, что он включает: окислительное декарбоксилирование пирувата, цикл Кребса и окислительное фосфорилирование. Про первые два говорили здесь.
Окислительное фосфорилирование — сложный процесс. Он включает три этапа: перенос электронов по дыхательной цепи, создание градиента концентрации протонов и синтез АТФ. Не слишком понятно, не правда ли? Ничего, сейчас разберёмся. Но сначала нужно вспомнить кое-что.
Общий путь катаболизма идёт в митохондрии. Нам нужно разобраться со строением мембран митохондрии, потому что без этого мы далеко не уедем. У митохондрии есть две мембраны: наружная и внутренняя. В наружной мембране есть поры, поэтому она проницаема для небольших молекул, в том числе ионов. Внутренняя мембрана непроницаема для любых молекул — там нет пор. Тут я загнул, всё-таки есть жирорастворимые молекулы, которым на мембрану плевать.
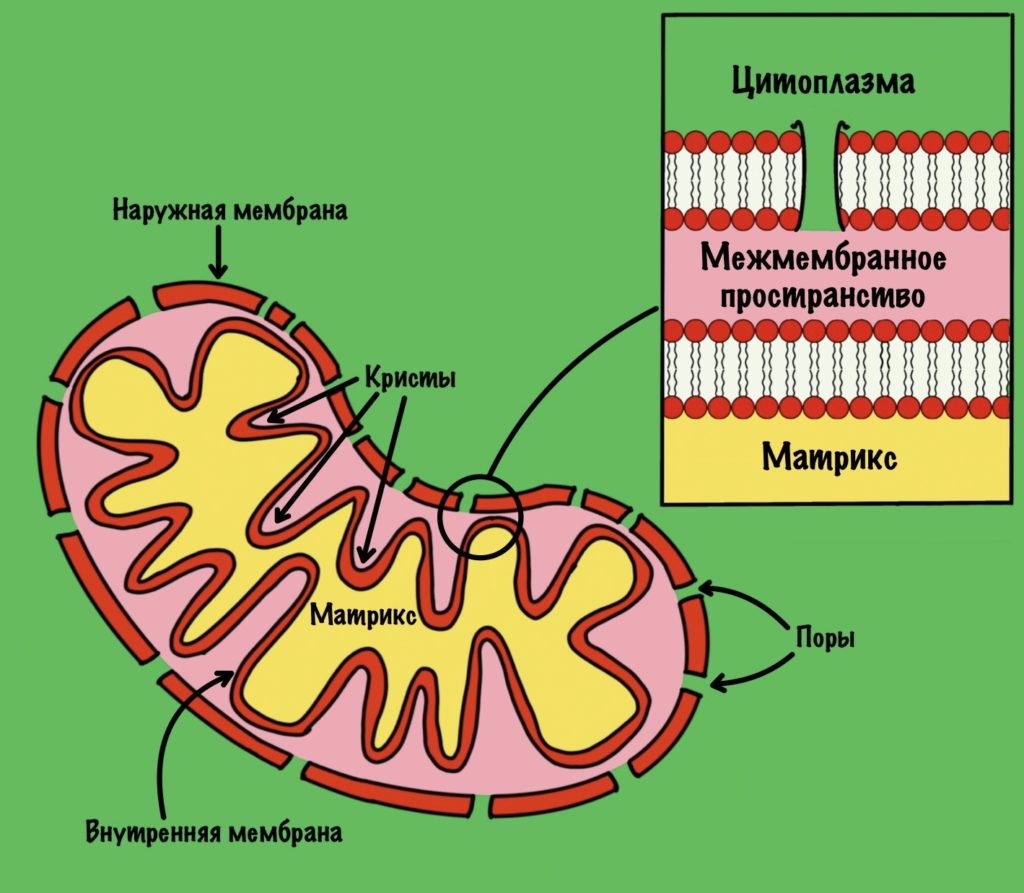
Внутренняя мембрана увеличивает свою площадь за счет формирования крист. Видите, какая она неровная? Матрикс — это пространство между внутренней мембраной. А между внутренней и наружной мембранами находится мемжмембранное пространство.
Зачем нужно было это вспомнить? Потому что во внутренней мембране находится дыхательная цепь (или цепь переноса электронов). Чем больше площадь этой мембраны, тем больше в ней комплексов дыхательной цепи. Да-да, она состоит из нескольких комплексов, но пока что нужно запомнить вот это:
Внутренняя мембрана непроницаема для молекул
Можно обойти этот запрет. Для этого нужны белки переносчики, которые перекинут молекулу на другую сторону.
Окислительное фосфорилирование
Теперь займёмся окислительным фосфорилированием. Какая была основная суть у катаболизма? Вытащить максимальное количество электронов из молекулы — с этим он отлично справился, если вспомнить катаболизм одной молекулы глюкозы, то в ходе неё были получены 24 электрона — в виде НАДH и ФАДH2. Из них: четыре в гликолизе, четыре в окислительном декарбоксилировании пирувата и шестнадцать в цикле трикарбоновых кислот.
Зачем это было нужно? Для окислительного фосфорилирования, а если точнее, то одной из его частей — цепи переноса электронов. Мы договорились, что она находится во внутренней мембране митохондрий. Дыхательная цепь состоит из четырёх комплексов, но пока не будем лезть в дебри. Сейчас просто разберёмся с механизмом окислительного фофорилирования.
Электроны, которые были получены в ходе катаболизма, попадают на комплексы дыхательной цепи. Там они перескакивают с одного комплекса на другой, пока не попадут на кислород. В ходе таких перескоков создаётся энергия. Сразу скажу, что её не слишком много. Но достаточно для того, чтобы транспортировать протоны из матрикса в межмембранное пространство. За счёт этого создаётся градиент концентрации протонов. Протоны стараются попасть обратно в матрикс митохондрий, но мембрана для них непроницаема. Так они и бьются в мембрану, пока не попадают на АТФ-синтазу. Можно подумать, что АТФ-синтаза это канал для протонов, но это не совсем так. Транспорт протонов из межмембранного пространства в матрикс создаёт большое количество энергии. Эта энергия используется АТФ-синтазой для синтеза АТФ. Чем-то напоминает вторично-активный транспорт.
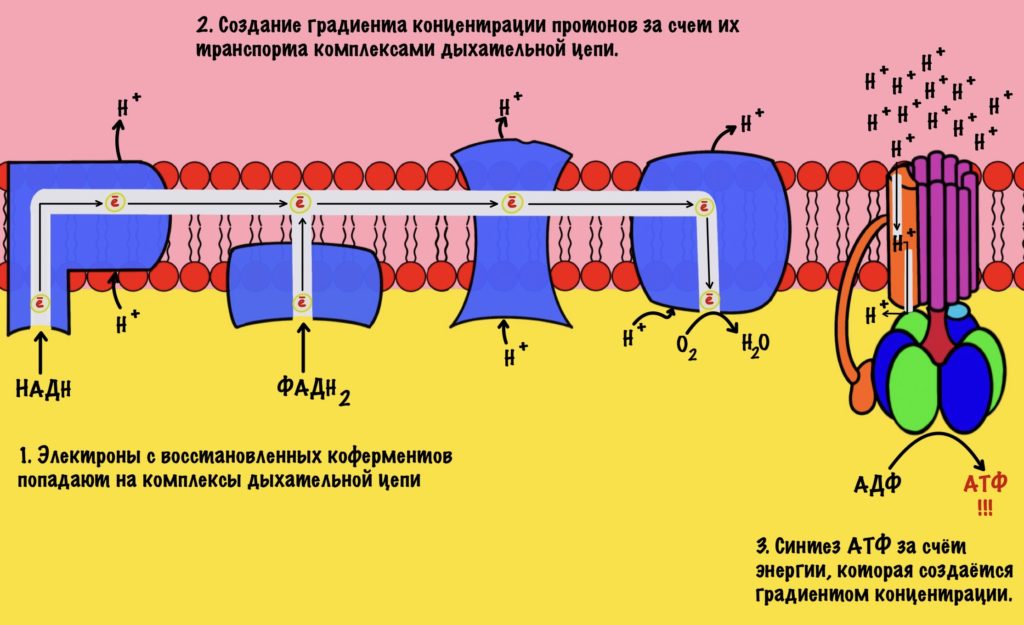
Представьте себе шланг. Вода под большим давлением идёт по шлангу, пока не дойдёт до его конца. Чем больше давление в шланге, тем сильнее из него вырывается вода. Примерно такой же принцип в работе окислительного фосфорилирования. Только напор воды создаёт дыхательная цепь, а единственное место выхода для неё — это АТФ-синтаза. Вода — это протоны водорода.
Из этого можно сделать вывод. Если мы не создадим градиент концентрации, то синтеза АТФ не будет. Получается, что работа цепи переноса электронов сопряжена с работой АТФ-синтазы. Это и есть окислительное фосфорилирование. В нём сопряжены три процесса: транспорт электронов по дыхательной цепи, транспорт протонов в межмембранное пространство и синтез АТФ. За первые два процесса отвечает дыхательная цепь, а за последний — АТФ-синтаза. Теперь, когда мы уяснили себе этот принцип, можно лезть в дебри!
Синтез АТФ сопряжён с работой дыхательной цепи
Дыхательная цепь состоит из четырёх комплексов
Комплексы — это гигантские белки. Они пронизывают внутреннюю мембрану митохондрий. Первый и второй комплекс принимают на себя электроны от восстановленных коферментов: НАДH и ФАДH2. При этом на первый комплекс попадают электроны с НАДH, а на второй комплекс с ФАДH2. Дальше мы поговорим об этом подробнее. Пока давайте ответим на простой вопрос: «Как заставить электроны перемещаться по дыхательной цепи?».
Электронам не выгодно прыгать с одного комплекса на другой. Им нет дела до создания градиента концентрации протонов. Митохондрии нужно заставить их перемещаться вдоль этой цепи. Для этого в комплексах дыхательной цепи находятся: коферменты, железо-серные белки, цитохромы и медь. Их строение будет разобрано далее. Но как они заставляют электроны передвигаться? С помощью редокс потенциалов. У них есть и другое название — окислительно-восстановительные потенциалы.
Дело тут такое, у молекул различное сродство к электронам. Чем больше окислительно-восстановительный потенциал молекулы, тем больше у неё сродство к электронам. Если расположить молекулы в порядке возрастания их потенциалов, то мы заставим двигаться электроны вдоль дыхательной цепи. На каждом комплексе у нас будет молекула, которая любит электроны больше, чем на предыдущем. Это заставит их работать! Вот редокс потенциалы для разных молекул.

Мы видим, что максимальный редокс потенциал у кислорода. У остальных молекул поменьше. Они будут располагаться в дыхательной цепи именно в такой последовательности — возрастания окислительно-восстановительных потенциалов.
У разных молекул — разные редокс потенциалы.
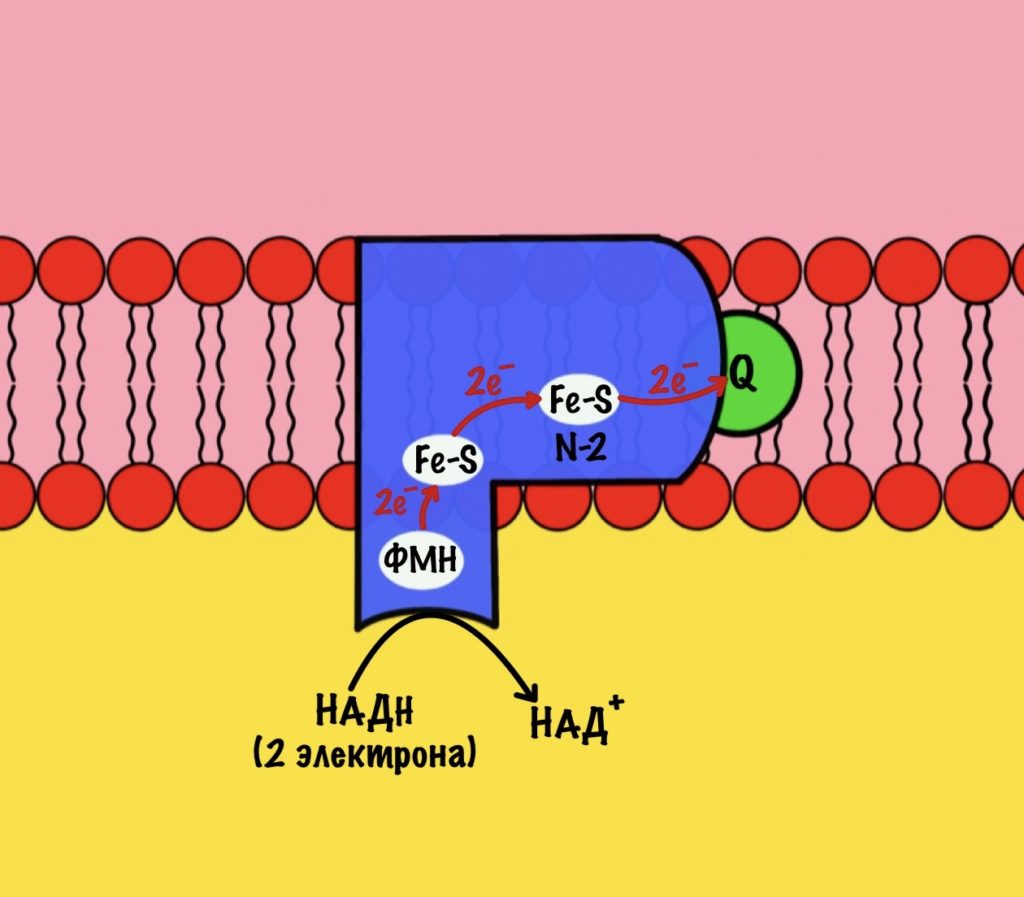
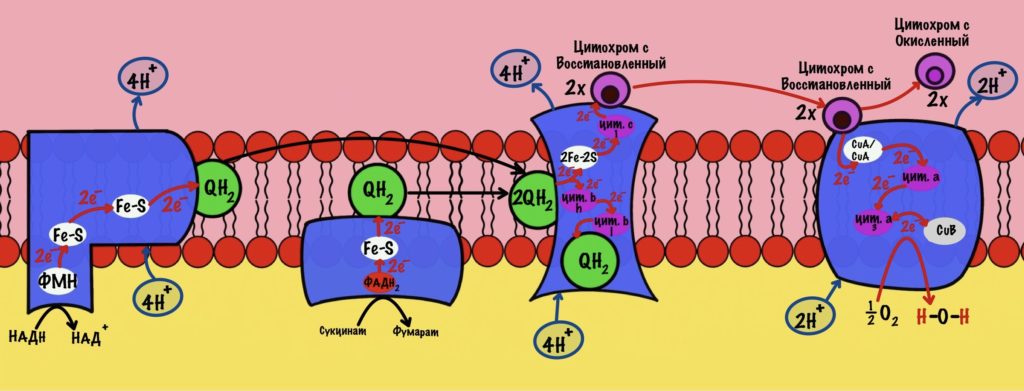
Комплекс 1 — НАДH-дегидрогеназа
Этот комплекс состоит из 45 полипептидных цепей! В его состав входит ФМН и железо-серные белки. Его основная задача — принять электроны от НАДH и переместить на убихинон (коэнзим Q10). Комплекс похож на перевёрнутую букву L: часть выходит в матрикс, а другая находится в мембране.
Восстановленный НАДH — из общего катаболизма — отдаёт гидрид ион (два электрона и протон) этому комплексу. Электроны сначала попадают на ФМН, а потом на железо-серные белки — всего их восемь. Я не буду рисовать их все, а нарисую только пару, последний — N-2.
Давайте посмотрим на строение железо-серных белков. Они могут включать один атом железа, два или четыре. Самые простые включают один атом железа — оно связано с четырьмя -SH группами. Железо-серные белки c двумя и четырьмя атомами железа связаны не только с остатками цистеина, но и неорганической серой.
