РАЗМНОЖЕНИЕ ОРГАНИЗМОВ
Размножение — это способность живых существ воспроизводить себе подобных. При этом обеспечивается непрерывность и преемственность жизни. Принято различать два основных типа размножения: бесполое и половое.
Сравнительная характеристика бесполого и полового размножения
| Показатель | Способ размножения | |
| бесполое | половое | |
| Родители | Одна особь | Обычно две особи (разного пола) |
| Потомство | Генетически точная копия родителя (клон) | Генетически отличны от обоих родителей |
| Главный клеточный механизм | Митоз | Мейоз |
| Время возникновения | Раньше полового | Позже бесполого |
| Клеточные источники наследственной информации для развития потомка | Многоклеточные: одна или несколько соматических клеток родителя; одноклеточные: клетка- организм как целое | Родители образуют половые клетки (гаметы) |
| Эволюционное значение | Обеспечивает воспроизведение большого количества идентичных особей, поддерживает наибольшую приспособленность в маломеняющихся условиях обитания, способствует стабилизирующему естественному отбору. Более выгодно в относительно постоянных условиях | Обеспечивает биологическое разнообразие видов, возможность освоения разнообразных условий обитания, увеличивает эволюционные перспективы, способствует движущему естественному отбору. Более выгодно в изменяющихся условиях |
Бесполое размножение
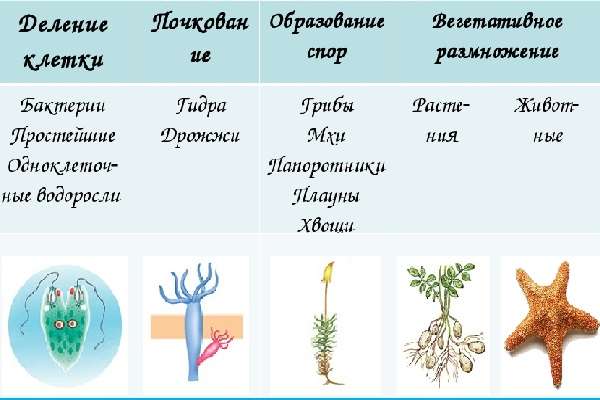
Основными формами бесполого размножения являются деление, спорообразование, почкование, фрагментация и вегетативное размножение. В двух первых случаях новый организм образуется из одной клетки родительской особи, в остальных — из группы клеток.
Формы бесполого размножения
| Форма | Примеры | Характеристика |
| Деление | Свойственна одноклеточным организмам | Самая простая форма бесполого размножения. Исходная материнская клетка делится на две или несколько более или менее одинаковых дочерних клеток. Множественное деление, когда одна материнская клетка даёт начало более чем двум дочерним клеткам, называетсяшизогонией. |
| Споруляция | Встречается у всех растений, грибов и некоторых простейших | Размножение посредством спор. Спора — это мелкая гаплоидная клетка, покрытая защитным покровом (споровой оболочкой), позволяющим переносить действие различных неблагоприятных факторов среды. У многих растений процесс образования спор (спорогенез) осуществляется в особых мешковидных структурах — спорангиях. У многих организмов споры служат не только для размножения, но и для расселения. Споры большинства организмов неподвижны и распространяются пассивно. Но у некоторых водорослей и грибов споры имеют жгутики (зооспоры) и способны активно передвигаться. |
| Почкование | Характерно для кишечнополостных | На теле материнской особи появляется небольшой вырост (почка), а затем происходит отделение (отпочкование) дочерней особи. Почкование многоклеточных организмов не следует путать с формой деления клетки одноклеточных. |
| Фрагментация | Свойственна для плоских, ленточных и кольчатых червей, иглокожих | Заключается в распаде тела многоклеточного организма на две или более части, которые затем превращаются в самостоятельные особи. Фрагментация возможна благодаря регенерации — восстановлению утраченных частей тела. |
| Вегетативное размножение | Характерно для многих групп растений — от водорослей до цветковых | От материнского организма отделяется достаточно хорошо дифференцированная часть (отводки, усы, корневые отпрыски, поросль) или же образуются особые структуры, специально предназначенные для вегетативного размножения (луковицы, клубни, корневища и др.). |
| Клонирование | Искусственный способ размножения, не встречающийся в естественных условиях | Клон — совершенно одинаковое в генетическом отношении потомство, полученное в результате имплантации ядра соматической клетки донора в яйцеклетку. Таким образом, получают зиготу, минуя «классическое» оплодотворение. |

Половое размножение
Половое размножение характерно для подавляющего большинства живых существ. Оно складывается из 4 основных процессов:
- Гаметогенез — образование половых клеток (гамет).
- Оплодотворение — слияние гамет и образование зиготы.
- Эмбриогенез — дробление зиготы и формирование зародыша.
- Постэмбриональный период — рост и развитие организма в послезародышевый период.
Половые клетки
Гаметы — половые клетки, при слиянии которых образуется зигота, из которой развивается новая особь. Гаметы имеют вдвое меньше хромосом, чем остальные клетки тела (соматические клетки). Они не способны делиться в отличие от большинства соматических клеток. Различают женские и мужские половые клетки. Половая принадлежность у высших форм (например, у позвоночных) определяется на генетическом уровне.
Мужские гаметы называются сперматозоидами (если они подвижны) или спермиями (если они лишены жгутикового аппарата и не способны активно передвигаться). Сперматозоиды имеют очень маленькие размеры. Они состоят из головки, шейки, средней части и хвоста (рис. 5.11).
В головке располагается ядро, содержащее ДНК. На переднем конце головки имеется акросома — видоизменённый комплекс Гольджи, который содержит литические ферменты для растворения оболочки яйцеклетки при оплодотворении. Хвост образован микротрубочками и служит для передвижения сперматозоида.
Женские гаметы называются яйцеклетками. Они, как правило, неподвижны, имеют большие, чем сперматозоиды, размеры, хорошо развитую цитоплазму и запас питательных веществ.
Яйцеклетки разных организмов отличаются друг от друга. В зависимости от количества в яйцеклетке желтка их делят на алецитальные, олиголецитальные, мезолецитальные, полилецитальные. В зависимости от характера распределения желтка в яйцеклетке различают гомо- или изолецитальные, телолецитальные, центролецитальные яйцеклетки.
Типы яйцеклеток
| Тип | Характеристика | Организмы |
| Изолецитальные (гомолецитальные) | Относительно мелкие с небольшим количеством равномерно распределённого желтка. Ядро в них располагается ближе к центру | Встречаются у червей, двустворчатых и брюхоногих моллюсков, иглокожих, ланцетника |
| Умеренно телолецитальные | Имеют диаметр около 1,5–2 мм и содержат среднее количество желтка, основная масса которого сосредоточена на одном из полюсов (вегетативном) На противоположном полюсе (анимальном), где желтка мало, находится ядро яйцеклетки | Характерны для осетровых рыб и земноводных |
| Резко телолецитальные | Содержат очень много желтка, занимающего почти весь объём цитоплазмы яйцеклетки. На анимальном полюсе находится зародышевый диск с активной, лишённой желтка цитоплазмой. Размеры этих яиц крупные — 10–15 мм и более. | Встречаются у некоторых рыб, пресмыкающихся, птиц и яйцекладущих млекопитающих |
| Центролецитальные | Характеризуются концентрацией желтка вокруг ядра, расположенного в центре, а периферические слои лишены питательных веществ | Характерны для насекомых |
| Алецитальные | Практически лишены желтка, имеют микроскопически малые размеры (0,1–0,3 мм) | Характерны для плацентарных млекопитающих, в том числе и для человека |
Образование половых клеток
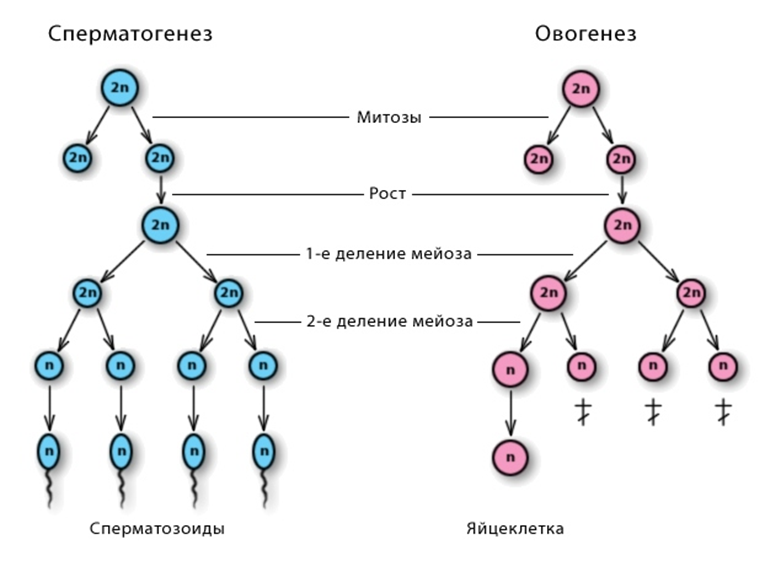
Процесс образования половых клеток — гаметогенез — протекает в половых железах (гонадах). У высших животных женские гаметы образуются в яичниках, мужские — в семенниках. Процесс образования сперматозоидов называют сперматогенезом, яйцеклеток — оогенезом (или овогенезом). Гаметогенез делят на несколько фаз: размножения, роста, созревания и выделяемую при сперматогенезе фазу формирования.
Фазы гаметогенеза
| Стадии | Число хромосом и хроматид | Сперматогенез | Овогенез |
| Размножение | 2n4c | Характеризуется многократными митотическими делениями клеток стенки семенника,приводящими к образованию многочисленныхсперматогоний. Эти клетки диплоидны. Фаза размножения у мужчин начинается с наступлением половой зрелости и продолжается постоянно в течение почти всей жизни | Характеризуется многократными митотическими делениями клеток стенки яичника, приводящими к образованию многочисленных оогоний (овогоний). Эти клетки диплоидны. В женском организме размножение оогоний начинается в эмбриогенезе и завершается к 3-му году жизни. |
| Рост | 2n4c | Сопровождается незначительным увеличением объёма цитоплазмы клеток, незначительным накоплением питательных веществ, необходимых для дальнейших делений, репликацией ДНК и удвоением хромосом. В фазе роста клетки получают названиесперматоцитов I порядка | Сопровождается значительным увеличением объёма цитоплазмы клеток, значительным накоплением питательных веществ, необходимых для дальнейших делений, репликацией ДНК и удвоением хромосом. В фазе роста клетки получают название ооцитов (овоцитов) I порядка |
| Созревание | 1n1c | В результате первого мейотического деления образуются два одинаковыхсперматоцита II порядка, каждый из которых после второго деления мейоза формирует по две сперматиды.В результате фазы созревания из каждой диплоидной клетки формируются 4 гаплоидные сперматиды | Профаза первого мейотического деления осуществляется ещё в эмбриональном периоде, а остальные события мейоза продолжаются после полового созревания организма. Каждый месяц в одном из яичников половозрелой женщины созревает одна яйцеклетка. При этом завершается I деление мейоза, образуются крупный ооцит II порядка и маленькое первое полярное (направительное) тельце, которые вступают во второе деление мейоза На стадии метафазы второго мейотического деления ооцит II порядка овулирует — выходит из яичника в брюшную полость, откуда попадает в яйцевод. Дальнейшее созревание его возможно лишь после слияния со сперматозоидом. Если оплодотворения не происходит, ооцит II порядка погибает и выводится из организма. В случае оплодотворения он завершает второе мейотическое деление, образуя зрелую яйцеклетку — оотиду (овотиду) — и второе полярное тельце. Полярные тельца никакой роли в оогенезе не играют и в конце концов погибают. В результате фазы созревания из каждой диплоидной клетки формируются гаплоидные клетки: 1 оотида и 3 полярных тельца. |
| Формирование | 1n1c | Из каждой сперматиды формируется сперматозоид с головкой, шейкой и хвостом. | Эта стадия отсутствует. |
Оплодотворение
Оплодотворение — это процесс слияния мужской и женской половых клеток (гамет), в результате которого образуется оплодотворённая яйцеклетка (зигота). То есть из двух гаплоидных гамет образуется одна диплоидная клетка (зигота).
Различают наружное оплодотворение, когда половые клетки сливаются вне организма, и внутреннее, когда половые клетки сливаются внутри половых путей особи; перекрёстное оплодотворение, когда объединяются половые клетки разных особей; самооплодотворение — при слиянии гамет, продуцируемых одним и тем же организмом; моноспермию и полиспермию — в зависимости от числа сперматозоидов, оплодотворяющих одну яйцеклетку.
Для большинства видов животных, обитающих или размножающихся в воде, свойственно наружное перекрёстное оплодотворение, которое осуществляется по типу моноспермии. Подавляющее большинство наземных животных и некоторые водные виды имеют внутреннее перекрёстное оплодотворение, причём для части птиц и рептилий характерна полиспермия. Самооплодотворение встречается среди гермафродитов, да и то в исключительных случаях.
У человека процесс оплодотворения происходит в маточной трубе, куда после овуляции попадает ооцит II порядка и могут находиться многочисленные сперматозоиды. При контакте с яйцеклеткой акросома сперматозоида выделяет ферменты, разрушающие оболочки яйцеклетки и обеспечивающие проникновение сперматозоида внутрь. После проникновения сперматозоида яйцеклетка формирует на поверхности толстую непроницаемую оболочку оплодотворения, препятствующую полиспермии.
Проникновение сперматозоида стимулирует ооцит II порядка к дальнейшему делению. Он осуществляет анафазу и телофазу II мейотического деления и становится зрелым яйцом. В результате в цитоплазме яйцеклетки оказываются два гаплоидных ядра, называемых мужским и женским пронуклеусами, которые сливаются с образованием диплоидного ядра — зиготы.
У цветковых растений, кроме слияния гаплоидных гамет — одного из спермиев с яйцеклеткой и образования диплоидной зиготы, из которой развивается зародыш семени, происходит слияние второго спермия с диплоидной вторичной клеткой и образование триплоидных клеток, из которых образуется эндосперм. Этот процесс называется двойным оплодотворением.
Для некоторых групп организмов характерны типы полового размножения (без оплодотворения), один из которых называется партеногенез. Партеногенез — развитие организма из неоплодотворёной яйцеклетки. Характерен для многих общественных насекомых (муравьёв, пчёл, термитов), а также для коловраток, дафний и даже некоторых рептилий. Встречается и у растений (одуванчик).
ИНДИВИДУАЛЬНОЕ РАЗВИТИЕ ОРГАНИЗМОВ
Типы онтогенеза
Онтогенез — индивидуальное развитие организма от зарождения до конца жизни (смерти или нового деления). У видов, размножающихся половым путём, он начинается с оплодотворения яйцеклетки. У видов с бесполым размножением онтогенез начинается с обособления одной клетки или группы клеток материнского организма. У прокариот и одноклеточных эукариотических организмов онтогенез представляет собой, по сути, клеточный цикл, обычно завершающийся делением или гибелью клетки.
Онтогенез есть процесс реализации наследственной информации особи в определённых условиях среды.
Различают два основных типа онтогенеза: прямой и непрямой.
При прямом развитии рождающийся организм в основном сходен со взрослым, а стадия метаморфоза отсутствует.
При непрямом развитии образуется личинка, отличающаяся от взрослого организма внешним и внутренним строением, а также характером питания, способом передвижения и рядом других особенностей.
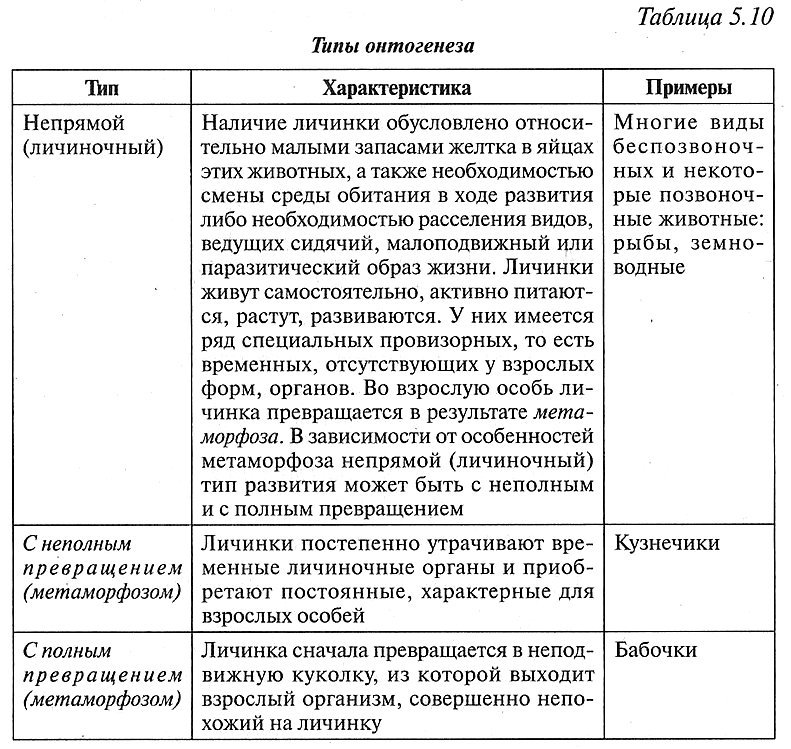
Типы онтогенеза
| Тип | Характеристика | Примеры |
| Непрямой (личиночный) | Наличие личинки обусловлено относительно малыми запасами желтка в яйцах этих животных, а также необходимостью смены среды обитания в ходе развития либо необходимостью расселения видов, ведущих сидячий, малоподвижный или паразитический образ жизни. Личинки живут самостоятельно, активно питаются, растут, развиваются. У них имеется ряд специальных провизорных, то есть временных, отсутствующих у взрослых форм, органов. Во взрослую особь личинка превращается в результате метаморфоза. В зависимости от особенностей метаморфоза непрямой (личиночный) тип развития может быть с неполным и с полным превращением. | Многие виды беспозвоночных и некоторые позвоночные животные: рыбы, земноводные |
| с неполным превращением (метаморфозом) | Личинки постепенно утрачивают временные личиночные органы и приобретают постоянные, характерные для взрослых особей. | Кузнечики |
| с полным превращением (метаморфозом) | Личинка сначала превращается в неподвижную куколку, из которой выходит взрослый организм, совершенно непохожий на личинку. | Бабочки |
| Тип | Характеристика | Примеры |
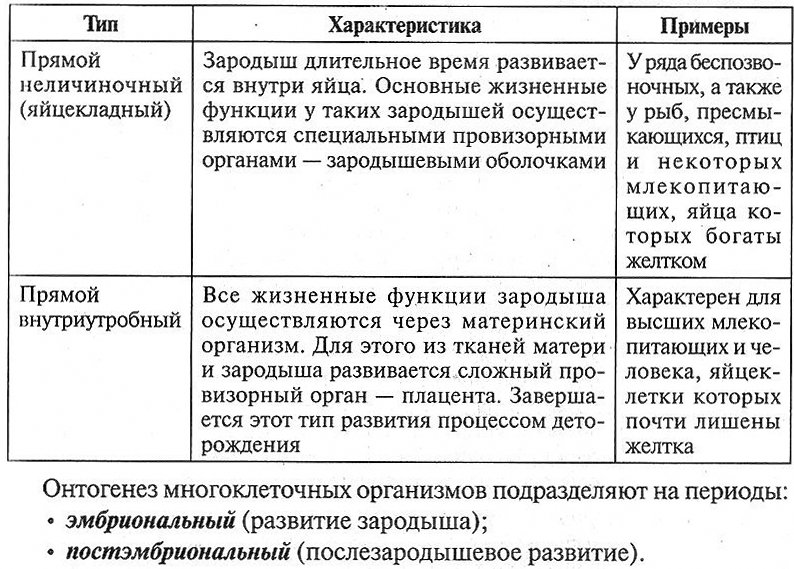
| Прямой неличиночный (яйцекладный) | Зародыш длительное время развивается внутри яйца. Основные жизненные функции у таких зародышей осуществляются специальными провизорными органами — зародышевыми оболочками. | У ряда беспозвоночных, а также у рыб, пресмыкающихся, птиц и некоторых млекопитающих, яйца которых богаты желтком |
| Прямой внутриутробный | Все жизненные функции зародыша осуществляются через материнский организм. Для этого из тканей матери и зародыша развивается сложный провизорный орган — плацента. Завершается этот тип развития процессом деторождения. | Характерен для высших млекопитающих и человека, яйцеклетки которых почти лишены желтка |
Онтогенез многоклеточных организмов подразделяют на периоды:
- эмбриональный (развитие зародыша);
- постэмбриональный (послезародышевое развитие).
Эмбриональное развитие
Эмбриональное развитие (эмбриогенез) начинается с момента оплодотворения, представляет собой процесс преобразования зиготы в многоклеточный организм и завершается выходом из яйцевых или зародышевых оболочек (при личиночном и неличиночном типах развития) либо рождением (при внутриутробном). Эмбриогенез включает процессы дробления, гаструляции, гисто- и органогенеза.
Эмбриогенез
| Этапы | Характеристика |
| Дробление | Ряд последовательных митотических делений зиготы, в результате которых происходит образование бластомеров. Образовавшиеся бластомеры не увеличиваются в размерах. В процессе дробления суммарный объём зародыша не изменяется, а размеры составляющих его клеток уменьшаются. Характер дробления у разных групп организмов различен и определяется типом яйцеклетки. Различают полное дробление, когда зигота дробится целиком, и неполное, когда дробится только часть её. Полное дробление, в свою очередь, бывает равномерным, если образующиеся бластомеры примерно одинаковы по величине, инеравномерным, если они отличаются по размерам. Дробление бывает синхронным илиасинхронным в зависимости от того, одновременно или нет происходит деление бластомеров. В результате ряда дроблений образуется морула, а из неё бластула, или сразу бластула. Морула — многоклеточный зародыш, состоящий из группы тесно прилегающих друг к другу клеток и напоминающий тутовую ягоду. Бластула — многоклеточный шаровидный зародыш с однослойной стенкой и полостью внутри. Бластула образуется в результатебластуляции, когда бластомеры смещаются к периферии, образуя бластодерму, образующаяся при этом внутренняя полость заполняется жидкостью и становится первичной полостью тела — бластоцелью. |
| Гаструляция | Процесс образования двух- или трёхслойного зародыша — гаструлы. Она образуется в результате перемещения клеток бластодермы. Образующиеся слои называютзародышевыми листками. Наружный слой клеток называется эктодермой, внутренний —энтодермой, слой клеток между ними называется мезодермой. Каждый из зародышевых листков дает начало тем или иным органам. В ряде случаев возможно смешанное происхождение. В зависимости от типа бластулы клетки в ходе гаструляции перемещаются по-разному. Выделяют четыре основных способа гаструляции: инвагинация (впячивание), эпиболия(обрастание), иммиграция (проникновение внутрь), деламинация (расслоение), которые в чистом виде почти не встречаются, что даёт основание выделять пятый способ — смешанный(комбинированный). |
| Гисто- и органогенез | Формирование тканей и органов зародыша в результате дифференцировки клеток и зародышевых листков. Дифференцировка — это процесс появления и нарастания морфологических, биохимических и функциональных различий между отдельными клетками и частями развивающегося зародыша. Процесс дифференцировки обеспечивается дифференциальной активностью генов, то есть активностью разных групп генов в различных типах клеток. Из эктодермы образуются нервная система, эпидермис кожи и его производные (роговые чешуи, перья и волосы, зубы).Из мезодермы образуются мускулатура, скелет, выделительная, половая и кровеносная системы. Из энтодермы образуются пищеварительная система и её железы (печень, поджелудочная железа), дыхательная система. |
Постэмбриональное развитие
Постэмбриональное (послезародышевое) развитие начинается с момента рождения (при внутриутробном развитии зародыша у млекопитающих) или с момента выхода организма из яйцевых оболочек и продолжается вплоть до смерти живого организма. Постэмбриональное развитие сопровождается ростом. При этом он может быть ограничен определённым сроком или длиться в течение всей жизни.