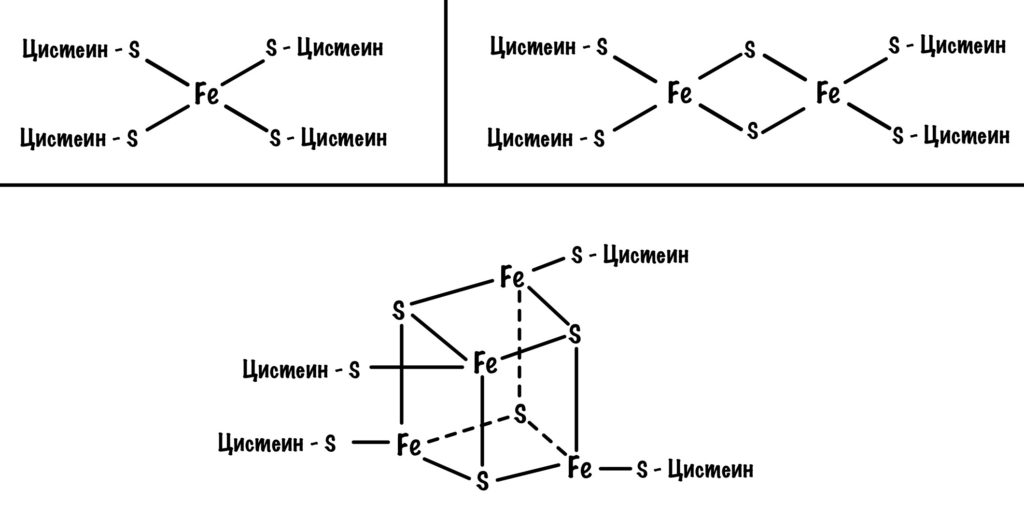
Железо в этих белках имеет заряд +3, но оно может присоединить электрон и перейти в +2. Это и происходит, когда электроны передвигаются от одного железно-серного белка к другому. Чем больше атомов железа в таких белках, тем больше электронов они могут присоединить к себе. Они что-то вроде склада электронов.
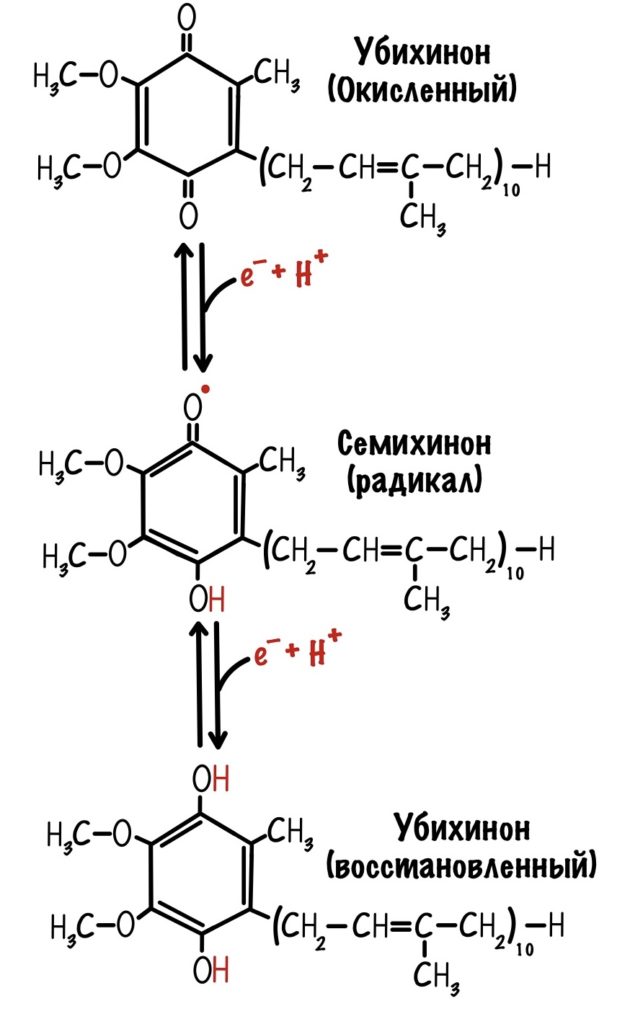
Последний железо-серный белок — N-2. Этот белок отдаёт электроны на убихинон. Убихинон — это жирорастворимая молекула, которая находится во внутренней мембране. Она присоединяется к первому комплексу и ждёт, пока комплекс отдаст ей электроны. Также он присоединяет два протона — один от НАДH, а другой из матрикса митохондрий. Вот как выглядит эта молекула. Убихинон переходит в восстановленную форму.
Мы видим то, о чём говорили в начале — серию окислительно-восстановительных реакций. Электроны из НАДH, через серию переносчиков первого комплекса, попадают на коэнзим Q10. В ходе этой серии реакций выделяется энергия. Энергия идёт на транспорт четырёх протонов из матрикса в межмембранное пространство. Точный механизм транспорта неизвестен, но он подтверждён практически. Так что первый комплекс — протонная помпа.
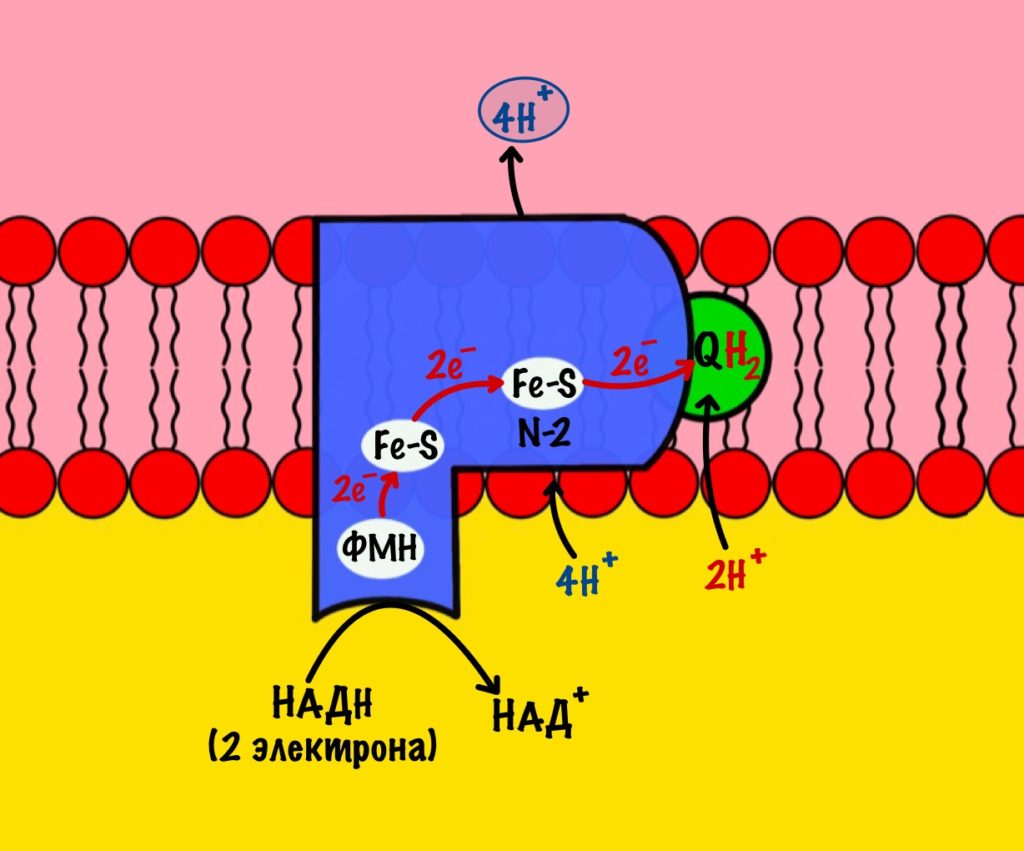
Первый комплекс переносит два электрона с НАДH на коэнзим Q10. Энергия окислительно-восстановительных реакций используется на транспорт четырёх протонов из матрикса в межмембранное пространство.
Далее убихинон отсоединяется от первого комплекса и путешествует по внутренней мембране. Он жирорастворимый, поэтому это не составляет ему труда. Восстановленный убихинон присоединится к третьему комплексу, которому и отдаст свои электроны и протоны.
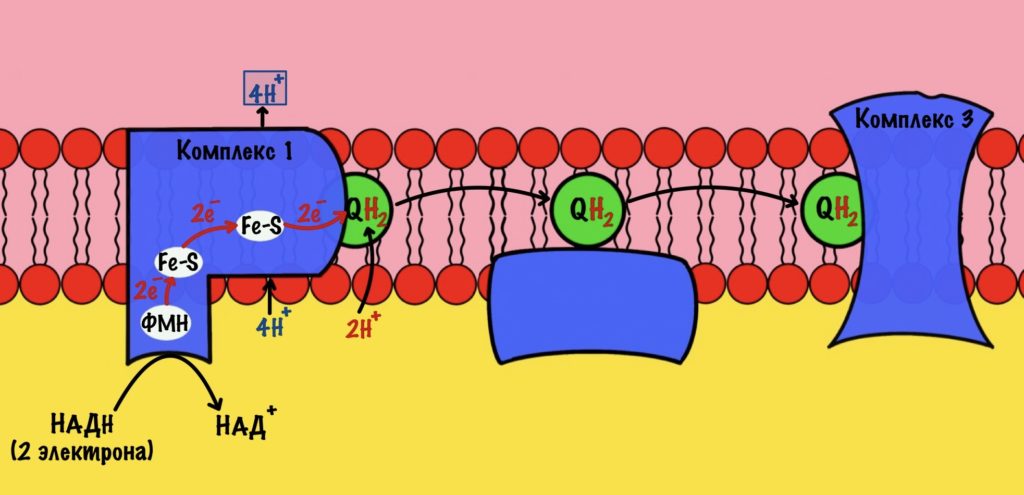
Стоп, а где второй комплекс? Их же всего четыре, почему убихинон переносит электроны сразу на третий? А сейчас разберёмся.
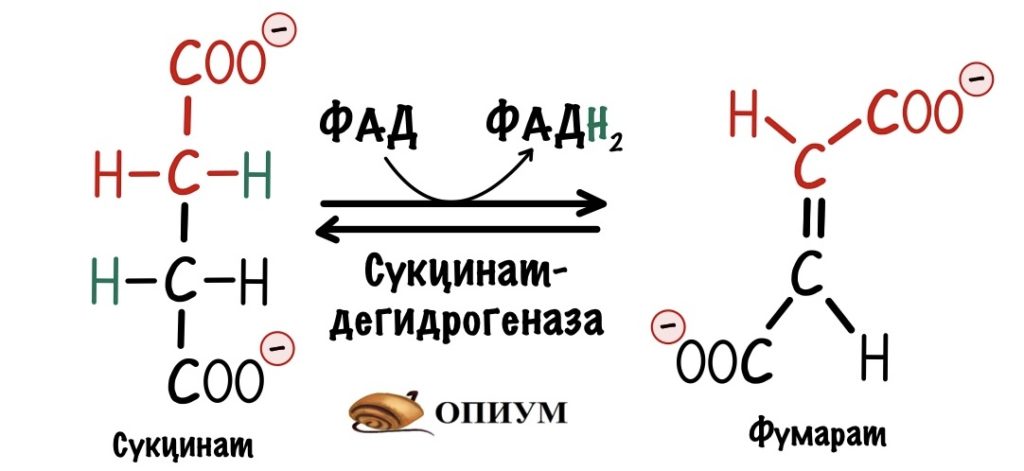
Второй комплекс — сукцинатдегидрогеназа
Мы уже встречались с ним — в реакциях цикла Кребса! Сукцинатдегидрогеназа катализировала превращение сукцината в фумарат. Кофермент этой реакции — ФАД. Это единственный фермент из цикла трикарбоновых кислот, который крепится к внутренней мембране митохондрий.
Задача второго комплекса — транспорт электронов с ФАДH2 их на убихинон. Второй комплекс намного меньше первого, он включает четыре субъединицы. Только одна ответственна за реакцию сукцинатдегидрогеназы — субъединица А. Кофермент ФАД тесно связан с этим комплексом. Остальные субъединицы транспортируют электроны. В комплексе есть три железо-серных белка и цитохром b. Железо-серные белки переносят электроны на убихинон. Цитохром b не участвует в транспорте электронов, он нужен для избежания утечек электронов. Иногда электроны сбиваются со своего пути, а это может привести к образованию токсичных форм кислорода. Это радикалы, которые могут натворить много неприятных дел внутри клетки. Так что цитохром b на подсейве.
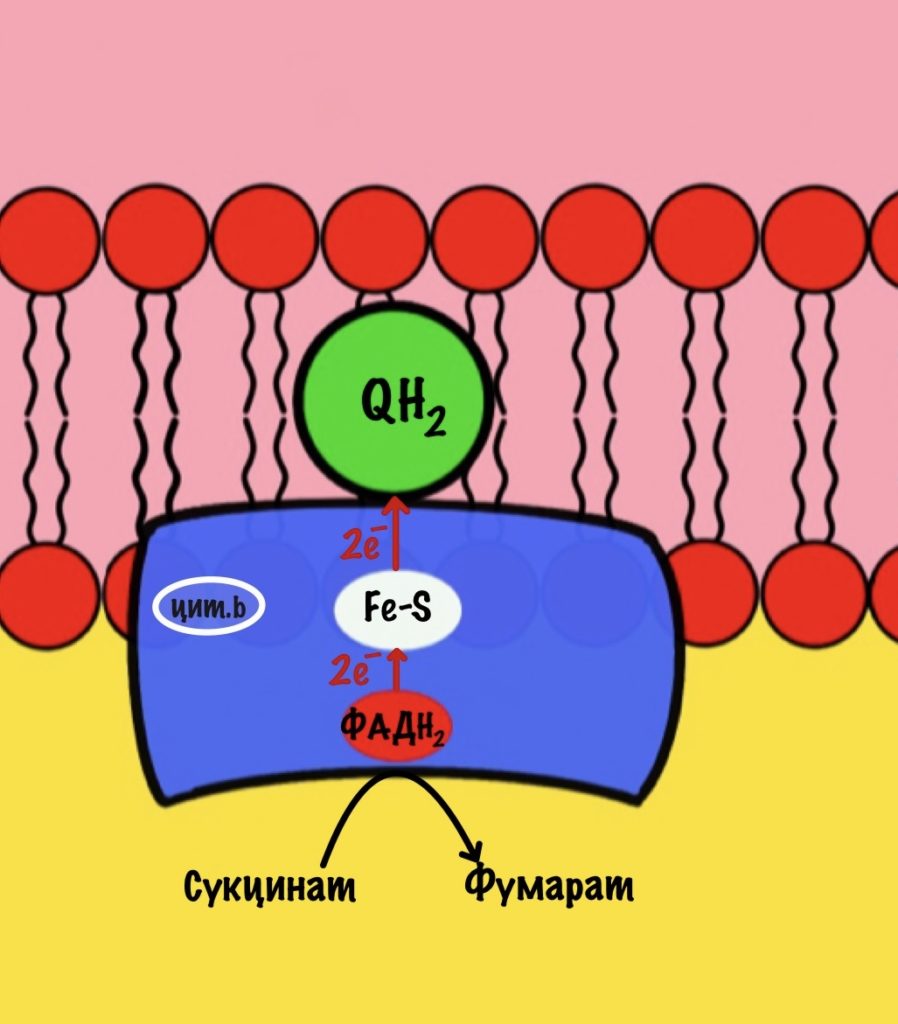
Убихинон принимает электроны с ФАДH2 не только от сукцинатдегидрогеназной реакции. Например, он может присоединять электроны, которые образовались в глицеролфосфатном челноке.
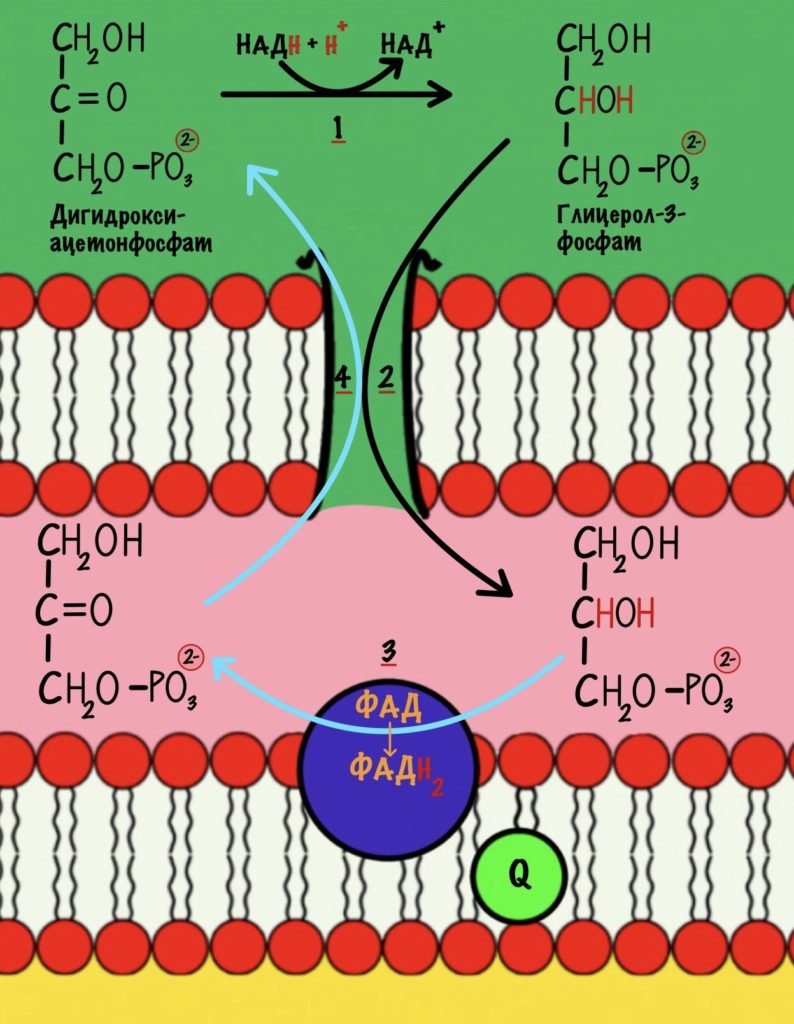
Второй комплекс не работает, как протонная помпа. Он не транспортирует протоны из матрикса в межмемрбанное пространство.
Второй комплекс переносит два электрона от ФАДH2 на убихинон. Энергия окислительно-восстановительных реакций не используется на транспорт протонов.
Дальше убихинон будет путешествовать по мембране, пока не присоединится к третьему комплексу. Там он отдаст электроны и протоны.
Третий комплекс — цитохром bc1 комплекс
Мы ещё не встречались с цитохромами, что это такое? Цитохромы — это белки, которые содержат в себе гем. Гем состоит из порфирина, который соединен с железом. Порфирин состоит из четырёх пятичленных колец. В каждом кольце есть атом азота, с помощью которого кольца соединяются с атомом железа. У порфирина могут быть разные радикалы, по ним цитохромы делятся на разные типы.
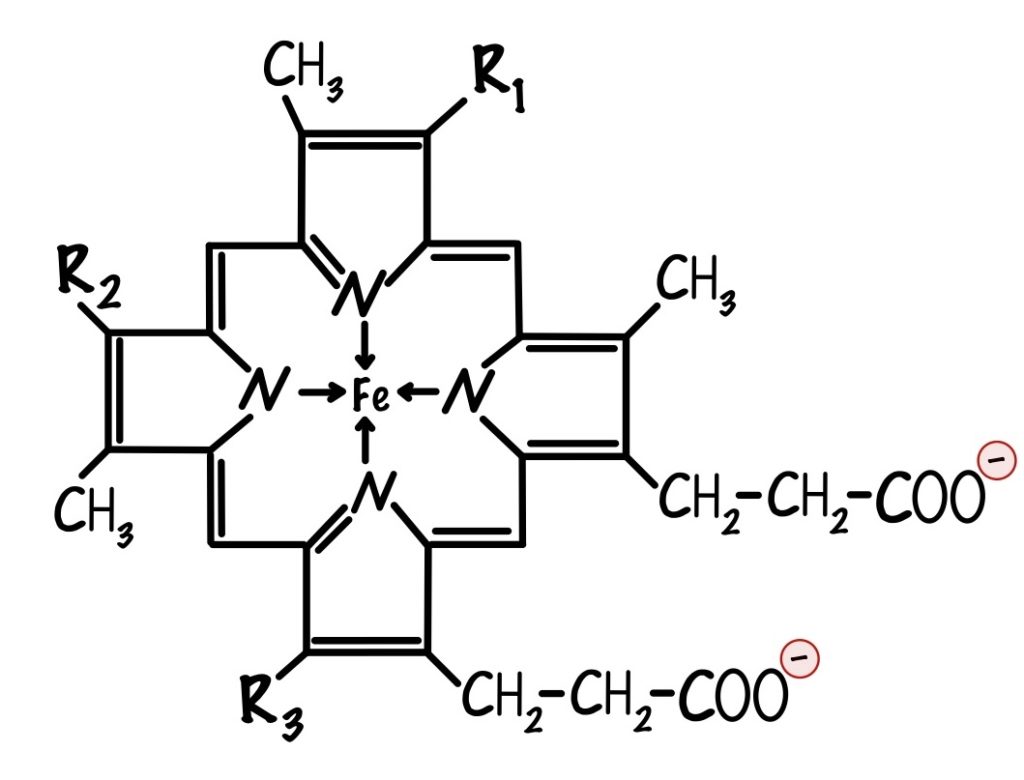
Важно понять, что в цитохромах есть один атом железа. Он может принимать электрон и менять степень окисления: из +3 в +2. Цитохромы могут принимать только один электрон, потому что в них один атом железа.
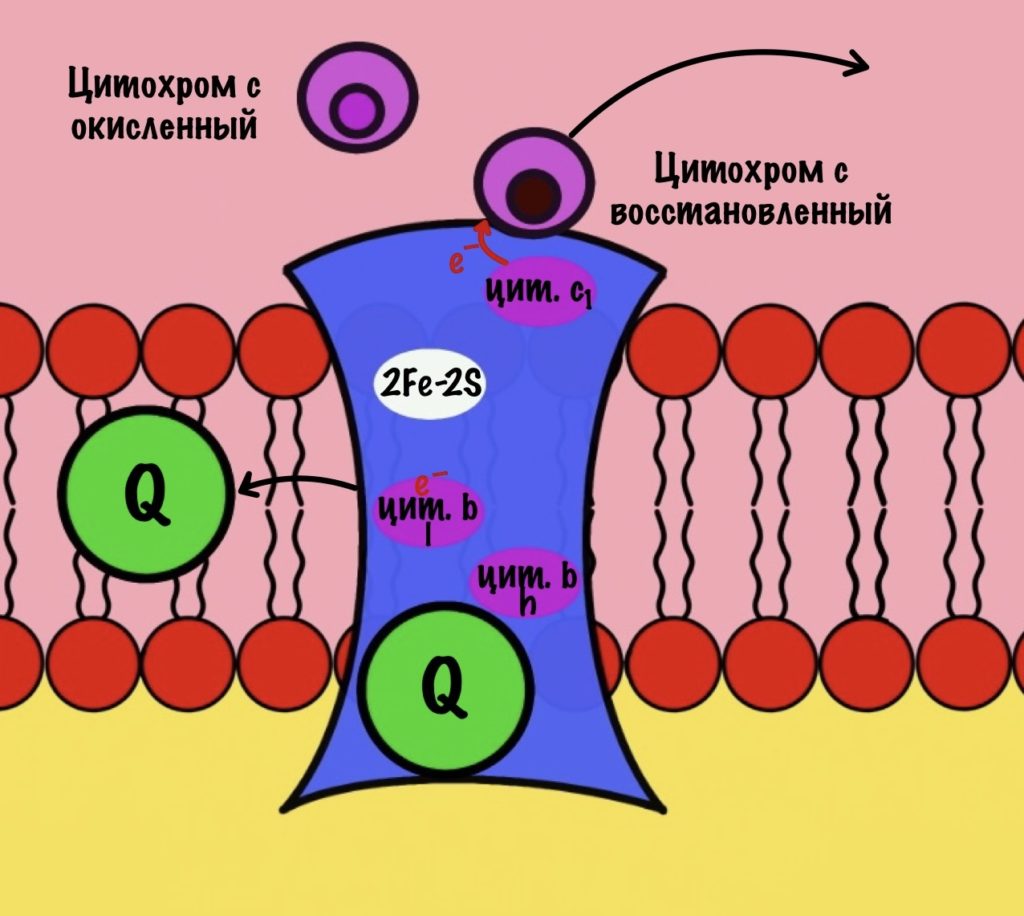
Третий комплекс состоит из двух субъединиц. В каждой субъединице есть: 2Fe-2S железо-серный белок, цитохром b, цитохром c1. В цитохроме b есть два порфириновых кольца, поэтому его делят на l и h. К субъединицам крепится цитохром c — он находится в межмембранном пространстве митохондрии. Цитохром c растворим в воде, в отличие от остальных цитохромов.
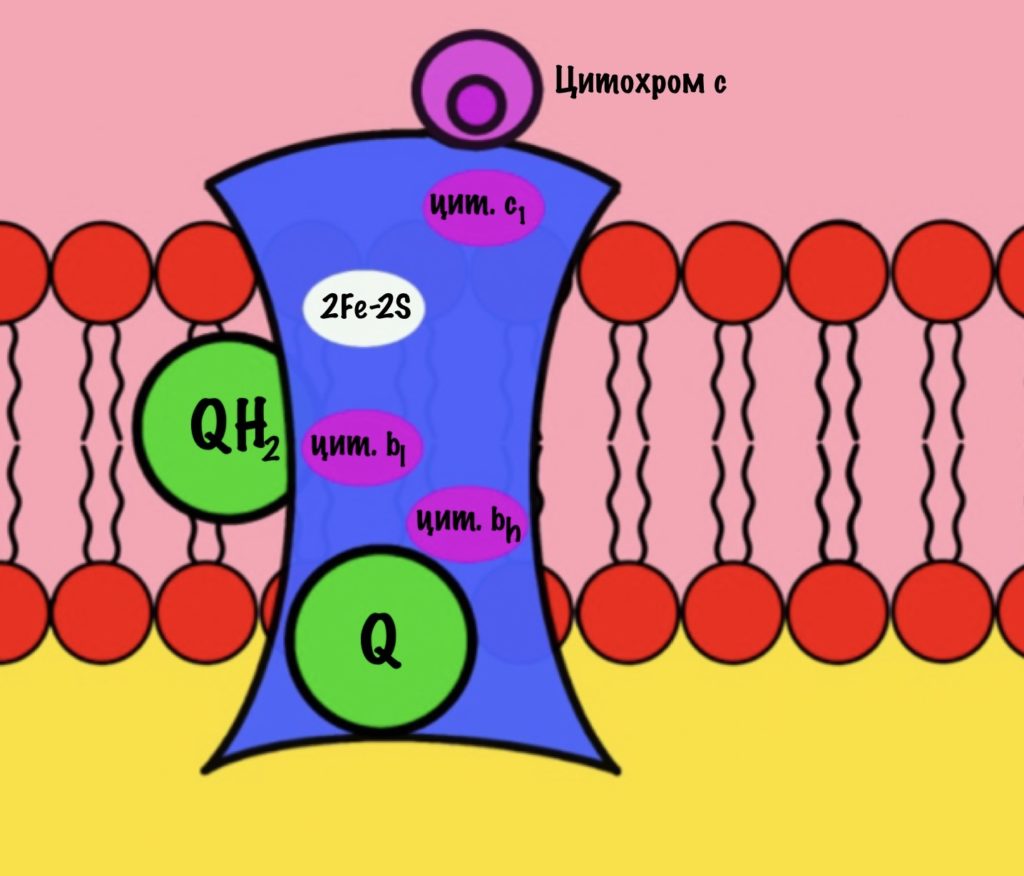
Задача этого комплекса — транспорт электронов с восстановленного убихинона на цитохром c. Разберём на примере одной субъединицы.
К комплексу присоединяется восстановленный убихинон. Он отдаёт один электрон на цитохром b, а другой на железо-серный белок. Дальше железо-серный белок отдаст электрон цитохрому c1. Почему только один? Вспоминаем, что железо может присоединить только один электрон — перейти в +2. Поэтому нужна такая развилка для электронов. В это же время от убихинона отщепляются два водорода, которые транспортируются в межмембранное пространство третьим комплексом. Убихинон становится окисленным и отсоединяется от третьего комплекса.
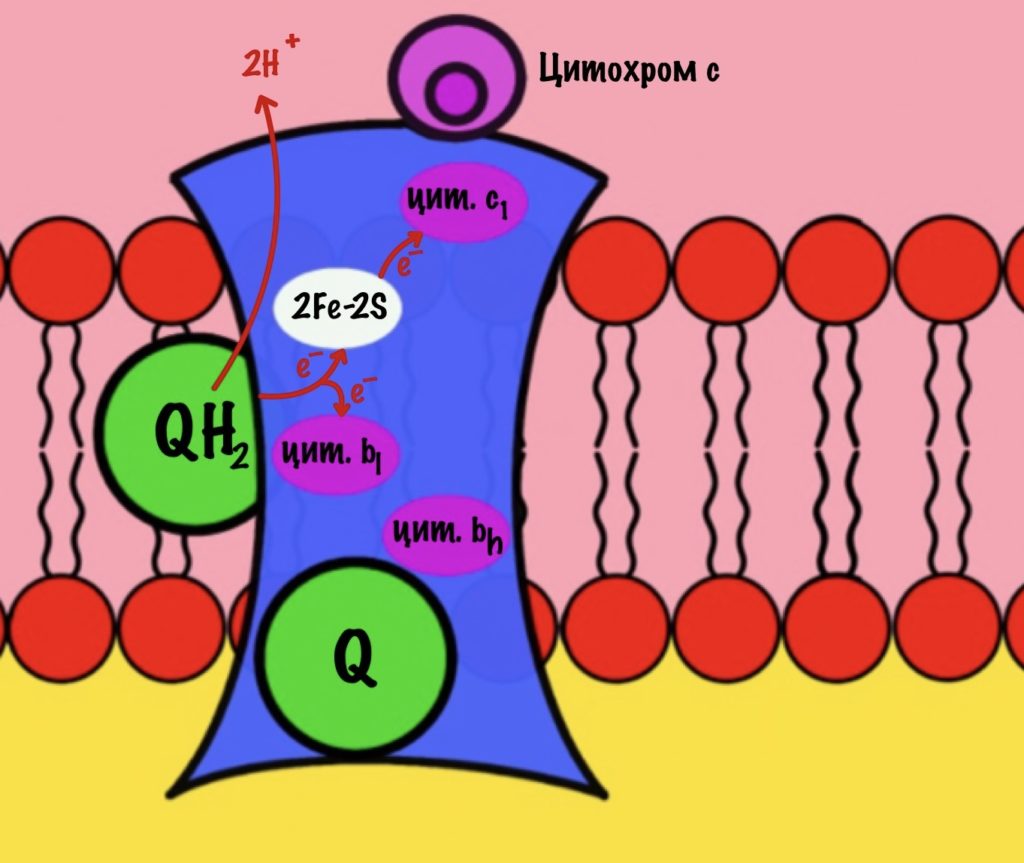
Цитохром c1 передаёт электрон цитохрому c. Тот говорит: «до связи». И отчаливает от третьего комплекса. Дальше восстановленный цитохром c попадёт на четвёртый комплекс, но об этом пока рано. К третьему комплексу присоединится следующий окисленный цитохром c.