ПРИМЕРЫ ЗАДАНИЙ
Часть А
А1. Двуслойное строение текла характерно для
1) кольчатых червей 3) кишечнополостных
2) насекомых 4) простейших
А2. Мезодермы нет у
1) дождевого червя 3) кораллового полипа
2) майского жука 4) крысы
А3. Прямое развитие происходит у
1) лягушки 2) саранчи 3) мухи 4) пчелы
А4. В результате дробления зиготы образуется
1) гаструла 3) нейрула
2) бластула 4) мезодерма
А5. Из энтодермы развивается
1) аорта 2) мозг 3) легкие 4) кожа
А6. Отдельные органы многоклеточного организма закладываются на стадии
1) бластулы 3) оплодотворения
2) гаструлы 4) нейрулы
А7. Бластуляция – это
1) рост клеток
2) многократное дробление зиготы
3) деление клетки
4) увеличение зиготы в размерах
А8. Гаструла зародыша собаки – это:
1) зародыш с образовавшейся нервной трубкой
2) многоклеточный однослойный зародыш с полостью тела
3) многоклеточный трехслойный зародыш с полостью тела
4) многоклеточный двухслойный зародыш
А9. Дифференциация клеток, органов и тканей происходит в результате
1) действия определенных генов в определенное время
2) одновременного действия всех генов
3) гаструляции и бластуляции
4) развития определенных органов
А10.[4]Какая стадия эмбрионального развития позвоночных животных представлена множеством неспециализированных клеток?
1) бластула 3) ранняя нейрула
2) гаструла 4) поздняя нейрула
Часть В
В1. Что из перечисленного относится к эмбриогенезу?
1) оплодотворение 4) сперматогенез
2) гаструляция 5) дробление
3) нейрогенез 6) овогенез
В2. Выберите признаки, характерные для бластулы
1) зародыш, у которого сформирована хорда
2) многоклеточный зародыш с полостью тела
3) зародыш, состоящий из 32 клеток
4) трехслойный зародыш
5) однослойный зародыш с полостью тела
6) зародыш, состоящий из одного слоя клеток
ВЗ. Соотнесите органы многоклеточного зародыша с зародышевыми листками, из которых закладываются эти органы

Часть С
С1. Приведите примеры прямого и непрямого постэмбрионального развития на примере насекомых.
Ответы Онтогенез. Часть А. А1 – 3. А2 – 3. А3 – 2. А4 – 2. А5 – 3. А6 – 4. А7 – 2. А8 – 3. А9 – 1. А10 – 1.
Часть В. В1 – 2, 3, 5. В2 – 2, 5, 6. В3 А – 1; Б – 3; В – 2; Г – 1; Д – 3; Е – 1; Ж – 2.
Часть С. С1 Такие насекомые, как прямокрылые, стрекозы, тли развиваются без метаморфоза, т.е. из яйца развивается личинка, постепенно превращающаяся во взрослую форму – имаго. Жуки, бабочки, мухи и другие насекомые развиваются с метамо– рофозом и проходят в своем развитии стадию куколки.
Индивидуальное развитие организма, или онтогенез, − это совокупность последовательных морфологических, физиологических и биохимических преобразований, претерпеваемых организмом от момента его зарождения до смерти. В онтогенезе происходит реализация наследственной информации, полученной организмом от родителей.
В онтогенезе выделяют два основных периода: эмбриональный и постэмбриональный. В эмбриональном периоде у животных формируется эмбрион, у которого закладываются основные системы органов. В постэмбриональном периоде завершаются формообразовательные процессы, происходит половое созревание, размножение, старение и смерть.
Эмбриональный период
Эмбриональный период начинается с образования зиготы и заканчивается рождением или выходом из яйцевых или зародышевых оболочек молодой особи. Он состоит из трех этапов: дробления, гаструляции и органогенеза.
Начальный этап развития оплодотворенного яйца носит название дробления. Через несколько минут или несколько часов (у разных видов по-разному) после внедрения сперматозоида в яйцеклетку образовавшаяся зигота начинает делиться митозом на клетки, называемые бластомерами. Этот процесс получил название дробления, так как в ходе него число бластомеров увеличивается в геометрической прогрессии, но они не вырастают до размеров исходной клетки, а с каждым делением становятся мельче. Бластомеры, образующиеся при дроблении, представляют собой ранние зародышевые клетки. Во время дробления митозы следуют один за другим, и к концу периода весь зародыш практически не отличается по размерам от зиготы.
Тип дробления яйца зависит от количества желтка и характера его распределения. Различают полное и неполное дробление. В бедных желтком яйцах наблюдается равномерное дробление. Полному дроблению подвергаются зиготы ланцетника и млекопитающих, так как они содержат мало желтка, и он распределен относительно равномерно.
В яйцах, богатых желтком, дробление может быть полным (равномерным и неравномерным) и неполным. Бластомеры одного полюса из-за обилия желтка всегда отстают в темпе дробления от бластомеров другого полюса. Полное, но неравномерное дробление характерно для амфибий. У рыб и птиц дробится лишь часть яйца, расположенная на одном из полюсов; происходит неполное дробление. Часть желтка остается вне бластомеров, которые располагаются на желтке в виде диска.
Рассмотрим более подробно дробление зиготы ланцетника. Дробление охватывает всю зиготу. Борозды первого и второго дробления проходят через полюса зиготы во взаимно перпендикулярных направлениях, в результате чего образуется зародыш, состоящий из четырех бластомеров (Рисунок 1).
Последующие дробления проходят попеременно в продольном и поперечном направлениях. На стадии 32 бластомеров зародыш напоминает ягоду шелковицы или малины. Он называется морулой.
При дальнейшем дроблении (примерно на стадии 128 бластомеров) зародыш расширяется, и клетки, располагаясь однослойно, образуют полый шар. Эта стадия называется бластулой. Стенка однослойного зародыша называется бластодермой, а находящаяся внутри полость — бластоцелью (первичной полостью тела).
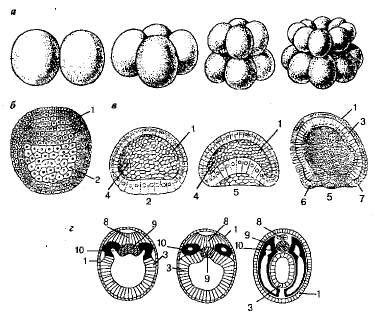
Рисунок 1. Начальные стадии развития ланцетника: а — дробление (стадия двух, четырех, восьми, шестнадцати бластомеров); б — бластула; в — гаструляция; г — схематический поперечный разрез через зародыш ланцетника: 1 — эктодерма; 2 — вегетативный полюс бластулы; 3 — энтодерма; 4 — бластоцель; 5 — рот гаструлы (бластопор); 6,7 — спинная и брюшная губы бластопора; 8 — образование нервной трубки; 9 — образование хорды; 10 — образование мезодермы.
Следующий этап эмбрионального развития — образование двуслойного зародыша — гаструляция. После того как бластула ланцетника полностью сформировалась, дальнейшее дробление клеток особенно интенсивно происходит на одном из полюсов. Вследствие этого они как бы втягиваются (впячиваются) внутрь. В результате образуется двуслойный зародыш. На этой стадии зародыш похож на чашу и называется гаструлой. Наружный слой клеток гаструлы называется эктодермой, или наружным зародышевым листком, а внутренний слой, выстилающий полость гаструлы — гастральную полость (полость первичного кишечника), носит название энтодермы, или внутреннего зародышевого листка. Полость гаструлы, или первичный кишечник, превращается у большинства животных на дальнейших этапах развития в пищеварительный тракт и открывается наружу первичным ртом, или бластопором. У червей, моллюсков и членистоногих бластопор превращается в рот взрослого организма. Поэтому их называют первичноротыми. У иглокожих и хордовых рот прорывается на противоположной стороне, а бластопор превращается в заднепроходное отверстие. Их называют вторичноротыми.
На стадии двух зародышевых листков заканчивается развитие губок и кишечнополостных. У всех остальных животных образуется третий — средний зародышевый листок, расположенный между эктодермой и энтодермой. Он называется мезодермой.
После гаструляции начинается следующий этап в развитии зародыша — дифференцировка зародышевых листков и закладка органов (органогенез). Вначале происходит формирование осевых органов — нервной системы, хорды и пищеварительной трубки. Стадия, на которой осуществляется закладка осевых органов, называется нейрулой.
Нервная система у позвоночных формируется из эктодермы в виде нервной трубки. У хордовых первоначально она имеет вид нервной пластинки. Эта пластинка растет интенсивнее всех остальных участков эктодермы и затем прогибается, образуя желобок. Края желобка смыкаются, возникает нервная трубка, которая тянется от переднего конца к заднему. На переднем конце трубки затем формируется головной мозг. Одновременно с образованием нервной трубки происходит формирование хорды. Хордальный материал энтодермы выгибается, так что хорда выделяется из общей пластинки и превращается в обособленный тяж в виде сплошного цилиндра. Нервная трубка, кишечник и хорда образуют комплекс осевых органов зародыша, который определяет двустороннюю симметрию тела. Впоследствии хорда у позвоночных животных замещается позвоночником, и только у некоторых низших позвоночных ее остатки сохраняются между позвонками даже во взрослом состоянии.
Одновременно с образованием хорды происходит обособление третьего зародышевого листка — мезодермы. Способов образования мезодермы несколько. У ланцетника, например, мезодерма, как и все основные органы, образуется вследствие усиленного деления клеток с двух сторон первичной кишки. В результате образуются два энтодермальных кармана. Эти карманы увеличиваются, заполняя собой первичную полость тела, края их отрываются от энтодермы и смыкаются между собой, образуя две трубки, состоящие из отдельных сегментов, или сомитов. Это и есть третий зародышевый листок — мезодерма. В середине трубок находится вторичная полость тела, или целом.
Дальнейшая дифференцировка клеток каждого зародышевого листка приводит к образованию тканей (гистогенез) и формированию органов (органогенез). Кроме нервной системы из эктодермы развивается наружный покров кожи — эпидермис, и его производные (ногти, волосы, сальные и потовые железы), эпителий рта, носа, анального отверстия, выстилка прямой кишки, эмаль зубов, воспринимающие клетки органов слуха, обоняния, зрения и т. д. (Таблица 1).
Из энтодермы развиваются эпителиальные ткани, выстилающие пищевод, желудок, кишечник, дыхательные пути, легкие или жабры, печень, поджелудочную железу, эпителий желчного и мочевого пузыря, мочеиспускательного канала, щитовидную и околощитовидную железы.
Производными мезодермы являются соединительнотканная основа кожи (дерма), вся собственно соединительная ткань, кости скелета, хрящи, кровеносная и лимфатическая системы, дентин зубов, брыжейка, почки, половые железы, мускулатура.
Таблица 1. Органы и ткани, образующиеся из зародышевых листков
|
Эктодерма |
Энтодерма |
Мезодерма |
|
Эпидермис кожи |
Хорда |
Гладкая мускулатура всех органов |
|
Ногти |
Эпителий желудка, пищевода, кишечника |
Скелетная мускулатура |
|
Волосы |
Эпителий трахеи, бронхов, лёгких |
Сердечная мышца |
|
Потовые железы |
Печень |
Соединительная ткань |
|
Вся нервная система: головной мозг, спинной мозг, ганглии, нервы |
Поджелудочная железа |
Кости |
|
Рецепторные клетки органов чувств |
Эпителий желчного пузыря |
Хрящи |
|
Хрусталик глаза |
Щитовидная и паращитовидная железа |
Дентин зубов |
|
Зубная эмаль |
Эпителий мочевого пузыря |
Кровь |
|
Эпителий мочеиспускательного канала |
Кровеносные сосуды |
|
|
Брыжейка |
||
|
Почки |
||
|
Семенники и яичники |
Зародыш животных развивается как единый организм, в котором все клетки, ткани и органы находятся в тесном взаимодействии. При этом один зачаток оказывает влияние на другой, в значительной мере определяя путь его развития. Кроме того, на темпы роста и развития зародыша оказывают влияние внешние и внутренние условия.