Что будет со вторым электроном? У цитохрома b есть два порфирина с железом: l и h. Электрон переходит с порфирина l на h, а дальше падает на убихинон. Но не тот же самый, а другой — окисленный. Он сидел в третьем комплексе и ждал своего часа. Также убихинон присоединяет протон водорода из матрикса — образуется полувосстановленный убихинон (семихинон). С этой молекулой пока ничего не сделаешь.
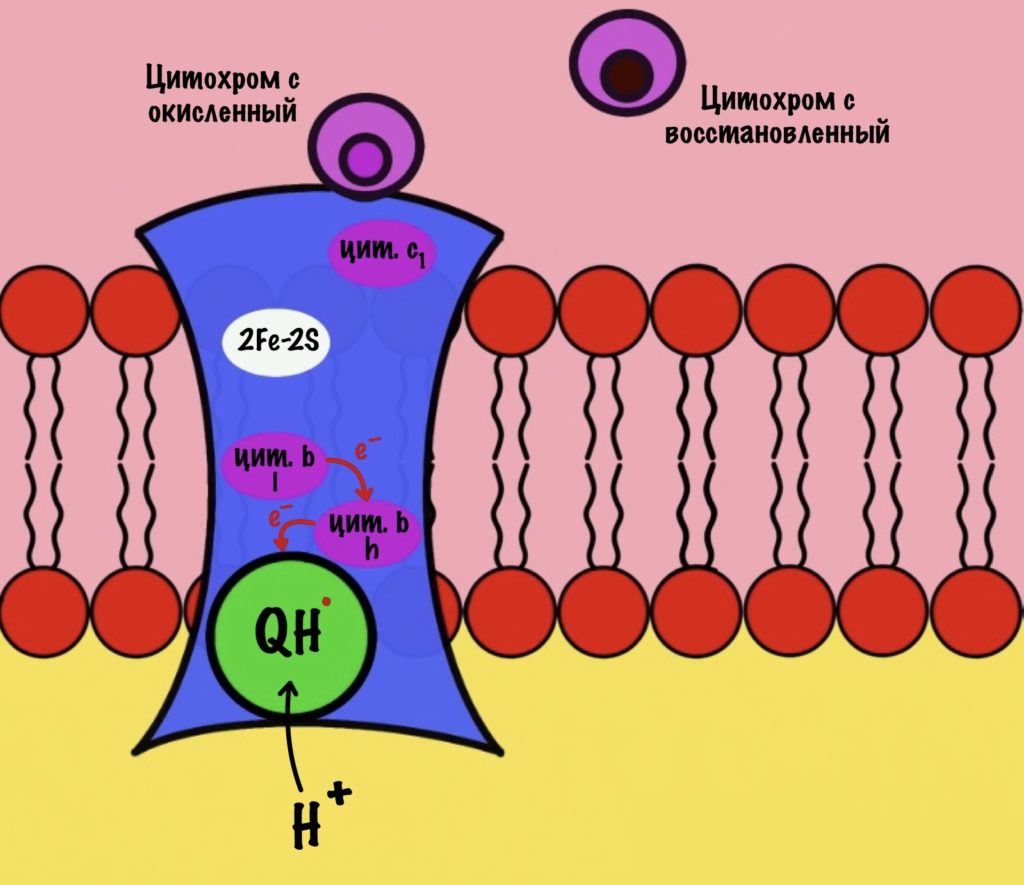
К третьему комплексу присоединяется ещё один восстановленный убихинон. Он снова отдаёт один электрон железо-серному белку, а другой цитохрому b. Два протона от убихинона вылетают в межмембранное пространство. От железо-серного белка электрон попадает на цитохром c1, а потом и на цитохром c. Ещё один электрон отправился на четвёртый комплекс.
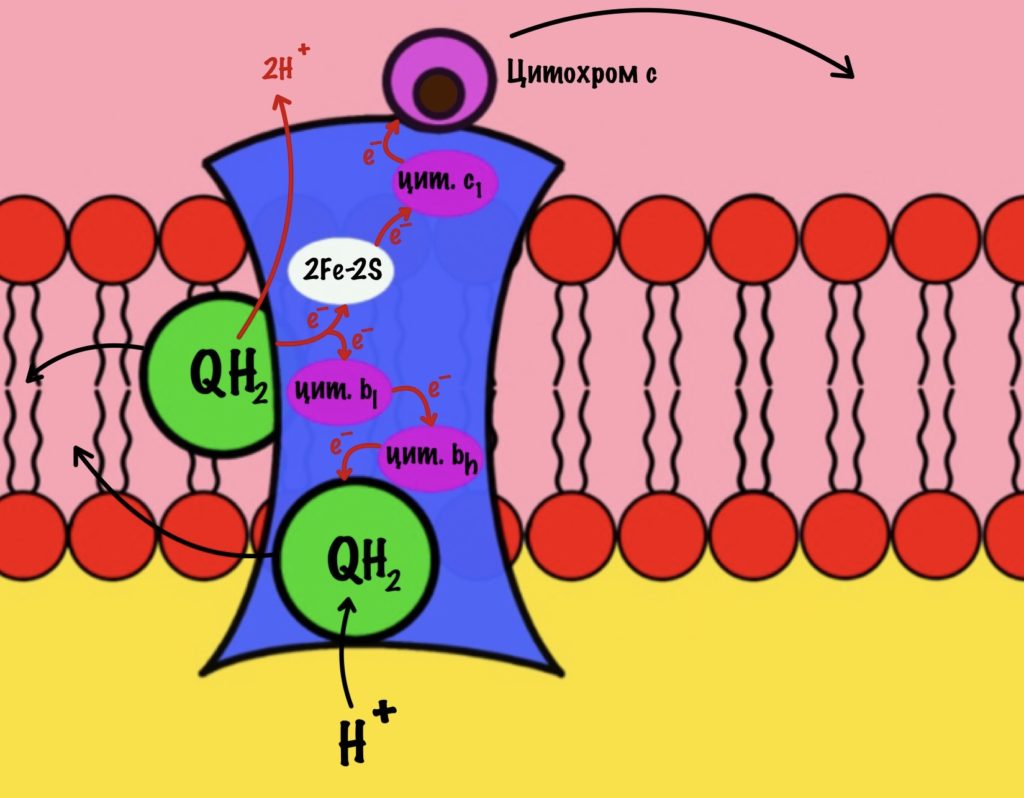
Второй электрон и протон из матрикса присоединяются к полувосстановленному убихинону, который ждёт своего часа. Он переходит в восстановленный убихинон. Восстановленный убихинон отсоединяется от третьего комплекса, чтобы присоединиться в другом месте к третьему комплексу))) Цикл завершён, у него даже есть своё название — цикл Q или убихиноновый цикл.
Третий комплекс переносит два электрона от убихинона на два цитохрома c. Энергия окислительно-восстановительных реакций используется на транспорт четырёх протонов в межмембранное пространство.
Четвёртый комплекс — цитохром c оксидаза
Цитохром с оксидаза имеет димерную структуру. Каждый мономер состоит из 13 субъединиц. Его задача — перенос электронов от цитохрома c на кислород. Кислород — это финальный акцептор электронов в дыхательной цепи.
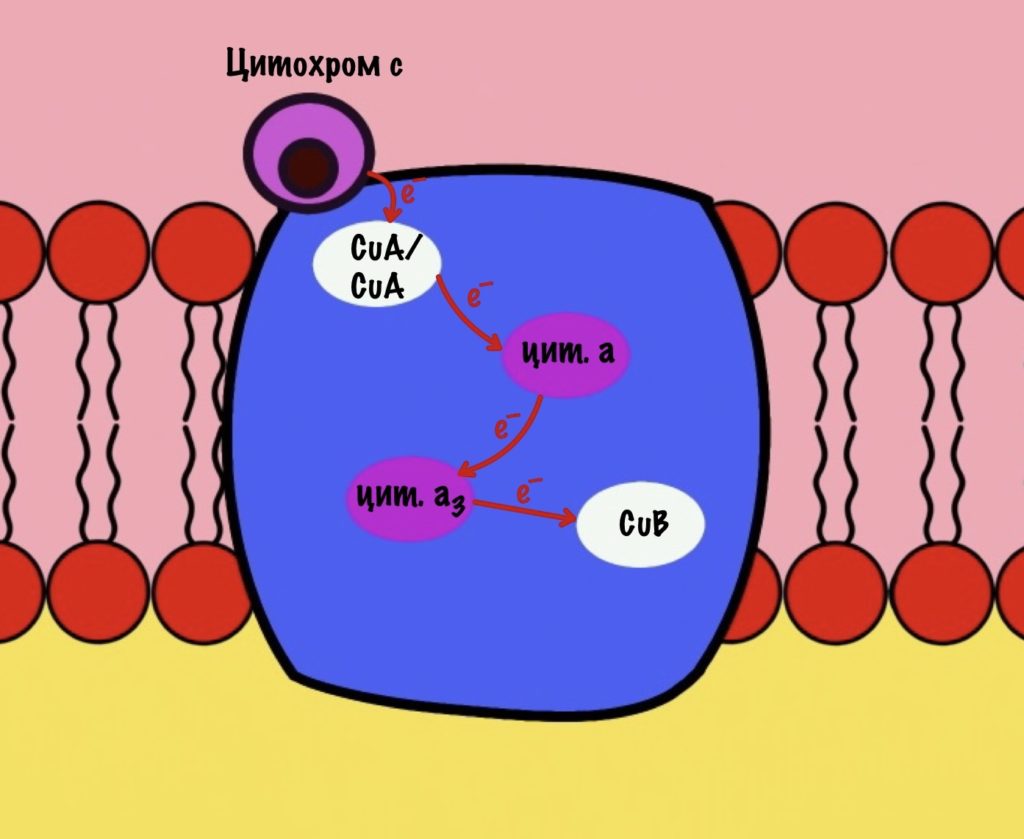
Второй мономер отвечает за связывание цитохрома c и присоединение его электрона. Электрон принимают на себя два атома меди, которые связаны ковалентной связью друг с другом. К четвёртому комплексу атомы меди присоединяются с помощью цистеина, гистидина и метионина — это напоминает железо-серные белки. За счёт такого строения эти два атома могут принять только один электрон. Я запишу эти ионы вот так — CuA/CuA.
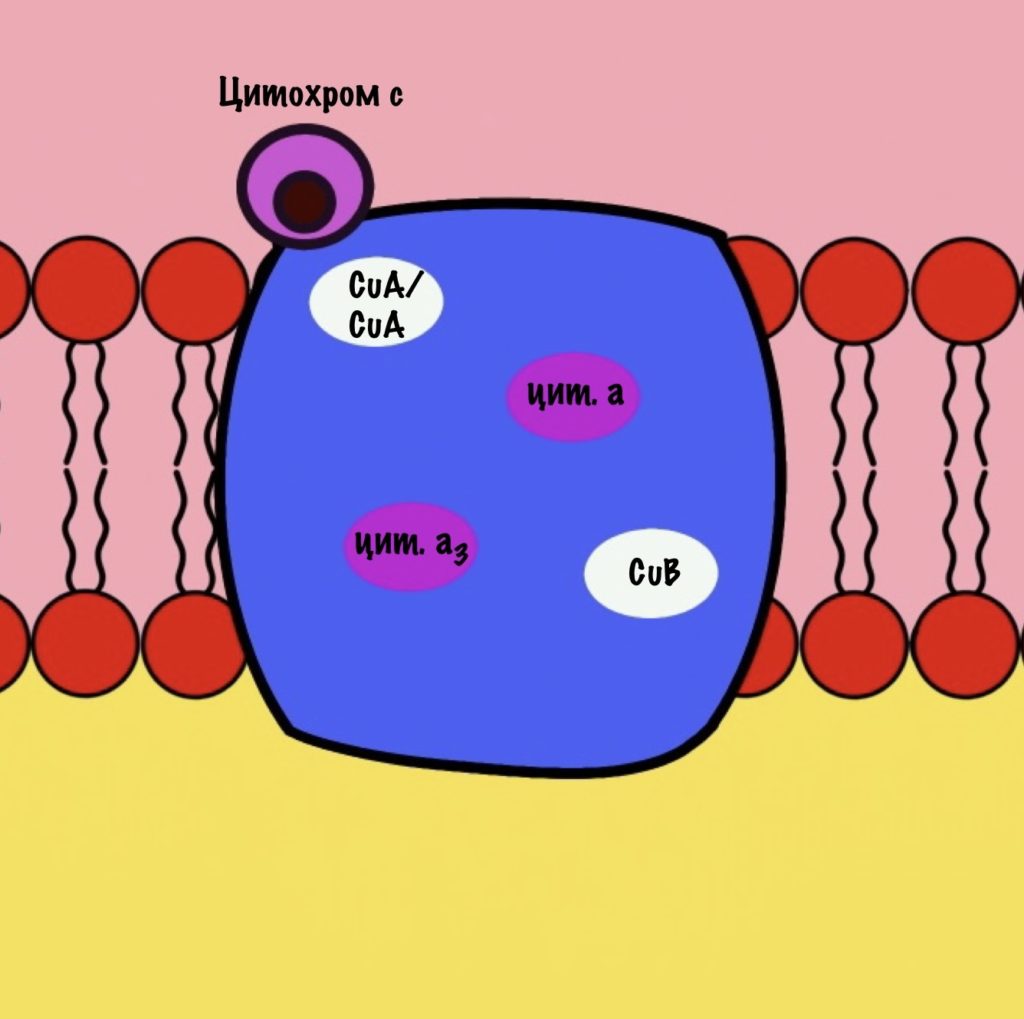
Первый мономер отвечает за передачу электронов на кислород. Он состоит из двух цитохромов: a и a3. Ещё в нём есть один ион меди +2 — CuB. Медь связана с цитохромом a3 (хоть на рисунке это и не видно).
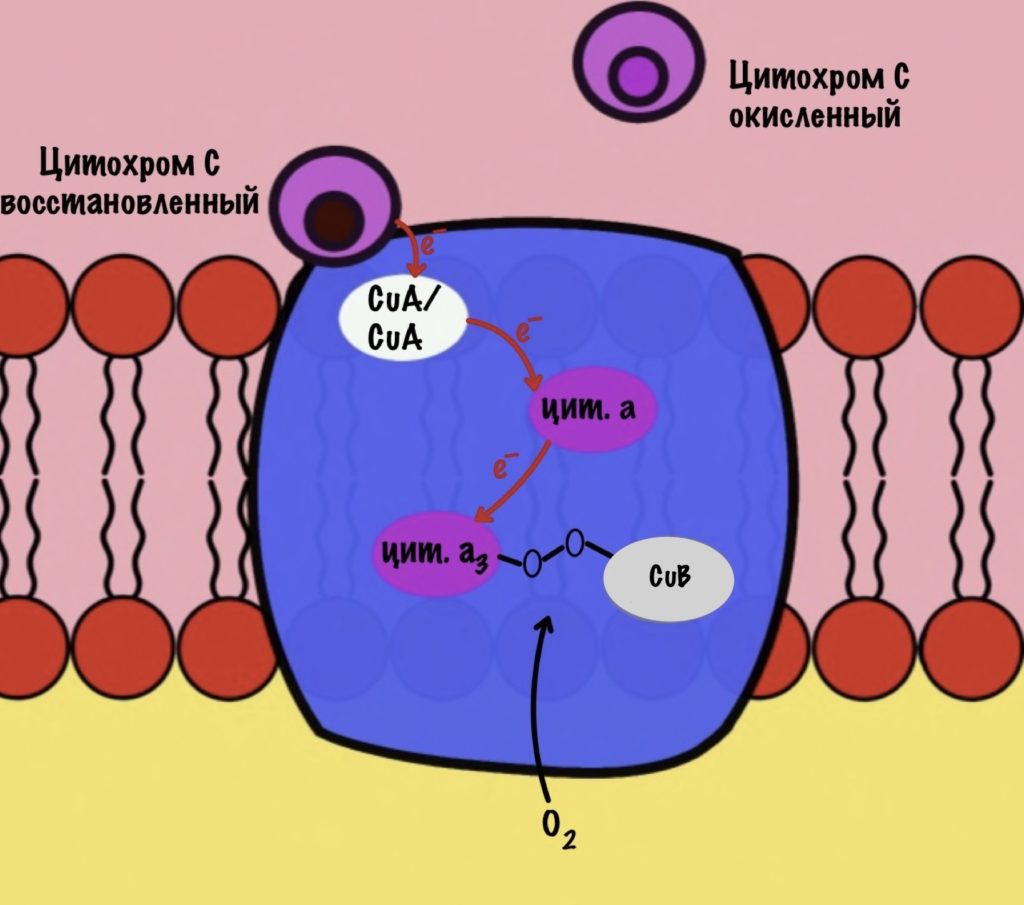
Цитохром c присоединяется к четвёртому комплексу и передаёт свой электрон на CuA/CuA. Далее атомы меди отдают электрон на цитохром a, а тот на цитохром a3. Они могут принять только по одному электрону — там одно железо +3. Цитохром a3 скидывает электрон на CuB. Медь меняет степень окисления — переходит из +2 в +1.