Зародышевые оболочки
Эмбриональное развитие организмов протекает по-разному у разных типов животных, но во всех случаях необходимая связь зародыша со средой обеспечивается специальными внезародышевыми органами, функционирующими временно и называемыми провизорными. Примерами таких временных органов являются желточный мешок у личинок рыб, плацента у млекопитающих.
Развитие зародышей высших позвоночных животных, в том числе и человека, на ранних стадиях развития весьма похоже на развитие ланцетника, но у них, уже начиная со стадии бластулы, наблюдается появление специальных зародышевых органов — дополнительных зародышевых оболочек (хориона, амниона и аллантоиса), обеспечивающих защиту развивающегося зародыша от высыхания и различного рода воздействий среды.
Наружная часть сферического образования, развивающегося вокруг бластулы, называется хорионом (Рисунок 2). Эта оболочка покрыта ворсинками. У плацентарных млекопитающих хорион вместе со слизистой оболочкой матки образует детское место, или плаценту, обеспечивающую связь плода с материнским организмом.
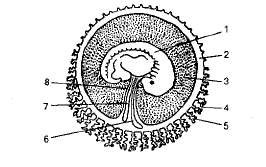
Рисунок 2. Схема зародышевых оболочек: 1 — зародыш; 2 — амнион и его полость (3), заполненная амниотической жидкостью; 4— хорион с ворсинками, образующими детское место (5); 6 — пупочный или желточный пузырь; 7 — аллантоис; 8 — пуповина.
Второй зародышевой оболочкой является амнион (лат. amnion — околозародышевый пузырь). Амнион зародыша заполнен жидкостью. Амниотическая жидкость — водный раствор белков, сахаров, минеральных солей, содержащий также гормоны. Количество этой жидкости у шестимесячного зародыша человека достигает 2 л, а к моменту родов — 1 л. Стенка амниотической оболочки — производное экто- и мезодермы.
Аллантоис (лат. alios — колбаса, oidos — вид) — третья зародышевая оболочка. Это зачаток мочевого мешка. Появляясь в виде небольшого мешковидного выроста на брюшной стенке задней кишки, он выходит через пупочное отверстие и очень быстро разрастается и охватывает амнион и желточный мешок. У различных позвоночных животных его функции различны. У пресмыкающихся и птиц в нем накапливаются продукты жизнедеятельности зародыша до вылупливания из яйца. У зародыша человека он не достигает больших размеров и исчезает на третьем месяце эмбрионального развития.
Постэмбриональное развитие
Органогенез завершается в основном к концу эмбрионального периода развития. Однако дифференцировка и усложнение органов продолжается и в постэмбриональном периоде.
После рождения или выхода из яйцевых оболочек наступает постэмбриональный, или послезародышевый, период онтогенеза. В постэмбриональном развитии можно выделить следующие периоды.
I. Ювенильный период (до полового созревания). Он протекает в зависимости от типа онтогенеза. В этом периоде различают прямое и непрямое развитие. Прямое развитие характерно для организмов с неличиночным и внутриутробным типом развития, непрямое – для организмов с личиночным типом развития.
При прямом развитии рождающийся организм имеет все органы, свойственные взрослому животному. Прямое (неличиночное) развитие характерно для рыб, пресмыкающихся и птиц, а также беспозвоночных, яйца которых богаты желтком, т. е. питательным материалом, достаточным для завершения онтогенеза. Прямое развитие осуществляется у высших млекопитающих (внутриутробный тип развития) и происходит не за счет питательных веществ яйцеклетки, а благодаря поступлению их из материнского организма. В связи с этим из тканей матери и зародыша образуются сложные провизорные органы, в первую очередь плацента.
Многим видам животных присуще непрямое развитие (развитие с превращением — метаморфозом). В этом случае эмбриональное развитие приводит к образованию личинки, которая значительно отличается по внешнему и внутреннему строению от взрослого организма, а затем куколки. Куколка, как правило, неподвижна, она не питается. Из нее развивается полностью сформировавшееся взрослое насекомое. В этом случае говорят о полном превращении (бабочки, мухи, комары, стрекозы). У насекомых с неполным превращением происходит постепенное изменение личинки, сходной со взрослым организмом, сопровождающееся линьками и увеличением размеров; стадия куколки отсутствует (кузнечик, саранча, клопы, вши, стрекозы).
Таблица 2. Насекомые с неполным и полным превращением
|
Насекомые с неполным превращением |
Насекомые с полным превращением |
|
Отряд Прямокрылые (кузнечики, сверчки, саранча, медведки) |
Отряд Сетчатокрылые (муравьиный лев, златоглазка) |
|
Отряд Таракановые (чёрный таракан, рыжий прусак, американский таракан) |
Отряд Жуки, или жесткокрылые (жужелицы, жуки, короеды, долгоносики) |
|
Отряд Богомолы (богомолы) |
Отряд Чешуекрылые, или бабочки (капустная белянка, «мёртвая голова», шелкопряд, комнатная моль, мучная моль) |
|
Отряд Привидениевые (палочник, листотелы) |
Отряд Двукрылые (мухи, комары, москиты, мошки) |
|
Отряд Термиты |
Отряд Блохи (собачья блоха) |
|
Отряд Стрекозы |
Отряд Перепончатокрылые (пчёлы, осы, муравьи, наездники) |
|
Отряд Пухоеды (собачья вошь) |
|
|
Отряд Вши (головная вошь, платяная вошь) |
|
|
Отряд Членистохоботные (цикады, листоблошки, червецы, тли) |
В подтипе позвоночных животных развитие с метаморфозом свойственно рыбам и амфибиям. Например, у лягушки из икринки развивается личинка (головастик), которая по строению, образу жизни и среде обитания резко отличается от взрослых животных. Так, у головастика имеются жабры, орган боковой линии, хвост, двухкамерное сердце, один, как у рыб, круг кровообращения. По достижении личинкой определенного уровня развития происходит ее метаморфоз, в процессе которого вырабатываются признаки взрослого организма. Так головастик превращается в лягушку. Наличие личиночной стадии в развитии земноводных обеспечивает им возможность жить в разной среде и использовать разные источники пищи: головастик живет в воде и питается растительной пищей, а лягушка ведет в основном наземный образ жизни и питается животной пищей. Такое явление наблюдается у многих насекомых. Смена среды обитания и, как следствие, смена образа жизни животного при переходе его от личиночной стадии к взрослому организму снижает интенсивность борьбы за существование внутри вида. Кроме того, у некоторых прикрепленных, малоподвижных или паразитических животных свободноплавающая личинка способствует расселению вида, расширению его ареала.
Переход функциональных систем на режим взрослого организма характеризуется интенсивным ростом органов и частей организма, установлением соответствующих пропорций тела. Выделяют два типа роста организма в процессе индивидуального развития:
1. Ограниченный, или определённый. Рост приурочен к определённым стадиям онтогенеза. Например, насекомые растут только в короткий период линек. У человека рост уже прекращается в возрасте около 15 лет. В период полового созревания наблюдается пубертатный скачок роста.
2. Неограниченный, или неопределённый. Такой тип роста наблюдается, например, у рыб, которые растут на протяжении всей жизни.
Рост, с одной стороны, контролируется генетически, а с другой, – зависит от условий, в которых протекает развитие.
II. Зрелый, или пубертатный, период. Характеризуется наибольшей самостоятельной активностью организма в окружающей среде.
III. Период старости. Старение – общебиологическая закономерность, свойственная всем живым организмам. Состояние старости в биологическом понимании достигается благодаря изменениям, составляющим содержание процесса старения. Эти изменения распространяются на все функциональные системы и обнаруживаются на любом структурно-функциональном уровне организации – будь то молекулярный, клеточный, тканевый, органный и организменный. Таким образом, биологический смысл старения заключается в том, что он делает неизбежной смерть. Смерть представляет собой универсальный механизм ограничения определённым пределом участия многоклеточных организмов в репродукции себе подобных. Без смерти не было бы смены поколений – одной из главных предпосылок эволюционного процесса.
Регенерация
Структуры организма образуются не только во время эмбриогенеза и роста организма, но и взамен утраченных участков тела. Регенерация – способность живых организмов со временем восстанавливать повреждённые ткани, а иногда и целые потерянные органы. Различают физиологическую и репаративную регенерацию.
Физиологическая регенерация – это процессы обновления функционирующих тканей и органов. Например, замена слущивающихся поверхностных слоёв кожи новыми, регенерация эпителия матки в связи с менструациями, смена перьев или шерстяного покрова.
Репаративная регенерация – процессы восстановления частей тела, происходящие после различного рода повреждений. Например, восстановление органа на месте повреждённого, образование рубца на месте кожной раны.
От момента образования зиготы и до выхода зародыша из яйцевых оболочек длится эмбриональный период развития.
Дробление зиготы
После того, как произошло оплодотворение — слияние сперматозоида и яйцеклетки, образовавшаяся зигота начинает интенсивно делиться.
Ее множественные митотические деления называют дроблением.
Важная особенность дробления в том, что не происходит увеличение в размере зародыша: клетки дробятся (делятся) настолько
быстро, что не успевают накопить цитоплазматическую массу. Дробление зиготы человека является полным неравномерным
асинхронным.
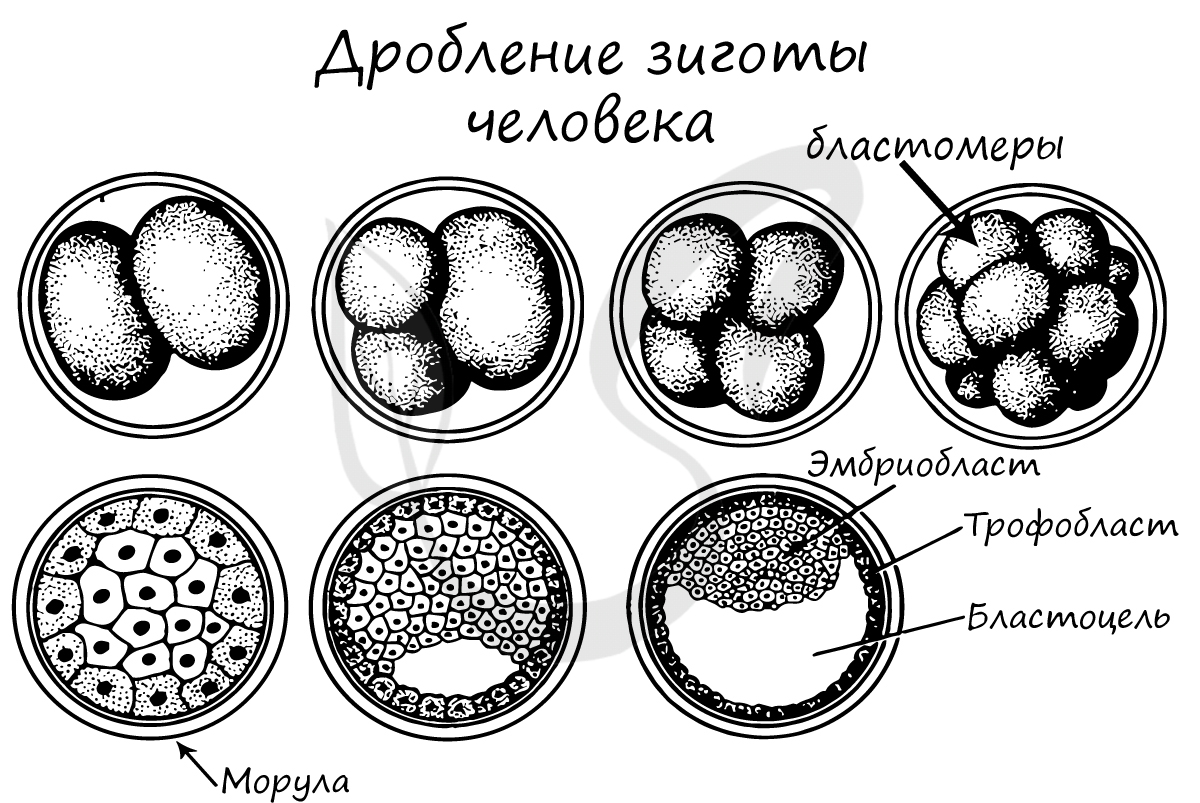
В результате дробления образуется морула. Морула (лат. morum — ягода тутового дерева) — стадия эмбрионального развития на этапе дробления, когда зародыш представляет собой компактную совокупность клеток (без полости внутри).