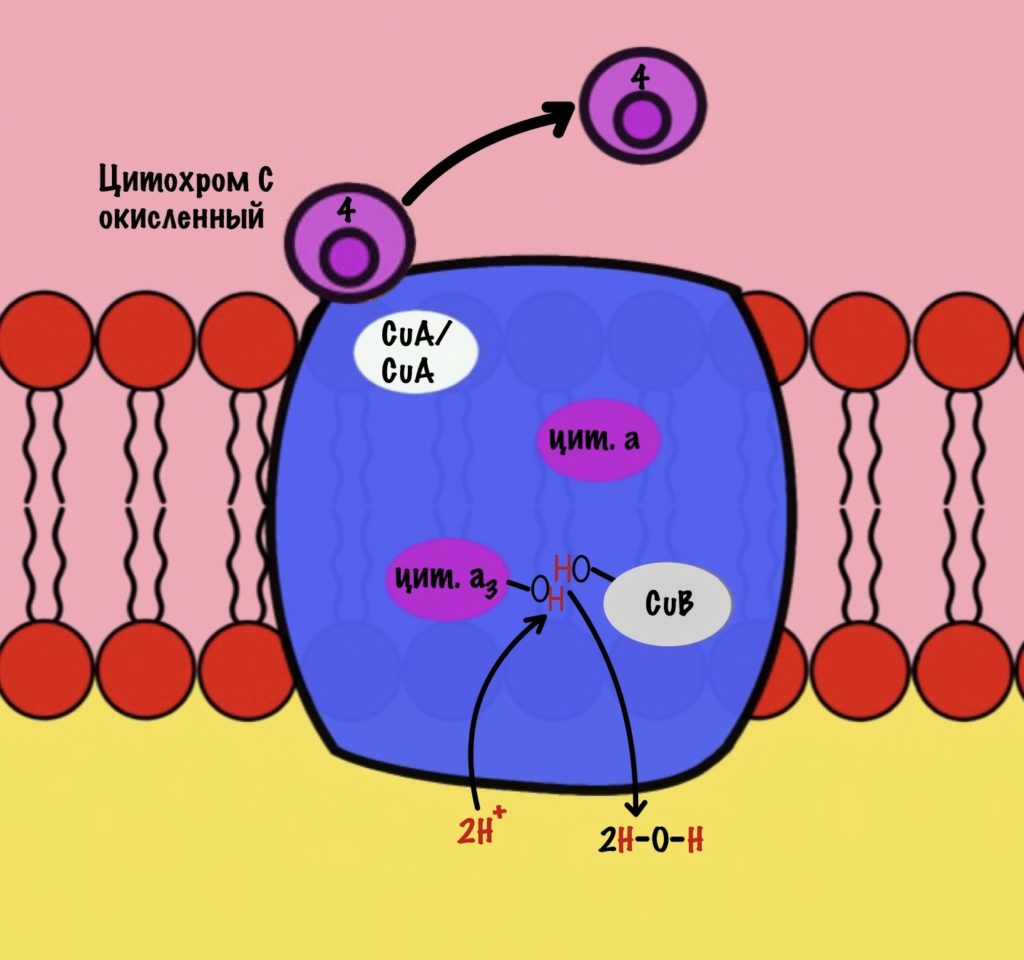
Дальше присоединяется второй цитохром c. Повторяется всё, что я написал выше. Но электрон не может попасть на CuB, потому что она уже +1, а больше электронов принять не может. Поэтому электрон останавливается на цитохроме a3. Теперь цитохром a3 и ион меди +1 могут связаться с кислородом — формируется пероксидный мостик. Кислород поглощает два электрона: один с цитохрома a3, а другой с CuB.
Присоединяется третий цитохром c. У меня ощущение, что я рассказываю анекдот с затянутой концовкой. Этот электрон используется для того, чтобы разрушить пероксидный мостик между железом и медью. Но нужен не только электрон, но и протон — он берется из матрикса митохондрии. Это приводит к разрушению пероксидного мостика. Остаётся цитохром а3, который связан с кислородом, и медь с гидроксильной группой.
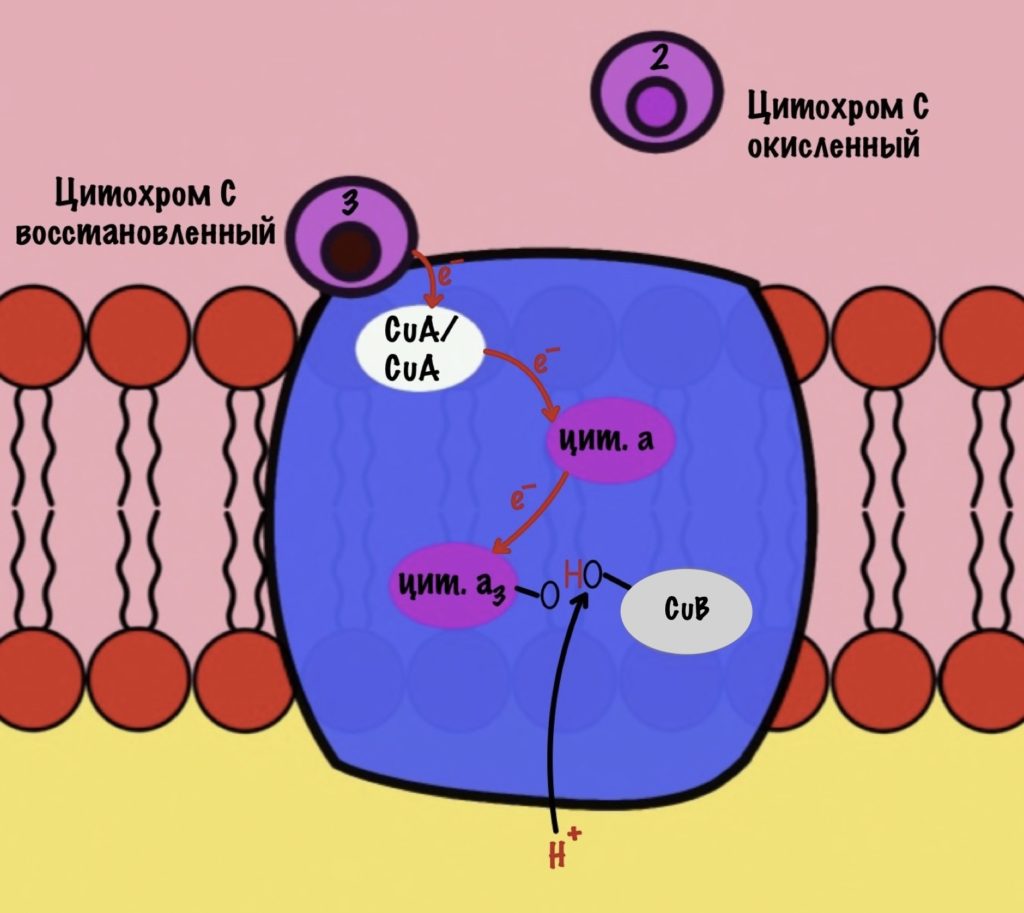
Присоединяется четвёртый электрон от цитохрома с, а один протон берется из матрикса. Это приводит к образованию гидроксильной группы на цитохроме а3.
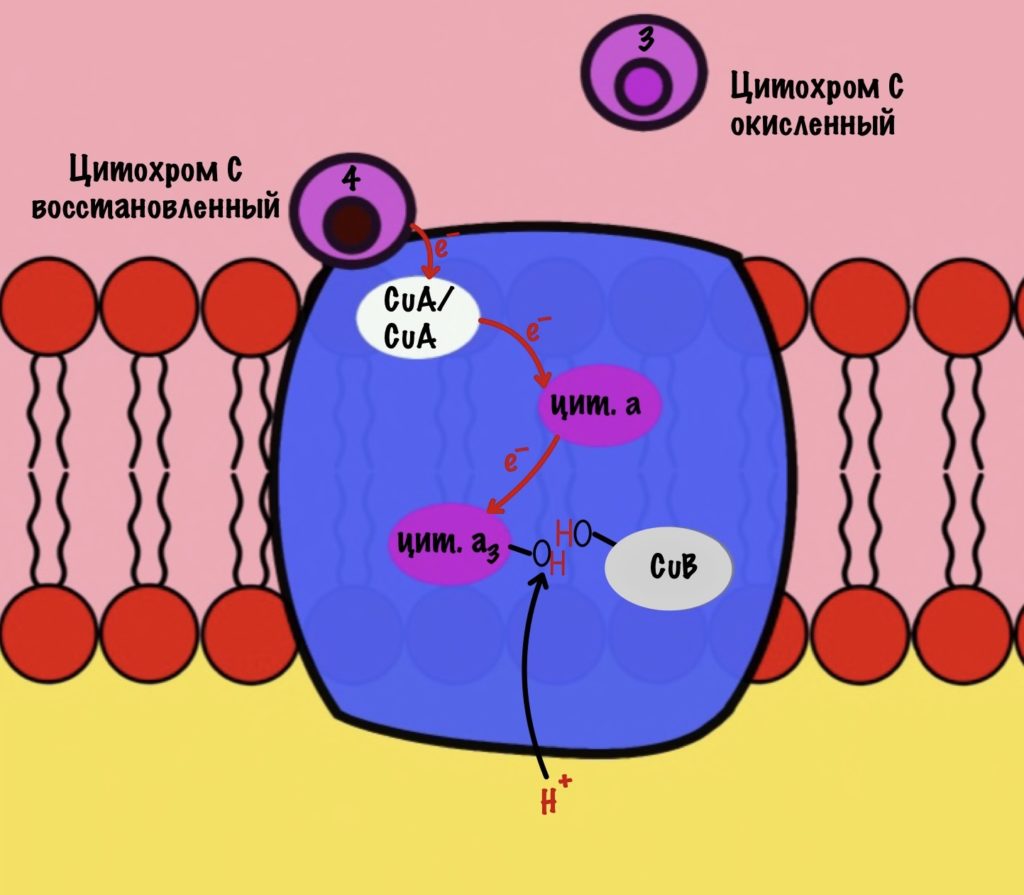
Из матрикса берутся ещё два протона — это приводит к образованию двух молекул воды. При этом цитохром а3 и медь возвращаются в свою начальную форму.
Обсудим, что у нас вышло. Мы присоединили четыре электрона к четвёртому комплексу, а также взяли из матрикса четыре протона водорода и кислород. В итоге мы получили две молекулы воды, да ещё избавились от опасных электронов. Они могут много чего натворить в клетке. Энергия окислительно-восстановительных реакций одного электрона используется на транспорт одного протона из матрикса в межмембранное пространство. Всего использовали четыре электрона — четыре протона вылетело в межмембранное пространство.
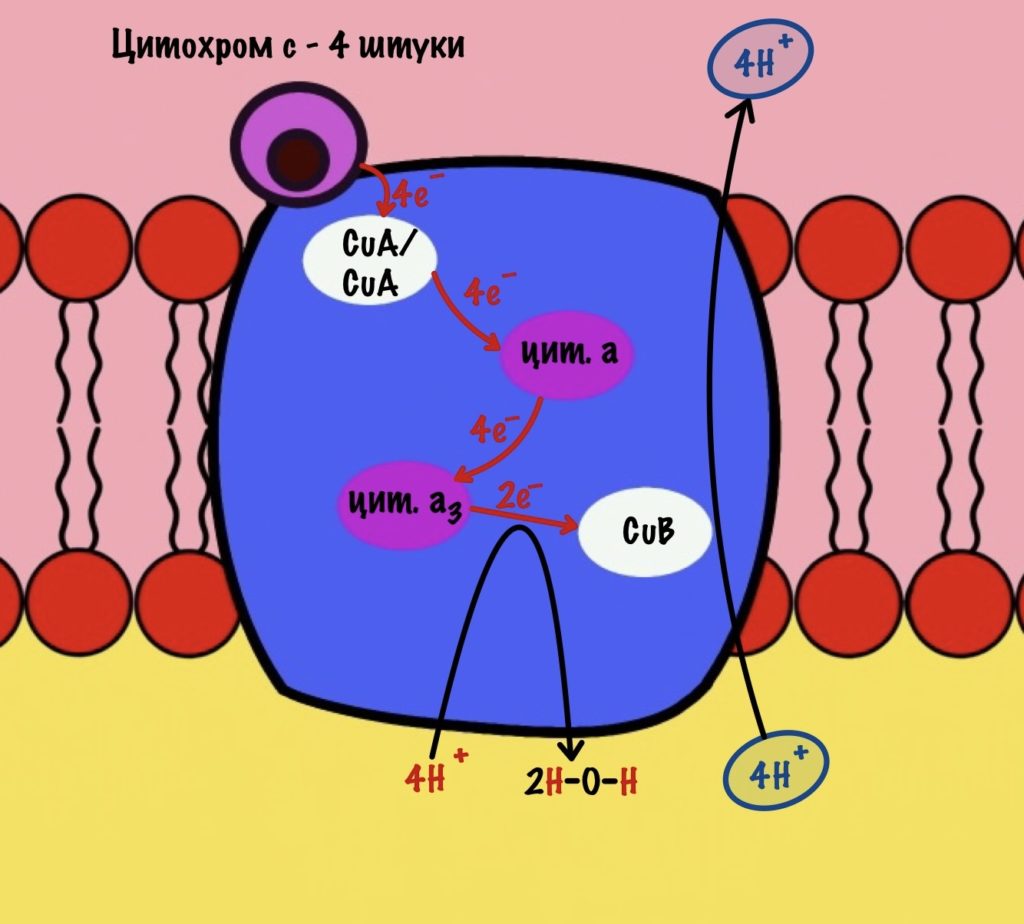
Воспоминаем, что до этого мы использовали только по два электрона во всех комплексах. Да и НАДH и ФАДH2 несли себе всего по два электрона. Не проблема, просто поделим всё на два. И у нас получится:
Четвёртый комплекс переносит два электрона на 1/2 кислорода — образуется молекула воды. Энергия восстановительных реакций используется для транспорта двух протонов водорода.
Общий подсчёт: сколько протонов транспортируется из матрикса за счёт двух электронов?
В ходе работы первого комплекса были транспортированы 4 протона. Третьего — 4 протона. А четвёртого — 2 протона. Второй комплекс протоны не транспортирует. Первый комплекс присоединяет НАДH, а второй — ФАДH2. Второй комплекс передаёт электроны на третий комплекс, пропуская первый. Получается, что энергия электронов одной молекулы НАДH используется на транспорт 10 протонов. А энергия электронов одной молекулы ФАДH2 на транспорт 6 протонов.
Энергия электронов 1 НАДH идёт на транспорт 10 протонов.
Энергия электронов 1 ФАДH2 идёт на транспорт 6 протонов.
Теперь нам нужно превратить эти протоны в энергию.
АТФ-синтаза
АТФ-синтаза — это фермент, который состоит из двух больших частей (или доменов): Fо и F1. Fо — это протонная помпа, которая встроена в мембрану. Если перевести название на русский — олигомицин-чувствительная фракция (часть). F1 — это каталитический домен, который синтезирует АТФ. F1 — первая фракция (часть).
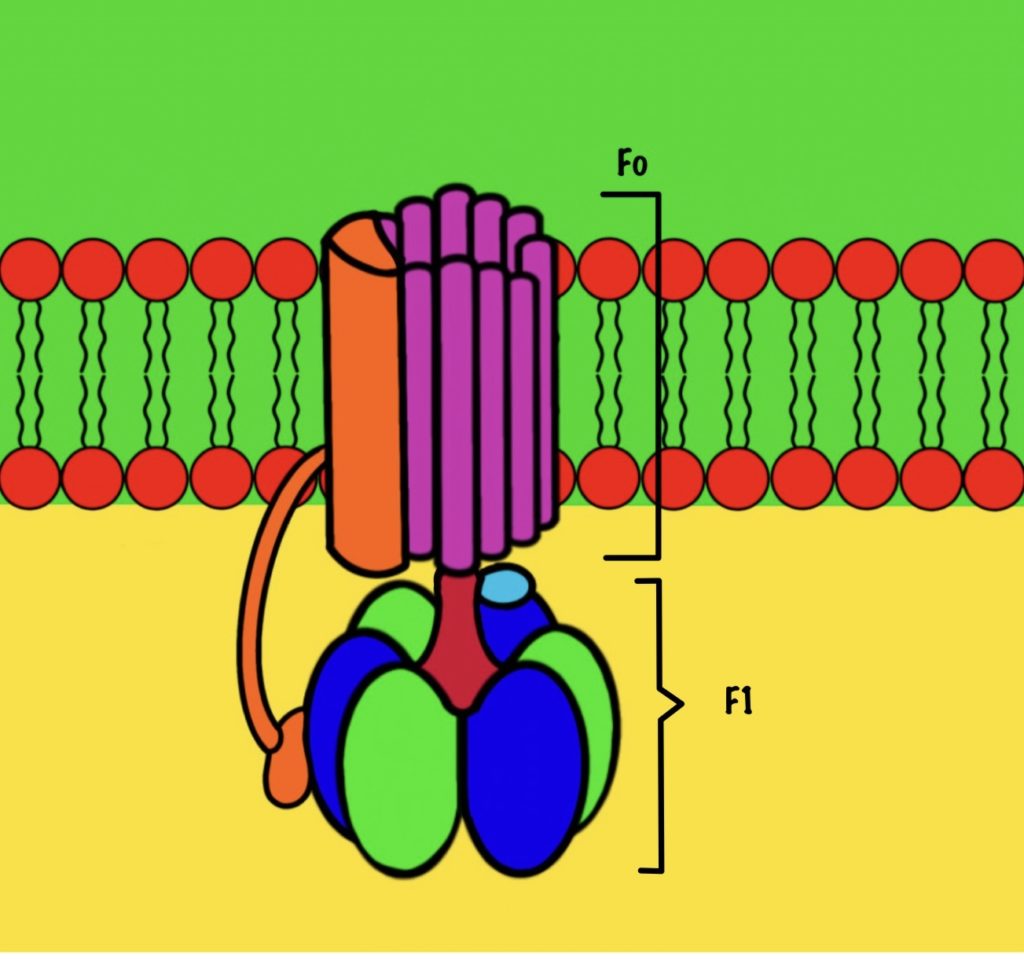
F0 домен состоит из 10-14 субъединиц c и одной субъединицы a. C субъединицы образуют жёсткое кольцо — оно вращается, когда присоединяет протон водорода. В субъединице a есть полуканалы для протонов. Именно полуканалы, а не целые. Один полуканал связан с межмембранным пространством, а второй с матриксом. Они отделены друг от друга.
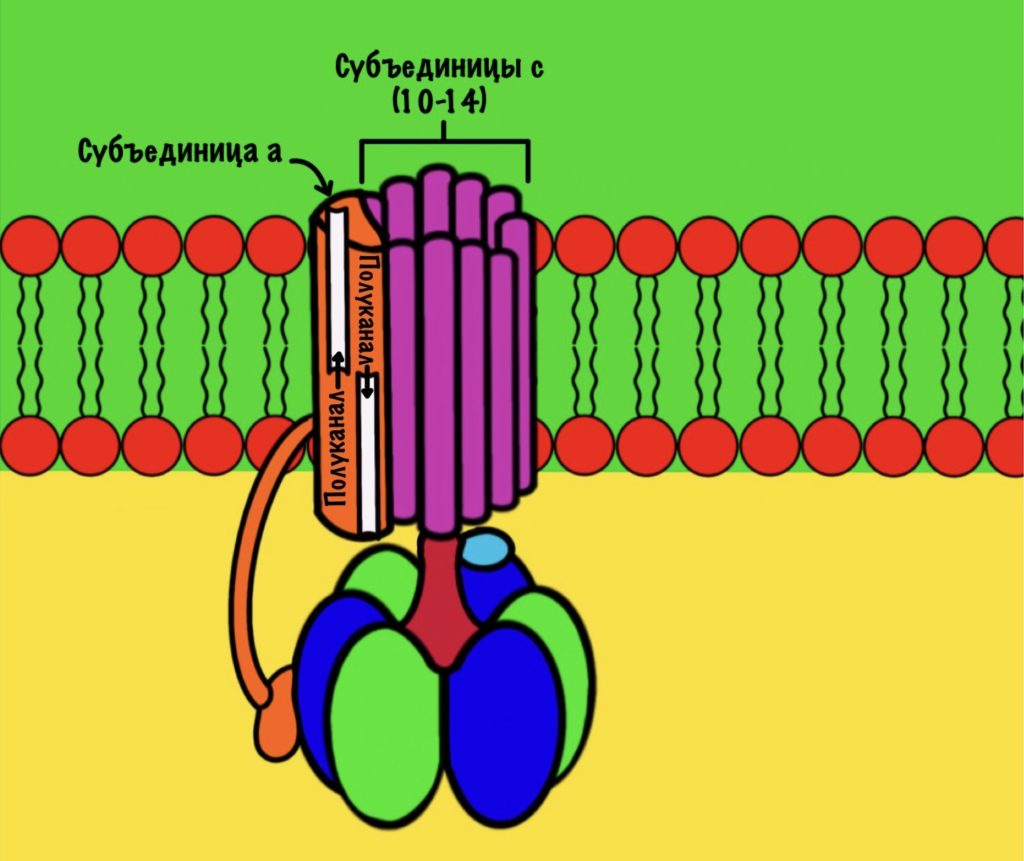
F1 домен состоит из пяти типов полипептидных цепей — альфа, бета, гамма, эпсилон и дельта. Три альфы и три беты образуют гексамер — похож на клевер. Он ответственен за синтез АТФ. Этот гексамер связан с 10-14 субъединицами с с помощью гамма и эпсилон цепей. Две эти цепи образуют ножку, которая вращается вместе с субъединицами c. Наибольшая роль здесь у гамма субъединицы. Вы можете подумать, что эта ножка будет вращать гексамер, но это не так! Гексамер всегда остаётся на месте, а вертится только ножка и субъединицы c.
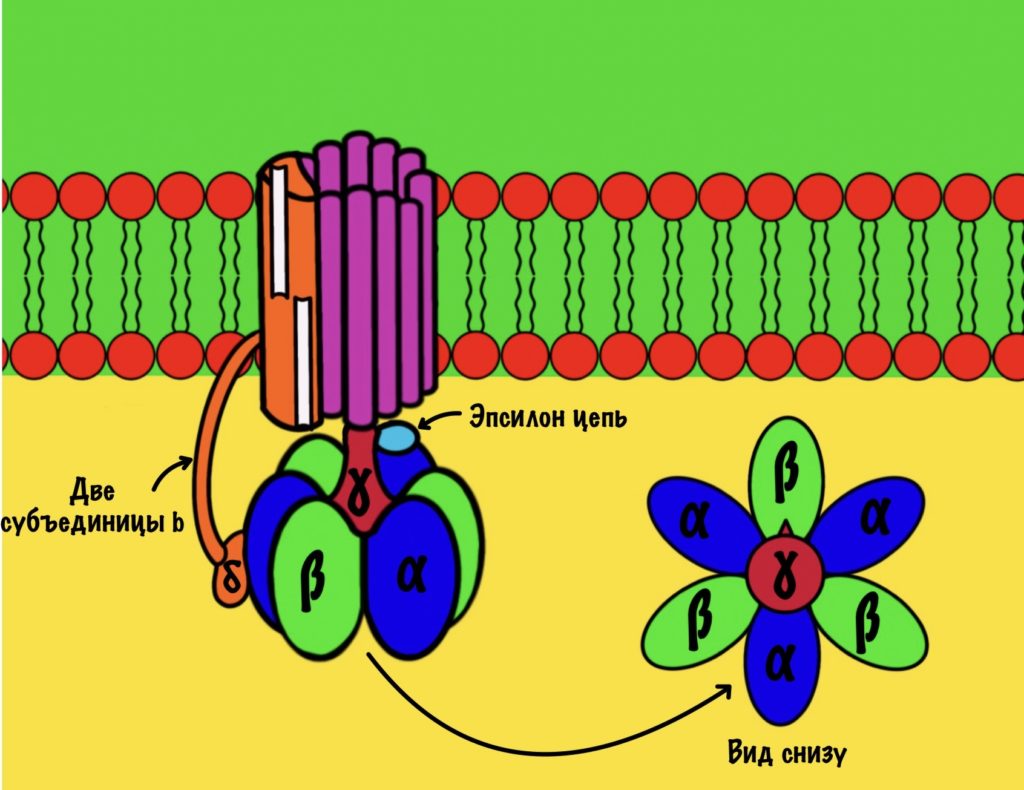
Дельта субъединица стабилизирует гексамер, она не даёт ему развалиться и вращаться вместе с ножкой. Ещё есть две субъединицы b, которые связывают между собой субъединицу a и субъединицу дельта.
Fо и F1 регион связаны через:
- Гамма ножку и эпсилон полипептидную цепь;
- Субъединицы b, которые связывают субъединицы a и дельта.
АТФ синтаза состоит из двух регионов:
- Вращательного — кольцо из субъединиц c и ножки гамма-эпсилон;
- Стационарного: гексамер из 3-альфа и 3-бета цепей, субъединицы a и b, а также дельта-субъединицы.
Регион F1
Начнём с него, а потом перейдём к вращению нашего кольца. Нас интересует гексамер, который участвует в синтезе АТФ. Гексамер состоит из трёх альфа и трёх бета цепей.

При этом альфа цепи всегда связаны с АТФ и не участвуют в его синтезе, поэтому они нас не интересуют. А вот бета цепи ответственны за:
- Связывание АДФ и неорганического фосфата;
- Синтез АТФ;
- Высвобождение АТФ.
Только один из этих процессов связан с вращением c-кольца, угадаете какой? Нет — это не синтез АТФ. Удивительно, но каждая из бета-цепей легко синтезирует АТФ. Они настолько сближают АДФ с фосфатом, а также создают гидрофобное окружение вокруг них, что реакция синтеза АТФ идёт без каких-либо проблем и затрат энергии. А вот высвободить образованную АТФ эти субъединицы не могут, они имеют слишком большое сродство к ней. Поэтому, если мы сравним обычную ферментативную реакцию с получением АТФ, то увидим такое…
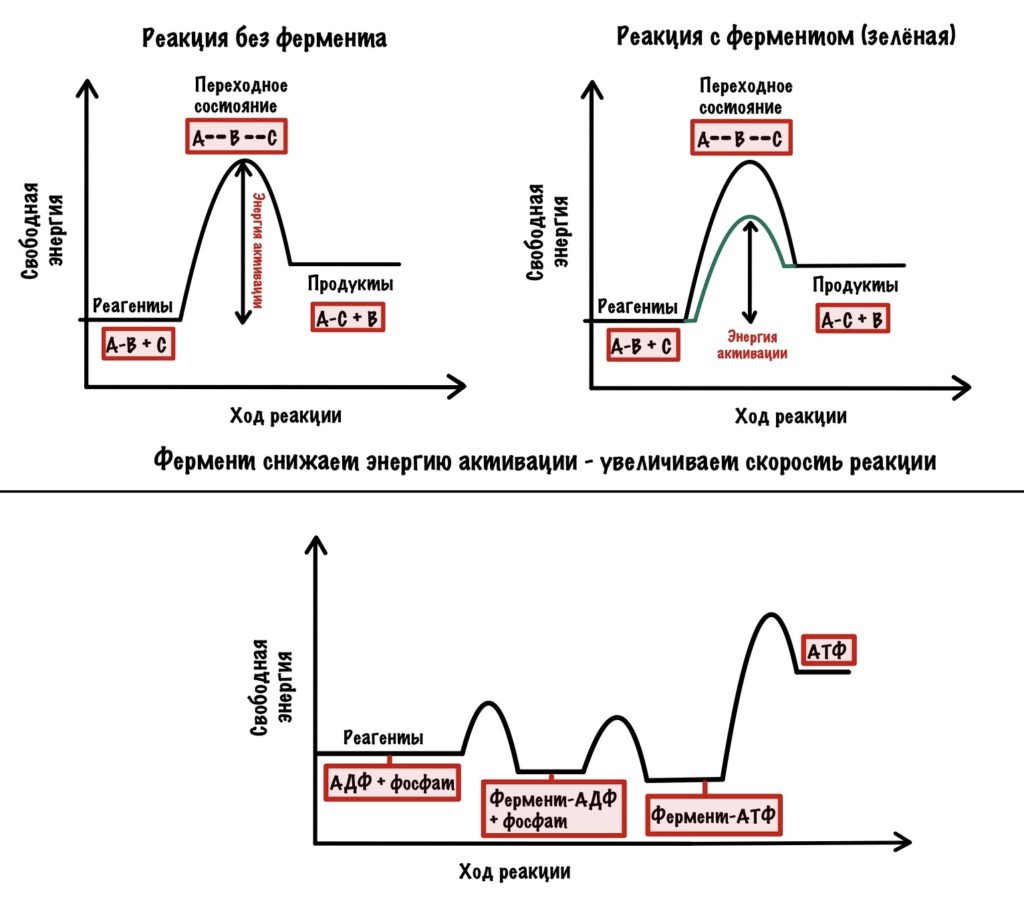
Видите? В образовании АТФ из АДФ и фосфата почти нет изменения свободной энергии, в отличие от ферментативной реакции. Но сколько нужно энергии для её высвобождения — жуть! Для того, чтобы высвободить АТФ нужно вращение, создаваемое c-кольцом. Вращение предоставляет энергию для высвобождения АТФ. Оно вызывает конформационные изменения в бета-субъединицах, благодаря которому они выпускают АТФ из своих лап.
Вернёмся к бета-цепям. Они могут быть в трёх состояниях: open, loose и tense (открытое, свободное и напряжённое). Или O, L и T состояния. В O-состоянии бета-субъединица присоединяет АДФ и неорганический фосфат. В L-состоянии бета закрывает фосфат и АДФ в ловушку — они не могут выбраться из субъединицы. В T-состоянии синтезируется АТФ, которая не может отсоединиться от фермента из-за его высокого сродства.