Бластуляция
Бластуляция — заключительный период дробления, в который зародыш называется бластулой.
После очередных этапов многократного деления образуется однослойный зародыш с полостью внутри — бластула (греч. blastos — зачаток).
Стенки бластулы состоят из бластомеров, которые окружают центральную полость — бластоцель (греч. koilos — полый).
Соединяясь друг с другом, бластомеры образуют бластодерму из одного слоя клеток.

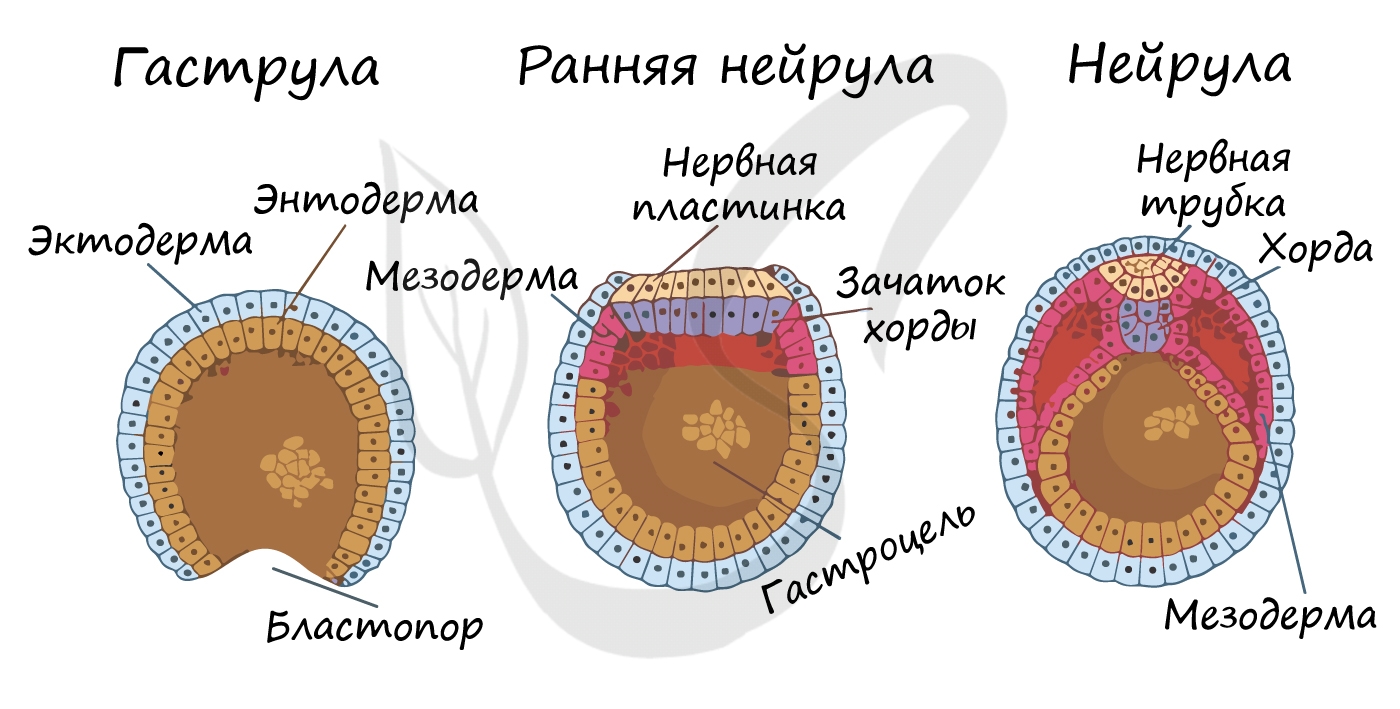
Гаструляция (греч. gaster — желудок, чрево)
Гаструляцией называют стадию эмбрионального развития, в ходе которой клетки, возникшие в результате дробления зиготы,
формируют три зародышевых листка: эктодерму, мезодерму и энтодерму.
Стенка бластулы начинает впячиваться внутрь — происходит инвагинация стенки. По итогу такого впячивания зародыш
становится двухслойным. Двухслойный зародыш называется — гаструла. Полость гаструлы называется гастроцель (полость первичной кишки), а отверстие, соединяющее
гастроцель и внешнюю среду — первичный рот (бластопор).

У первичноротых животных на месте первичного рта (бластопора) образуется ротовое отверстие. К первичноротым относятся:
кишечнополостные, плоские, круглые и кольчатые черви, моллюски, членистоногие.
У вторичноротых на месте бластопора формируется анальное отверстие, а ротовое отверстие образуется на противоположном
полюсе. К вторичноротым относят хордовых и иглокожих (морских звезд, морских ежей).

При впячивании части бластулы (инвагинации) клетки бластодермы мигрируют внутрь и становятся энтодермой (греч. entós — внутренний). Оставшаяся
часть бластодермы снаружи называется эктодермой (греч. ἔκτος — наружный).
Между энто- и эктодермой из группы клеток формируется третий зародышевый листок — мезодерма (греч. μέσος — средний).

Нейрула
Эта стадия следует за гаструлой. Ранняя нейрула представляет собой трехслойный зародыш, состоящий из энто-, экто- и мезодермы. На этапе
нейрулы происходит закладка отдельных органов.
Важно отметить, что на стадии нейрулы происходит процесс нейруляции — закладывание нервной трубки. Нервная пластинка, образовавшаяся
на ранних этапах, прогибается внутрь, при этом ее края сближаются и, замыкаясь, формируют нервную трубку.

Итак, как уже было сказано, на стадии нейрулы закладываются отдельные органы. Эктодерма образует покровный эпителий и нервную пластинку,
мезодерма (из которой в дальнейшем появятся все соединительные ткани), энтодерма — окружает полость первичной кишки (гастроцель), образуя кишечник. От энтодермы отшнуровывается хорда.
Все три зародышевых листка требуют нашего особого внимания, а также понимания того, какие органы и структуры из них образуются.
Эктодерма (греч. ἔκτος — наружный) — наружный зародышевый листок, образует головной и спинной мозг, органы чувств, периферические нервы, эпителий кожи, эмаль зубов, эпителий ротовой полости, эпителий промежуточного и анального отделов прямой кишки, гипофиз, гипоталамус.
Мезодерма (греч. μέσος — средний) — средний зародышевый листок, образует соединительные ткани: кровеносную и лимфатическую системы, костную и хрящевую ткань, мышечные ткани, дентин и цемент зубов, а также выделительную (почки) и половую системы (семенники, яичники).
Энтодерма (греч. entós — «внутренний») — внутренний зародышевый листок, образует эпителий пищевода, желудка, кишечника, трахеи, бронхов, легких, желчного пузыря, мочевого пузыря и мочеиспускательного канала, печень и поджелудочную
железу, щитовидную и паращитовидную железы.

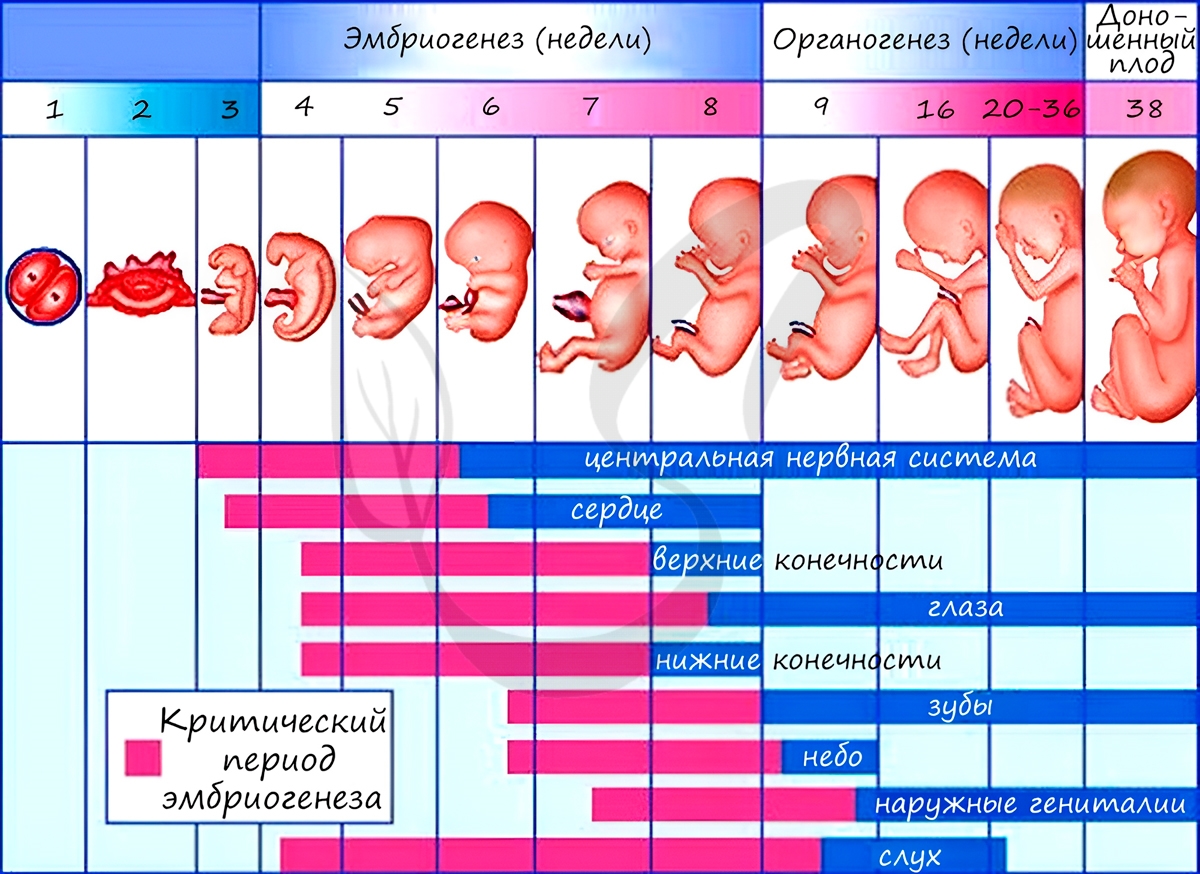
Из зародышевых листков образуются ткани, органы и системы органов. Такой процесс называется органогенезом. В период закладки органов важное значение имеет воздержание матери от вредных привычек (алкоголь, курение), которые могут нарушить процесс дифференцировки клеток и привести к тяжелейшим аномалиям, уродствам плода.
Некоторые лекарства также могут оказывать на плод тератогенный эффект (греч. τέρας — чудовище, урод), приводя к развитию уродств.
Периоды закладки органов и систем органов вследствие их большой важности носят название критических периодов эмбриогенеза.
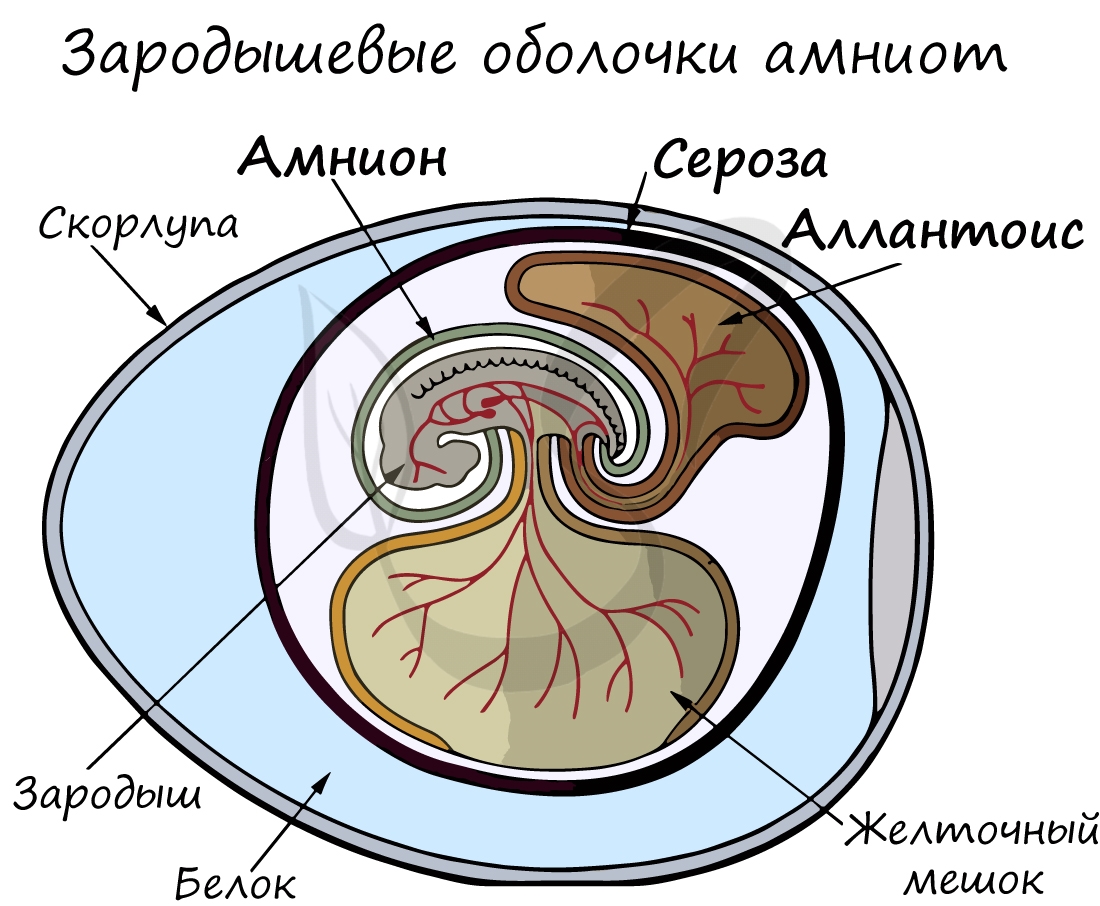
Анамнии и амниоты
Анамнии, или низшие позвоночные — группа животных, не имеющая зародышевых оболочек (зародышевого органа — аллантоиса и амниона).
Анамнии проводят большую часть жизни в воде, без которой невозможно их размножение.
К анамниям относятся рыбы, земноводные.

Амниоты — группа высших позвоночных, характеризующаяся наличием зародышевых оболочек. К амниотам относятся пресмыкающиеся, птицы
и млекопитающие.
Зародышевый орган, аллантоис, является органом дыхания и выделения.
За счет особых оболочек, развивающихся в ходе эмбрионального развития, амниона и серозы, у амниот формируется амниотическая полость.
В ней находится зародыш, окруженный околоплодными водами. Благодаря такому гениальному устройству, амниотам для размножения и
развития более не нужно постоянное нахождение в водоеме, они «обрели независимость» от него.
Развитие плода происходит в мышечном органе — матке, которая, сокращаясь во время родов, стимулирует изгнание плода через
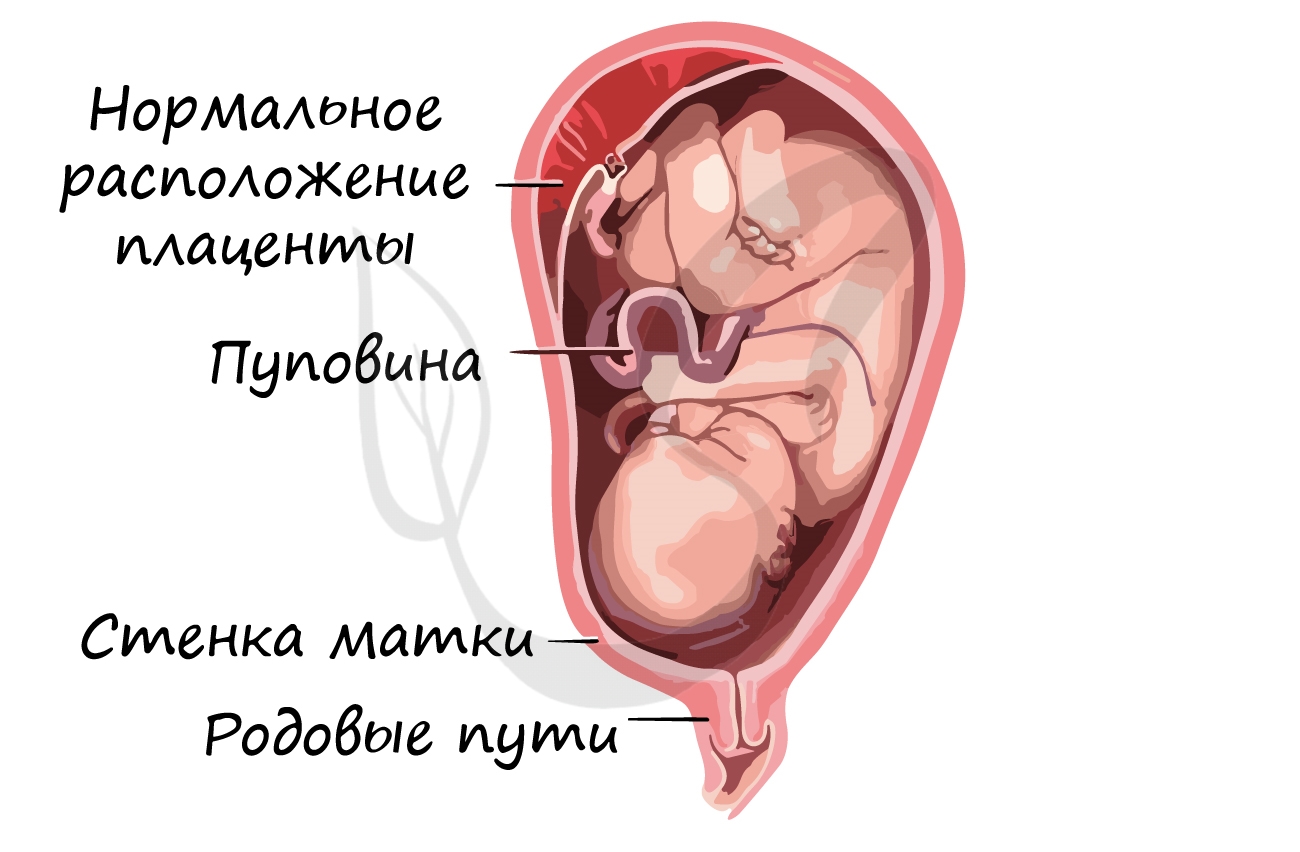
родовые пути. Питание осуществляется через плаценту — «детское место» — орган, который с одной стороны омывается кровью
матери, а с другой — кровью плода. Через плаценту происходит транспорт питательных веществ и газообмен.
Соединяет плаценту и плод особый орган — пуповина, внутри которой проходят артерии, вены.
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Материалы для подготовки к ЕГЭ. Онлайн-Справочник по биологии.
5. Размножение и индивидуальное развитие организмов
ВСЕ РАЗДЕЛЫ СПРАВОЧНИКА
5. Размножение и индивидуальное
развитие организмов
5.1. Деление клеток.
5.1.1. Хромосомный набор.
Хромосомный набор — совокупность хромосом, содержащихся в ядре. В зависимости от хромосомного набора клетки бывают соматическими и половыми (табл. 5.1).


5.1.2. Клеточный цикл
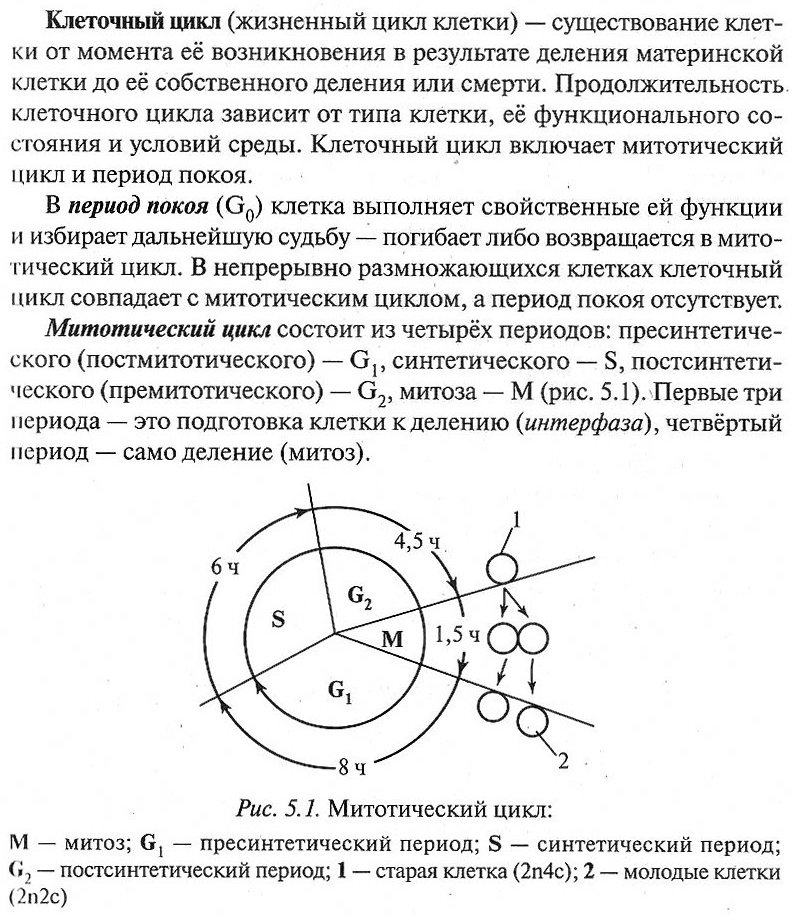
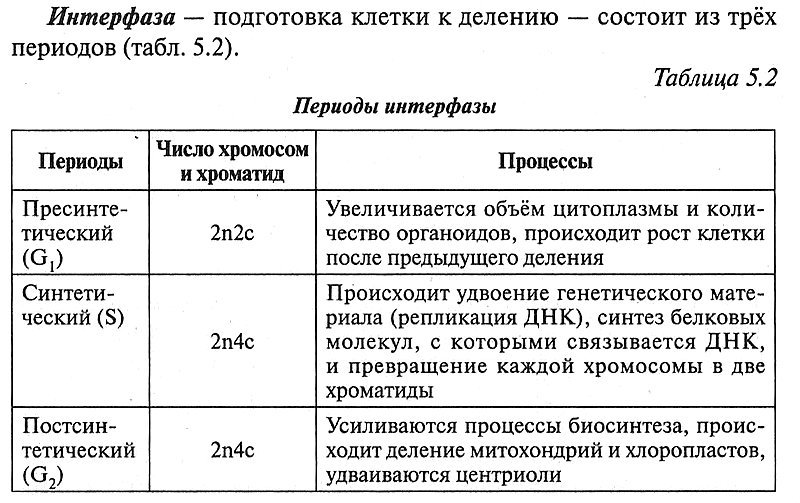
5.1.3. Деление эукариотических клеток
5.1.4. Деление прокариотических клеток


5.2. Размножение организмов
Размножение — это способность живых существ воспроизводить себе подобных. При этом обеспечивается непрерывность и преемственность жизни. Принято различать два основных типа размножения: бесполое и половое (табл. 5.5).