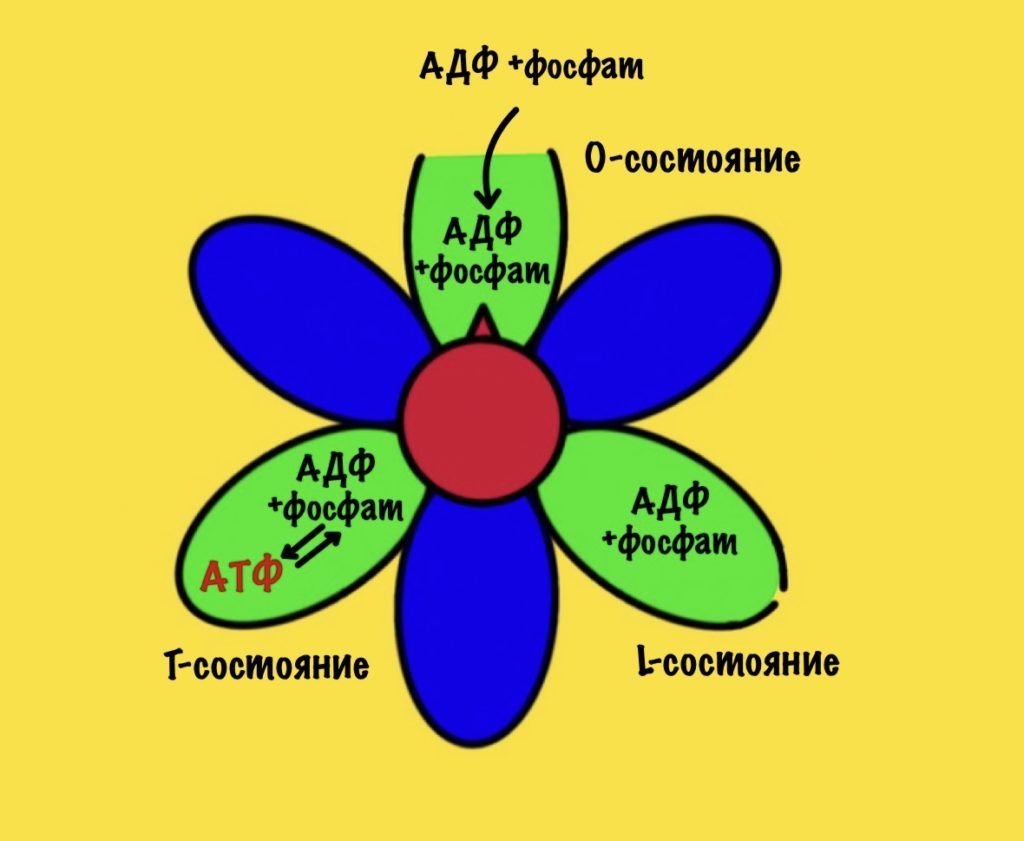
В АТФ-синтазе каждая из бета-субъединиц находится в одном из этих состояний. Одна в O, другая в L, а третья в T. Вращение гамма ножки вызывает переход бета-субъединицы из одного состояния в другое. Получается, что любые две бета-субъединицы не могут существовать в одном состоянии одновременно.
Ножка вращается на 120 градусов по часовой стрелке (если смотреть снизу). Это вызывает конформационные изменения в субъединицах, что приводит к переходу из одного состояния в другое. При этом одна бета-субъединица переходит из O-состояния в L. Вторая из L-состояния в T. А третья из T-состояния в O. В О состоянии бета-субъединица отсоединяет АТФ, а потом связывается со следующим АДФ и неорганическим фосфатом.
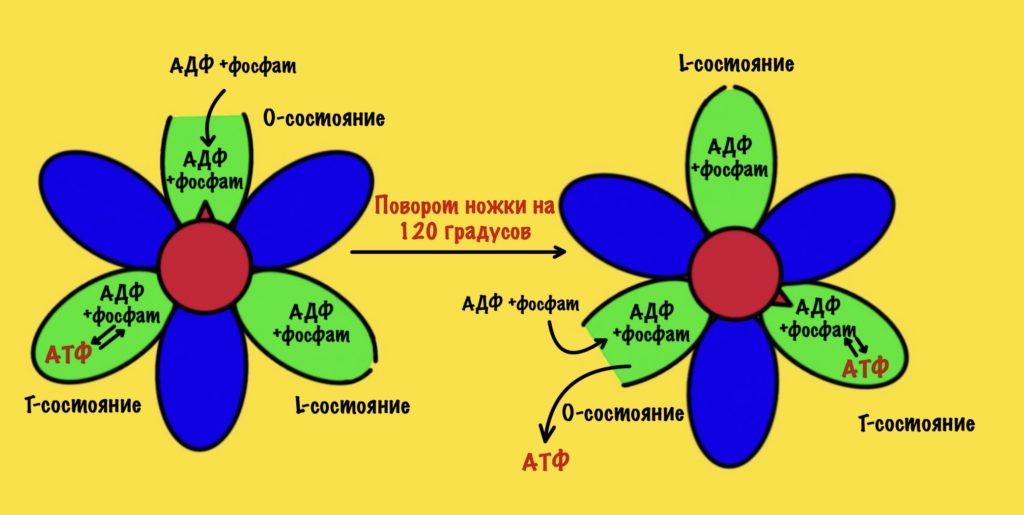
Давайте сделаем полный круг вращения. 360 градусов — это три поворота для нашей ножки. Каждая субъединица изменит свое состояние три раза. Получается, что в ходе такого вращения будет синтезировано три молекулы АТФ.
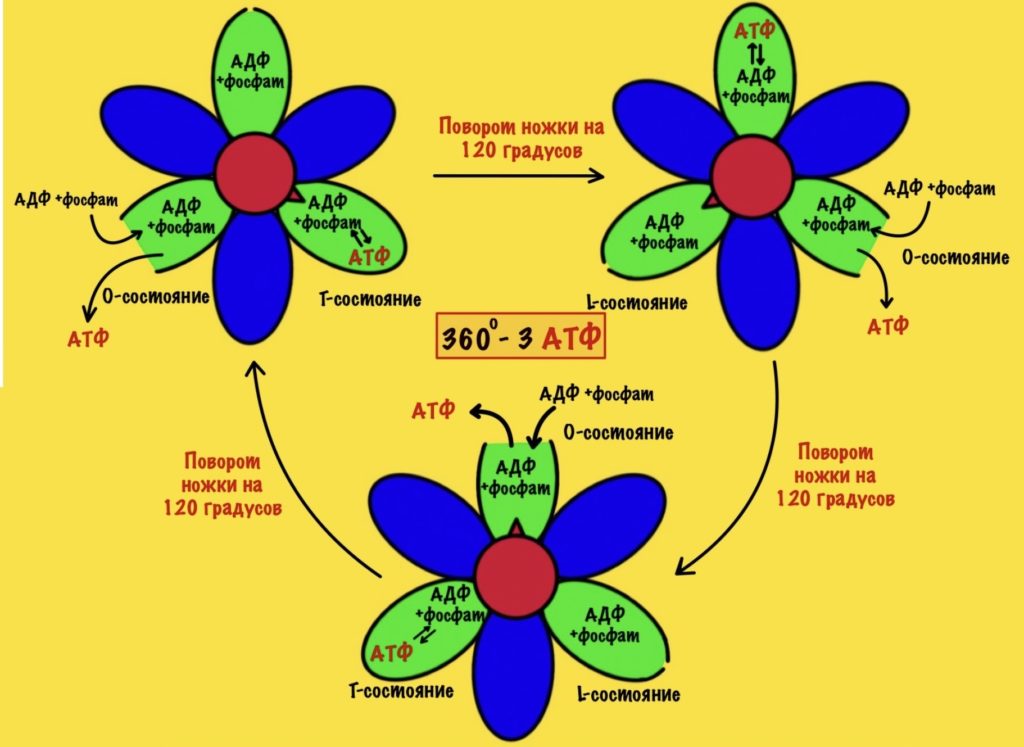
Регион Fo
Теперь нужно понять механизм вращения ножки. Субъединица a связана с кольцом из 10-14 субъединиц c. Напоминаю, что в а-субъединице есть полуканалы для протонов.
Если посмотреть на этот полуканал, то мы увидим кое-что интересное. В конце полуканала находится аргинин от субъединицы a. Это положительно-заряженная аминокислота. Аргинин взаимодействует с аспартатом из субъединицы c. Аспартат — это отрицательно-заряженная аминокислота. Аспартат и аргинин взаимодействуют, как разноимённо-заряженные частицы. Немного разверну рисунок для удобства.
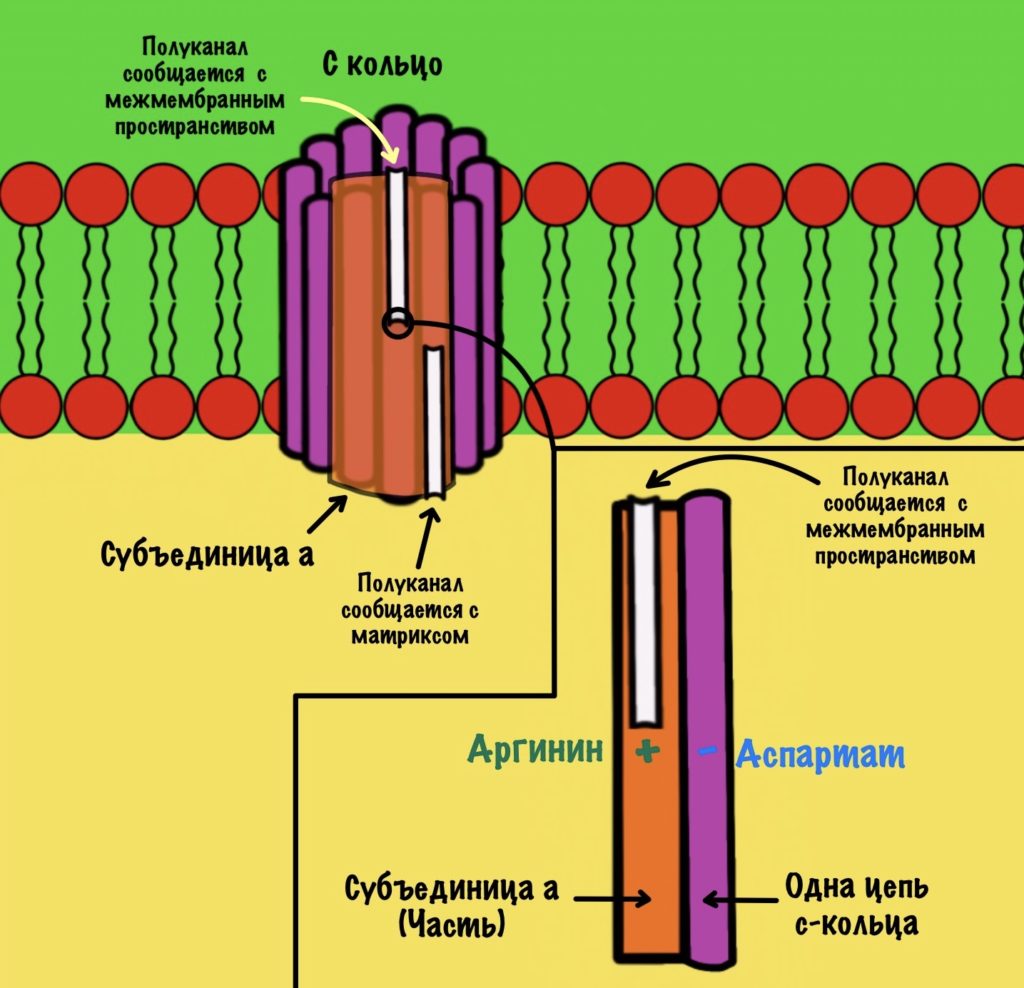
Протон пролетает через полуканал а субъединицы и присоединяется к аспартату. Из-за того, что у аспартата отрицательный заряд, а у протона положительный. Аспартат и аргинин перестают взаимодействовать друг с другом, так как аспартат потерял свой отрицательный заряд.
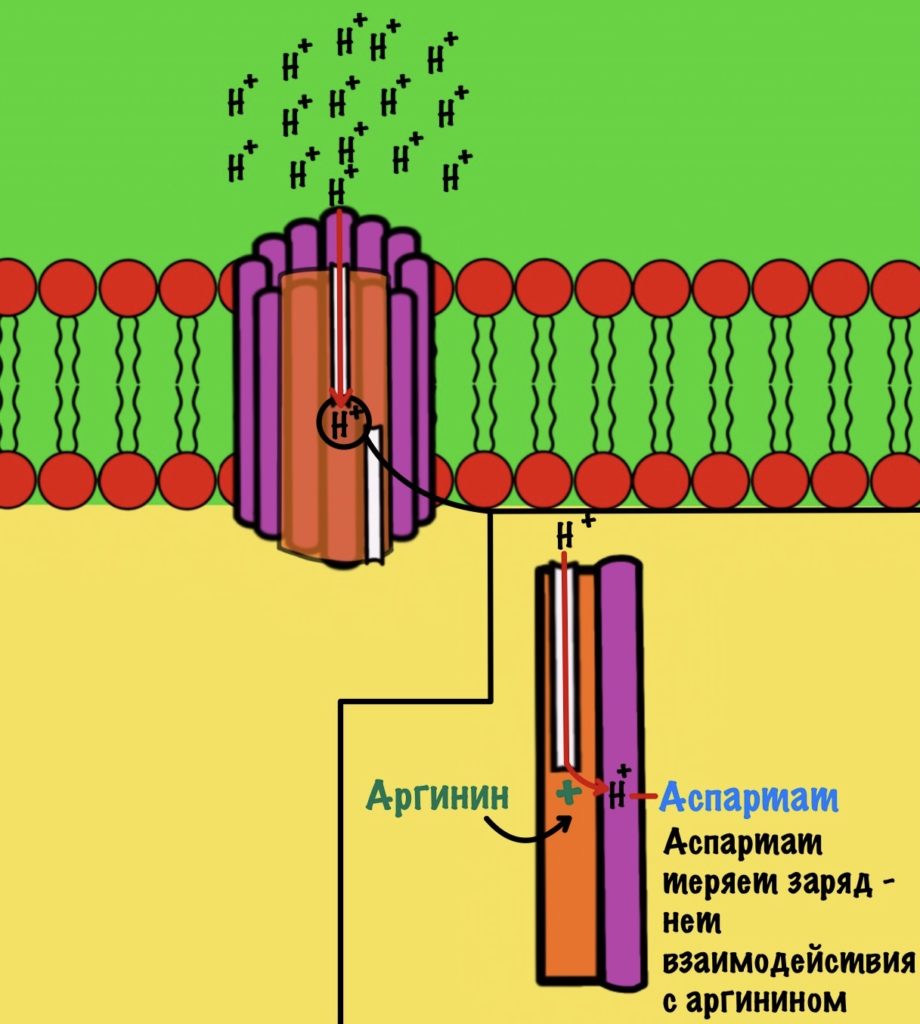
Это приводит к вращению всего кольца субъединиц c по часовой стрелке — на одну субъединицу c. При этом вращается и ножка гамма-эпсилон, что приводит к конформационным изменениям в гексамере. Ну это тот,который синтезирует АТФ.