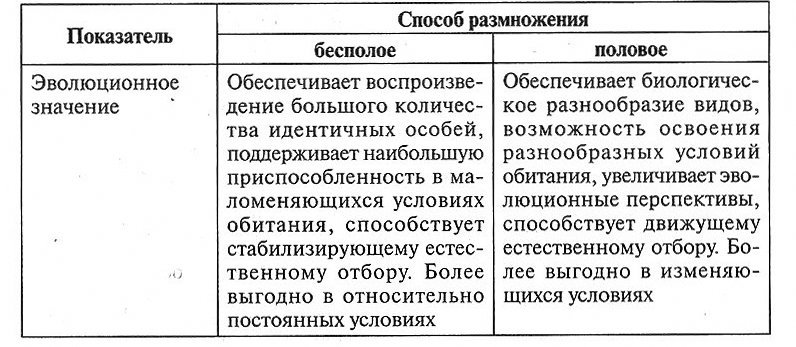
Процесс полового размножения обычно осуществляется между двумя физиологически различными особями — мужской и женской. Они формируют особые половые клетки (гаметы), при слиянии которых образуется зигота. При этом геномы родительских клеток смешиваются, поэтому потомки генетически отличаются от каждого из родителей и друг от друга. У гермафродитов половое размножение может происходить с участием только одной особи, но только в случае отсутствия второй особи этого вида, поскольку получение генетической информации от двух разных организмов эволюционно более предпочтительно.
В процессе бесполого размножения участвует только одна особь. Образования гамет не происходит. Организм либо просто делится на две или более частей, либо формирует специальные структуры, из которых восстанавливаются новые индивиды, генетически идентичные материнской особи.
Бесполое размножение возникло раньше полового. Оно обеспечивает воспроизведение большого количества идентичных особей и более выгодно в относительно постоянных условиях.
Половое размножение появилось более 3 млрд лет назад. При половом размножении происходит объединение генетической информации от двух особей одного вида (родителей) в наследственном материале потомка. То есть биологическое значение полового размножения заключается не только в самовоспроизведении особей, но и в обеспечении биологического разнообразия видов, их адаптивных возможностей и эволюционных перспектив. Это делает половое размножение биологически более прогрессивным, чем бесполое.
5.2.1. Бесполое размножение.
Основными формами бесполого размножения являются деление, спорообразование, почкование, фрагментация и вегетативное размножение (табл. 5.6). В двух первых случаях новый организм образуется из одной клетки родительской особи, в остальных — из группы клеток.
5.2.2. Половое размножение
5.2.3. Типы редукции числа хромосом.
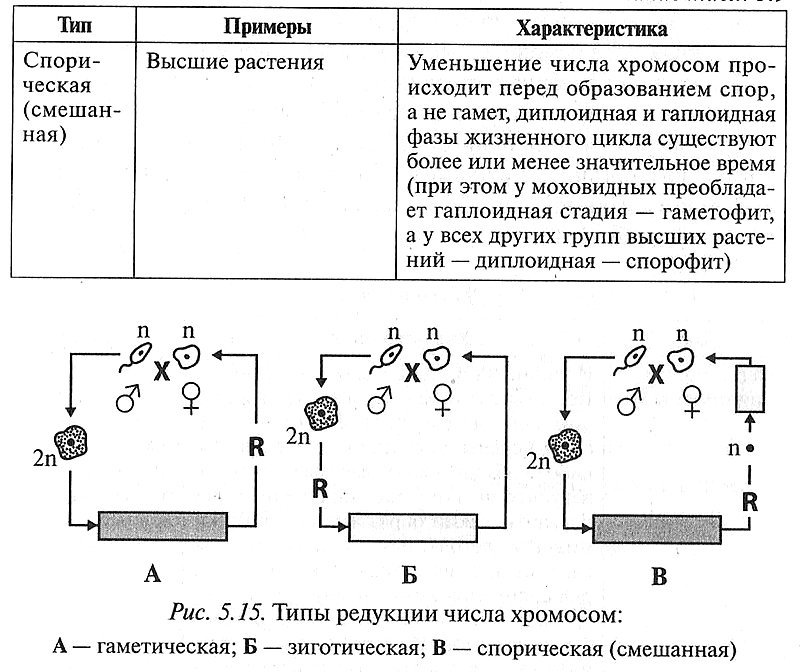
Мейоз и происходящая в ходе него редукция (уменьшение) числа хромосом у разных групп живых организмов происходят в разные периоды жизненного цикла. Существуют три типа редукции числа хромосом: гаметическая, зиготическая, спорическая (смешанная) (табл, 5.9, рис. 5.15).

5.3. Индивидуальное развитие организмов
5.3.1. Типы онтогенеза
Онтогенез — индивидуальное развитие организма от зарождения до конца жизни (смерти или нового деления). У видов, размножающихся половым путём, он начинается с оплодотворения яйцеклетки. У видов с бесполым размножением онтогенез начинается с обособления одной клетки или группы клеток материнского организма. У прокариот и одноклеточных эукариотических организмов онтогенез представляет собой, по сути, клеточный цикл, обычно завершающийся делением или гибелью клетки.
Онтогенез есть процесс реализации наследственной информации особи в определённых условиях среды. Различают два основных типа онтогенеза: прямой и непрямой.
При прямом развитии рождающийся организм в основном сходен со взрослым, а стадия метаморфоза отсутствует.
При непрямом развитии образуется личинка, отличающаяся от взрослого организма внешним и внутренним строением, а также характером питания, способом передвижения и рядом других особенностей (табл. 5.10).
5.3.2. Эмбриональное развитие
Эмбриональное развитие (эмбриогенез) начинается с момента оплодотворения, представляет собой процесс преобразования зиготы в многоклеточный организм и завершается выходом из яйцевых или зародышевых оболочек (при личиночном и неличиночном типах развития) либо рождением (при внутриутробном). Эмбриогенез включает процессы дробления, гаструляции, гисто- и органогенеза (табл. 5.11).
5.3.3. Постэмбриональное развитие
Постэмбриональное (послезародышевое) развитие начинается с момента рождения (при внутриутробном развитии зародыша у млекопитающих) или с момента выхода организма из яйцевых оболочек и продолжается вплоть до смерти живого организма. Постэмбриональное развитие сопровождается ростом. При этом он может быть ограничен определённым сроком или длиться в течение всей жизни.
Все стадии индивидуального развития любого организма подвержены влиянию факторов внешней среды, в которой он формируется. Температура, свет, влажность, разнообразные химические вещества (ядохимикаты, алкоголь, никотин, ряд лекарственных препаратов и др.) могут нарушать нормальный ход онтогенеза и приводить к формированию различных заболеваний и смерти.
ВСЕ РАЗДЕЛЫ СПРАВОЧНИКА
Материалы для подготовки к ЕГЭ. Онлайн-Справочник по биологии.
5. Размножение и индивидуальное развитие организмов.
Просмотров:
14 069
Воспроизведение организмов. Онтогенез.
[su_spoiler title=”Разделы из кодификатора, соответствующие заданию №8″]
Воспроизведение организмов, его значение. Способы размножения, сходство и различие полового и бесполого размножения. Оплодотворение у цветковых растений и позвоночных животных. Внешнее и внутреннее оплодотворение.
Онтогенез и присущие ему закономерности. Эмбриональное и постэмбриональное развитие организмов. Причины нарушения развития организмов.
Закономерности изменчивости. Ненаследственная (модификационная) изменчивость. Норма реакции. Наследственная изменчивость: мутационная, комбинативная. Виды мутаций и их причины. Значение изменчивости в жизни организмов и в эволюции.
Селекция, ее задачи и практическое значение. Вклад Н.И. Вавилова в развитие селекции: учение о центрах многообразия и происхождения культурных растений; закон гомологических рядов в наследственной изменчивости. Методы селекции и их генетические основы. Методы выведения новых сортов растений, пород животных, штаммов микроорганизмов. Значение генетики для селекции. Биологические основы выращивания культурных растений и домашних животных.
Биотехнология, ее направления. Клеточная и генная инженерия, клонирование. Роль клеточной теории в становлении и развитии биотехнологии. Значение биотехнологии для развития селекции, сельского хозяйства, микробиологической промышленности, сохранения генофонда планеты. Этические аспекты развития некоторых исследований в биотехнологии (клонирование человека, направленные изменения генома).
[/su_spoiler]
На самом деле, с частью тем уже приходилось столкнуться еще в 7 задании: изменчивость и размножение. Номера не такие уж страшные. Из себя они, в большинстве своем, представляют соотношение пары наименований с характеристикой или чем-то подобным. Кроме того, встречаются и задания на выбор нескольких вариантов ответа. На «Решу ЕГЭ» упражнений мало, в сумме около 50-60, так что готовьте дополнительные сборники для качественной подготовки.
Заданий относится к повышенному уровню, за правильное выполнение можно получить 2 балла.
Итак, ближе к делу. «Решу ЕГЭ» выделяет 3 раздела:
- Воспроизведение организмов/Биотехнология
- Закономерности наследственности и изменчивости
- Онтогенез. Жизненный цикл растений/Зародышевые листки
Выглядит сложно, но, если разобраться, то все не так плохо. Можно выделить 20 типов заданий, это не так много, потому что несколько заданий могут относиться к одной теме.
Воспроизведение организмов и Биотехнология
Половое и бесполое размножение
[su_note note_color=”#defae6″]
Установите соответствие между двумя основными формами размножения и их признаками.
| ПРИЗНАКИ | ФОРМА РАЗМНОЖЕНИЯ |
|
А) происходит без образования гамет Б) участвует лишь один организм В) происходит слияние гаплоидных ядер Г) образуется потомство идентичное исходной особи Д) у потомства проявляется комбинативная изменчивость Е) происходит с образованием гамет |
1) бесполое 2) половое |
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| А | Б | В | Г | Д | Е |
[/su_note]
Основная черта полового размножения – участие в процессе гамет (половых клеток) двух разнополых организмов, потомство притом получается разнородным, так как проявляется наследственная изменчивость, а именно – комбинативная.
У бесполого размножения, соответственно, все наоборот: потомство идентичное, участвует одна особь.
| А | Б | В | Г | Д | Е |
| 1 | 1 | 2 | 1 | 2 | 2 |
Ответ: 112122.
[su_note note_color=”#defae6″]
Установите соответствие между способом размножения и конкретным примером: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
| ПРИМЕР | СПОСОБ РАЗМНОЖЕНИЯ |
|
А) спорообразование папоротника Б) образование гамет хламидомонады В) образование спор у сфагнума Г) почкование дрожжей Д) нерест рыб |
1) бесполое 2) половое |
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| А | Б | В | Г | Д |
[/su_note]
Половое размножение как-то более понятно, поэтому почему бы не начать с него. Во-первых, найдем слово «гамета» во вариантах ответа. Оно соответствует половому размножению.
Читаем дальше. Нерест рыбы. Это просто. Ну, конечно же, это половое размножение.
Почкование – однозначно бесполое. Очень известный пример данного типа размножения, поэтому вряд ли вызовет вопросы.
Еще одно важное слово, характеризующее половое размножение – «зигота». Зигота – это результат слияния гамет, соответственно, она диплоидна.
И у мха, и у папоротника есть зиготы. В ответах этого слова нет, так что относится к половому размножению эти варианты не могут.
| А | Б | В | Г | Д |
| 1 | 2 | 1 | 1 | 2 |
Ответ: 12112.