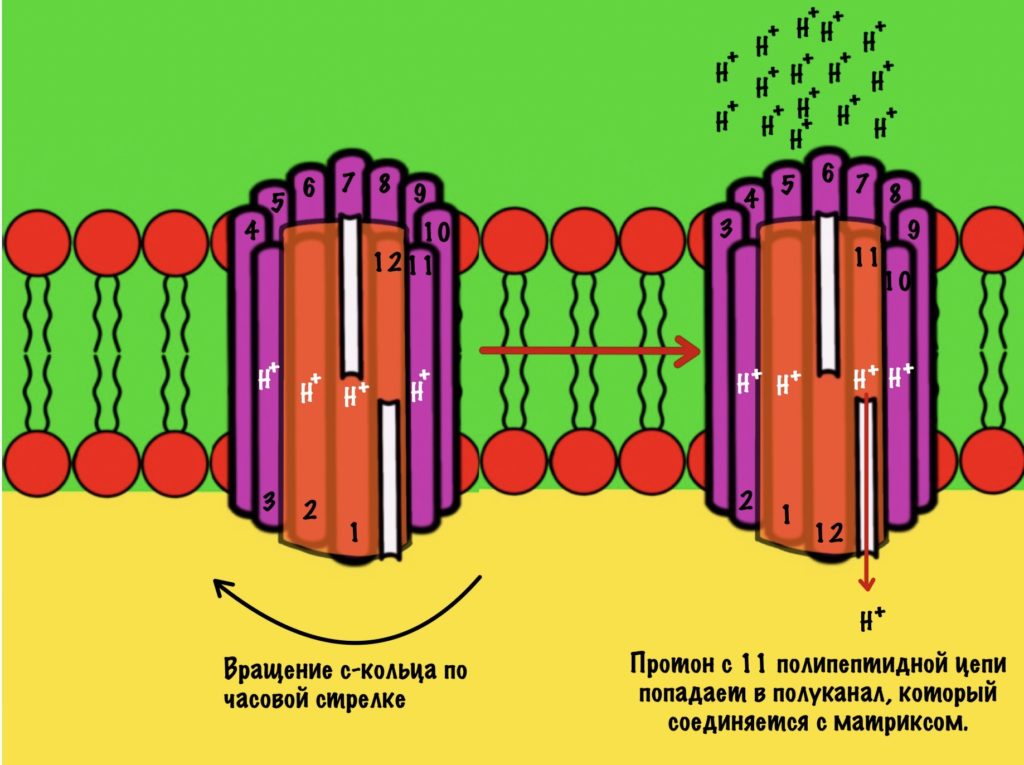
11 субъединица взаимодействует с каналом, который соединяется с матриксом. Аспартат этой субъединицы отдаёт протон водорода — он попадает в матрикс. Дальше 12 субъединица присоединит протон из межмембранного пространства, что приведёт к следующему вращению кольца. Теперь 11 субъединица присоединит протон и так далее… Удивительно, но природа придумала двигатель так давно!
Если мы хотим провести полное вращение c кольца — 360 градусов, то нам нужно присоединить к каждой субъединице с по одному протону. Так и происходит. Помните, что в кольце от 10 до 14 c субъединиц? Значит на полное вращение нам понадобится 10-14 протонов. В моём вот было 12 полипептидных цепей, значит для полного круга нужно присоединить 12 протонов.
При этом произойдёт полное вращение ножки — три раза по 120 градусов. Значит каждый гексамер изменит своё состояние три раза — три поворота по 120 градусов. Получается, что одно полное вращение с кольца, а с ним и ножки, приводит к синтезу трёх молекул АТФ.
Сколько протонов нужно для синтеза одной молекулы АТФ?
10-14 протонов на одно вращение на 360 градусов = 3 молекулы АТФ
Получается, что для синтеза одной молекулы АТФ нужно от 3,33 до 4,66 протонов. Или в среднем — 4 протона.
Для синтеза одной молекулы АТФ нужно 4 протона.
При транспорте электронов с НАДH было получено 10 протонов = 2,5 молекулы АТФ.
При транспорте электронов с ФАДH2 было получено 6 протонов = 1,5 молекулы АТФ.
Так что теперь вы не поведётесь на то, что пишут в некоторых учебниках: «Из одного НАДH синтезируется 3 молекулы АТФ, а из одного ФАДH2 — 2 молекулы АТФ. Поздравляю!
Нарушение работы окислительного фосфорилирования
Ух, ну и системку мы разобрали. Согласитесь, что она очень сложная. Но при этом отлично справляется со своей работой — в норме. Но давайте немного отойдём от нормы. Время поиграть в доктора!
Представьте себе, что у пациента анемия — снизились эритроциты и/или гемоглобин. Меньше кислорода поступает в ткани и органы, а значит в митохондриях дела плохи. Электроны не могут попасть на свой финальный акцептор — кислород. Электроны застревают в комплексах дыхательной цепи. Они не создают градиент концентрации, поэтому нет синтеза АТФ. Пациент с анемией чувствует общую слабость именно из-за этого.
В организме истощился запас железа. Тут два патологических механизма. Дефицит железа приводит к снижению гемоглобина, а дальше вы уже знаете. Железо идёт не только на синтез гемоглобина, но и цитохромов — компонентов третьего и четвёртого комплекса дыхательной цепи. Если нет акцепторов электронов, то нет и окислительного фосфорилирования.
Дефицит меди. Медь входит в четвёртый комплекс дыхательной цепи, поэтому если её не будет, то комплекс перестанет работать.
Дефицит белка в рационе или нарушение его всасывания. Причин множество, но нас они не интересуют. Белок используется для синтеза собственных белков организма — ферментов и комплексов дыхательной цепи.
Гиповитаминозы. Коснёмся только двух, хотя почти каждый гиповитаминоз будет приводить к нарушениям. НАД+ и ФАД — это производные витаминов. НАД+ — никотиновой кислоты (витамин B3). ФАД — рибофлавина (витамин B2). Если словить два этих гиповитаминоза, то у нас просто не будет начальных акцепторов электронов. Так что электрончики не смогут запрыгнуть на дыхательную цепь с НАДH и ФАДH2.
Дефицит углеводов и липидов в рационе/нарушение их всасывания. Тут всё понятно, если нет субстратов, то откуда взять электроны?
Разобщители окислительного фосфорилирования
Мы договорились, что работа дыхательной цепи сопряжена с синтезом АТФ. Но есть молекулы, которые разобщают эти два процесса. Как? Разобщители — это жирорастворимые молекулы, которые могут проходить через внутреннюю мембрану митохондрий. Но если бы они только проходили! Эти молекулы присоединяют к себе протоны, а потом переносят их в матрикс митохондрии. Они мешают митохондрии создать градиент концентрации протонов, который так важен для синтеза АТФ.
В итоге получается так, что дыхательная цепь работает, но синтеза АТФ нет. Энергия транспорта электронов рассеивается в виде тепла. Я хочу, чтобы вы помнили о разобщителях, которые встречаются в нашем организме: жирных кислотах, неконъюгированном билирубине и трийодтиронине. Это не значит, что в норме они сильно влияют на работу окислительного фосфорилирования. Но при патологических состояниях…
У пациента с ожирением в крови повышается количество жирных кислот. Чем больше жирных кислот, тем больше шанс, что они попадут в митохондрии и повлияют на окислительное фосфорилирование.
Некоъюгированный билирубин повышается при гемолизе эритроцитов или нарушениях функции печени. Его накопление приводит к разобщению работы дыхательной цепи и синтеза АТФ. Представьте какую слабость чувствует пациент при гемолизе эритроцитов — в митохондрии не попадает кислород, да ещё непрямой билирубин вызывает разобщение окислительного фосфорилирования. Мрак.
Трийодтиронин повышается при тиреотоксикозе. Поэтому эти пациенты чувствуют жар и быстро теряют в весе — энергия переноса электронов рассеивается в виде тепла.
Мы и сами можем вызывать разобщение, назначив дикумарин — антикоагулянт непрямого действия. Его побочный эффект — это разобщение окислительного фосфорилирования.
Подсчёт АТФ: сколько молекул можно получить с помощью окислительного фосфорилирования?
Теперь, когда мы понимаем все механизмы, можно подсчитать сколько АТФ было получено в ходе катаболизма одной молекулы глюкозы.
В ходе гликолиза было получено две молекулы НАДH. Они транспортируются в матрикс митохондрий — ведь гликолиз шёл в цитоплазме. Для этого используются челночные механизмы. Глицеролфосфатный челнок переносит электроны с НАДH на ФАД — образуется ФАДH2. Малат-аспартатный переносит электроны на митохондриальный НАД+ — переходит в НАДH. Теперь понимаете, почему это важно? Будет получено разное количество АТФ.
Остальные восстановленные коферменты уже находятся в митохондрии, поэтому их электроны можно сразу отправить на дыхательную цепь. Допустим, что мы использовали малат-аспартатный челнок, тогда в матриксе митохондрии находятся: 10 молекул НАДH и 2 молекулы ФАДH2.
10 х 2,5 АТФ + 2 х 1,5 АТФ = 28 молекул АТФ.
Если мы использовали глицеролфосфатный челнок, то в матриксе находятся: 8 молекул НАДH и 4 молекулы ФАДH2.
8 x 2,5 АТФ + 4 х 1,5 АТФ = 26 молекул АТФ.
В ходе окислительного фосфорилирования из одной молекулы глюкозы синтезируется 28 или 26 молекул АТФ.
Но это не общий выход катаболизма глюкозы, потому что в нём были и другие реакции синтеза АТФ — субстратного фосфорилирования. Если вспомнить гликолиз, то там были две таких реакции.
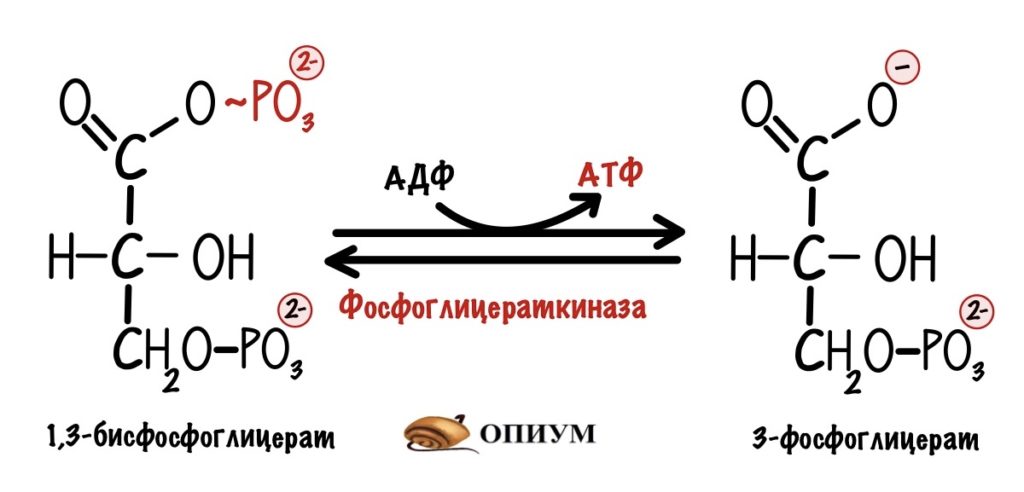
И вот вторая:
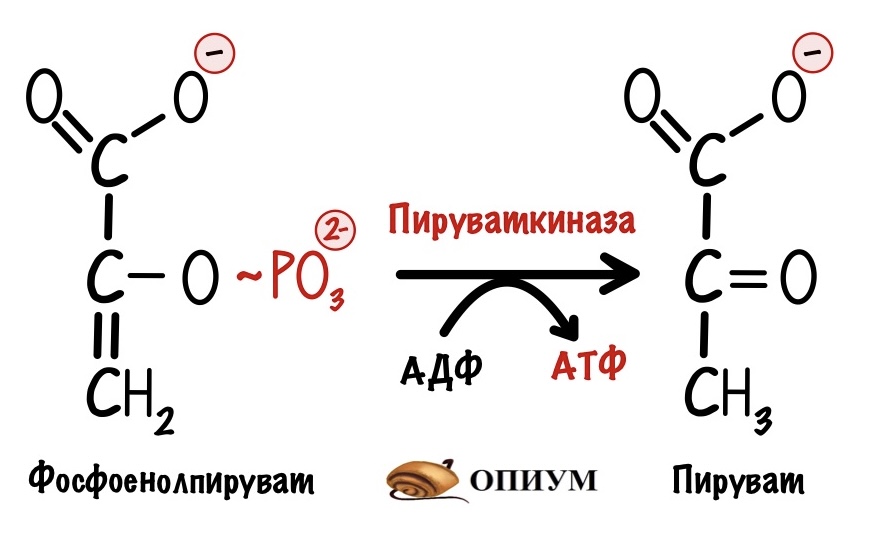
Мы видим, что из одной триозы образуется ещё 2 молекулы АТФ. Но в ходе гликолиза были образованы две триозы, поэтому всего там было получено 4 молекулы АТФ. Но это ещё не всё…
На активацию молекулы глюкозы мы затратили 2 молекулы АТФ — первая и третья реакции гликолиза.
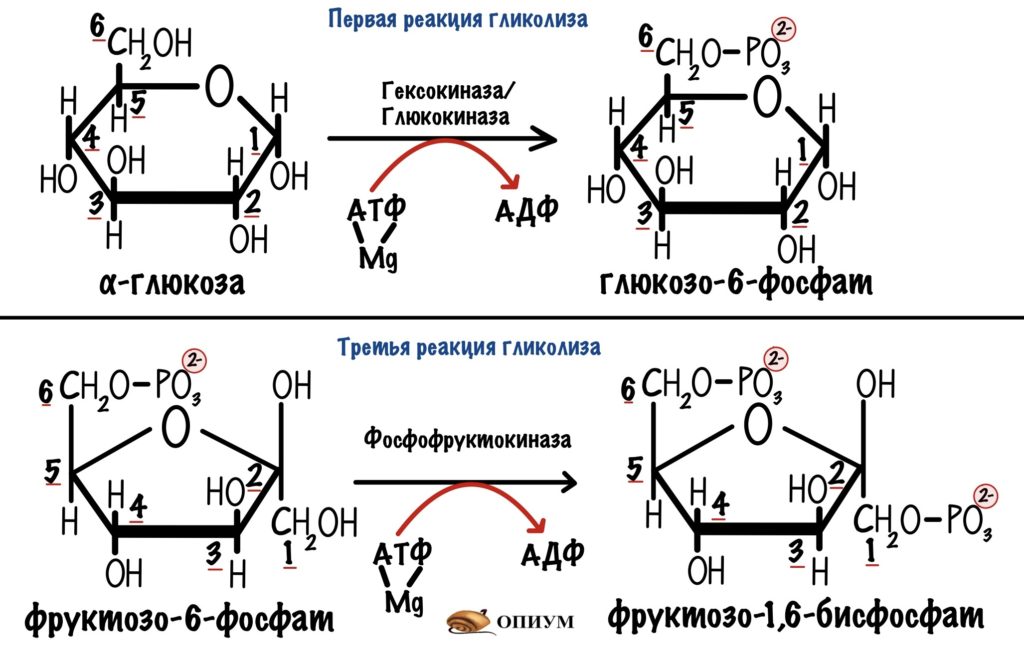
Так что общий выход гликолиза — 2 молекулы АТФ. Но и это ещё не всё!
Вспоминаем цикл трикарбоновых кислот, где были получены ещё две молекулы ГТФ. ГТФ равно АТФ, так как можно легко их переводить друг в друга.

Сделаем общий подсчёт. Гликолиз = 2 АТФ и 2 НАДH. Окислительное декарбоксилирование пирувата = 2 НАДH. ЦТК = 6 НАДH + 2 ФАДH2 + 2 ГТФ (АТФ). Итого:
2 АТФ + 10 НАДH + 2 ФАДH2 + 2 ГТФ (АТФ) = 2 + 25 + 3 + 2 = 32 молекулы АТФ
Это для малат-аспартатного челнока. Если будет использован глицеролфосфатный, то:
2 АТФ + 8 НАДH + 4 ФАДH2 + 2 ГТФ (АТФ) = 2 + 20 + 6 + 2 = 30 молекул АТФ
Из одной молекулы глюкозы образуется 32 молекулы АТФ, либо 30. Всё зависит от челнока, который был использован клеткой.