Гаметогенез
[su_note note_color=”#defae6″]
Установите соответствие между процессом, происходящим при сперматогенезе, и зоной, в которой происходит данный процесс.
| ПРОЦЕСС | СТАДИЯ СПЕРМАТОГЕНЕЗА |
|
А) митотическое деление первичных половых клеток Б) образование диплоидных сперматогониев В) образование сперматоцитов 1-го порядка Г) мейотическое деление клеток Д) образование гаплоидных сперматид |
1) зона роста 2) зона размножения 3) зона созревания |
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| А | Б | В | Г | Д |
[/su_note]
Гаметогенез-процесс развития и созревания половых клеток: сперматозоидов и яйцеклеток.
На каждой стадии у клеток есть свои названия:
Клетки, получившиеся в результате митозов называются сперматогонии и овогонии.
Клетки, которые делятся первый раз мейозом- сперматоцит и овоциты I порядка.
Второй раз мейоз – сперматоциты и овоциты II порядка.
После второго мейоза сперматоциты становятся сперматидами, а потом- сперматозоидами.
После второго мейоза 3 клетки отмирают, а одна становится яйцеклеткой.
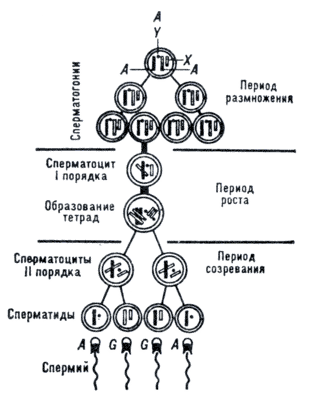
Рассмотрим еще одну схему. Стадия размножения охватывает многократное деление митозом первичных половых клеток.
Увеличение размеров клеток и первый мейоз- стадия роста.
Второй мейоз и до конца – созревание.
Соединим данные с названиями клеток, стадиями и делениями:
| А | Б | В | Г | Д |
| 2 | 2 | 1 | 3 | 3 |
Ответ: 22133.
Типы развития насекомых
- Прямое – ребенок похож на родителя, только меньше по размерам и у него недоразвиты некоторые органы (млекопитающие, птицы).
- Непрямое (с превращением, с метаморфозом) – ребенок (личинка) сильно отличается от родителя (лягушки, насекомые).
При непрямом развитии уменьшена конкуренция между детьми и взрослыми, поскольку они живут в разных местах и питаются разной пищей.
У всех насекомых развитие непрямое, превращение может быть полное и неполное.
Полное: из яйца развивается личинка, она питается, растет, затем превращается в покоящуюся стадию куколку, внутри которой происходит полная перестройка всех органов, из куколки выходит взрослое насекомое (имаго).
Неполное: стадия куколки отсутствует.

[su_note note_color=”#defae6″]
Установите соответствие между насекомым и типом его развития: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
| НАСЕКОМОЕ | ТИП РАЗВИТИЯ |
|
А) медоносная пчела Б) майский жук В) азиатская саранча Г) капустная белянка Д) зеленый кузнечик |
1) с неполным превращением 2) с полным превращением |
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| А | Б | В | Г | Д |
[/su_note]
Для выполнения этого задания нужно не только выучить что такое полноенеполное превращение, но и отряды насекомых (или как минимум по представителю).
| А | Б | В | Г | Д |
| 2 | 2 | 1 | 2 | 1 |
Ответ: 22121.
Различные методы
[su_note note_color=”#defae6″]
Установите соответствие между методом селекции и его использованием в селекции растений и животных.
| МЕТОД | ОБЪЕКТ |
|
А) массовый отбор Б) отбор по экстерьеру В) получение полиплоидов Г) искусственный мутагенез Д) испытание родителей по потомству |
1) селекция растений 2) селекция животных |
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| А | Б | В | Г | Д |
[/su_note]
Начнем рассуждать.
Массовый отбор, наверно, больше характерен для растений нежели, для животных. Растений просто больше и они занимают большие площади территории.
Экстерьер – внешний вид. Это относится к селекции животных. Выведение новых пород основано и на этом тоже, на внешнем виде.
Полиплоидов скорее легче получить у растений, чем у животных. Они проще.
Животных не подвергают искусственному мутагенезу из-за смертности.
Родители, потомство – явно говорится о животных.
| А | Б | В | Г | Д |
| 1 | 2 | 1 | 1 | 2 |
Ответ: 12112.
[su_note note_color=”#defae6″]
Установите соответствие между методами и областями науки и производства, в которых эти методы используются: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
| МЕТОДЫ | ОТРАСЛИ |
|
А) получение полиплоидов Б) метод культуры клеток и тканей В) использование дрожжей для производства белков и витаминов Г) метод рекомбинантных плазмид Д) испытание по потомству Е) гетерозис |
1) селекция 2) биотехнология |
Запишите в таблицу выбранные цифры под соответствующими буквами.
| А | Б | В | Г | Д | Е |
[/su_note]
Разберемся с терминологией:
Биотехнология — это производство необходимых человеку продуктов и материалов с помощью живых организмов, культивируемых клеток и биологических процессов.
Селекция: получение полиплоидов; испытание по потомству; гетерозис. Биотехнология: метод культуры клеток и тканей; использование дрожжей для производства белков и витаминов; метод рекомбинантных плазмид.
Гетерозис — увеличение жизнеспособности гибридов вследствие унаследования определённого набора аллелей различных генов от своих разнородных родителей.
По простому, если обобщить, то в биотехнологии из одного генерируют другое, а при селекции используют существующие материалы и работают с ними.
| А | Б | В | Г | Д | Е |
| 1 | 2 | 2 | 2 | 1 | 1 |
Ответ: 122211.
[su_note note_color=”#defae6″]
Установите соответствие между приёмами и методами биотехнологии: для этого к каждому элементу первого столбца подберите соответствующий элемент из второго столбца.
| ПРИЁМЫ | МЕТОДЫ |
|
А) работа с каллусной тканью Б) введение плазмид в бактериальные клетки В) гибридизация соматических клеток Г) трансплантация ядер клеток Д) получение рекомбинантной ДНК и РНК |
1) клеточная инженерия 2) генная инженерия |
Запишите в таблицу выбранные цифры под соответствующими буквами:
| А | Б | В | Г | Д |
[/su_note]
Здесь все совсем легко: где клетка – там клеточная инженерия, где не клетка- генная. Ну и ткань состоит из клеток.
| А | Б | В | Г | Д |
| 1 | 2 | 1 | 1 | 2 |
Ответ: 12112.
Закономерности наследственности и изменчивости
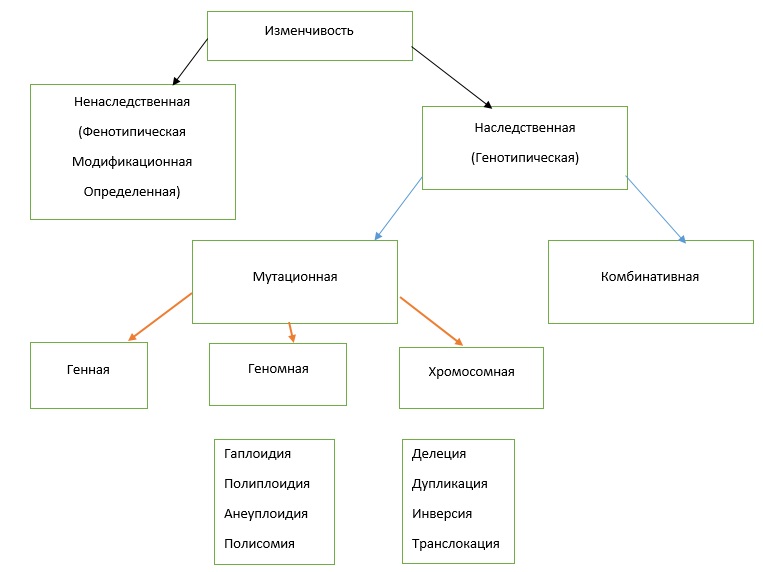
Наследственность — способность организмов передавать свои признаки и особенности развития потомству. Благодаря этой способности все живые существа сохраняют в своих потомках характерные черты вида.
Изменчивость — разнообразие признаков среди представителей данного вида, а также свойство потомков приобретать отличия от родительских форм
Генная мутация– изменения ДНК в пределах одного гена.
Геномная мутация– мутация, в результате которой происходит изменение числа хромосом.
- Гаплоидия– уменьшение в кариотипе соматических клеток числа хромосом вдвое.
- Полиплоидия– увеличение в кариотипе соматических клеток числа хромосом в какое-то количество раз.
- Анеуплоидия– изменение в кариотипе соматических клеток числа хромосом на какое-то число.
- Полисомия– появление в генотипе дополнительных половых хромосом (X,Y).
Хромосомные мутации– изменения в структуре хромосом.
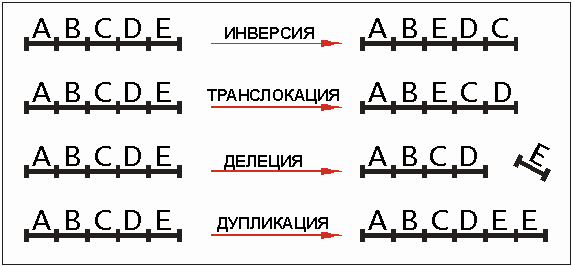
- Делеция– утрата участка хромосомы.
- Дупликация– удвоение участка хромосомы.
- Инверсия– поворот на 180 градусов участка хромосомы
Транслокация– перестановка участка хромосомы на другое место.
Мутации
[su_note note_color=”#defae6″]
Установите соответствие между характеристикой мутации и ее типом.
| ХАРАКТЕРИСТИКА | ТИП МУТАЦИИ |
|
А) включение двух лишних нуклеотидов в молекулу ДНК Б) кратное увеличение числа хромосом в гаплоидной клетке В) нарушение последовательности аминокислот в молекуле белка Г) поворот участка хромосомы на 180 градусов Д) уменьшение числа хромосом в соматической клетке Е) обмен участками негомологичных хромосом |
1) хромосомная 2) генная 3) геномная |
Запишите в таблицу выбранные цифры под соответствующими буквами.
| А | Б | В | Г | Д | Е |
[/su_note]
Разберемся с вариантами ответов:
1.нуклеотиды-генная
2.кратное увеличение числа хромосом-геномная
3.аминокислоты, состоящие из триплетов нуклеотидов- генная
4.поворот хромосомы- хромосомная
5.уменьшение числа хромосом- геномная
6.обмен участками хромосом- хромосомная
| А | Б | В | Г | Д | Е |
| 2 | 3 | 2 | 1 | 3 | 1 |
Ответ:232131.
[su_note note_color=”#defae6″]
Установите соответствие между характеристикой изменчивости и её примерами: к каждой позиции, данной в первом столбце, подберите соответствующую позицию к рисунку из второго столбца.
| ПРИЗНАКИ ОРГАНИЗМА | ОРГАНИЗМ |
|
А) изменчивость носит групповой характер Б) приводит к созданию новых генотипов В) наследуется Г) проявляется в новых комбинациях признаков Д) изменения носят только фенотипический характер Е) изменения определяются нормой реакции |
 |
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| А | Б | В | Г | Д | Е |
[/su_note]
Если у растения листья вытянулись из-за недостатка света, но растение изменилось только внешне, то есть фенотипически. В генотипе у него ничего не изменилось. Это был пример ненаследственной изменчивости, она же фенотипическая, она же модификационная.
Ситуация 2. У животных в размножении участвуют две особи, то в процессе кроссинговера генетический материал как бы перетасовывается, как карты в колоде, но в соответствии с вариантами развития признака. Есть гены, отвечающие, к примеру за цвет глаз. Пусть в роду у одной особи были карие и зеленые глаза, у другой-зеленые и голубые. Какую «карту» вы вытяните- неизвестно. Может, это будет карий и голубой, где карий доминирует. Может- карий и зеленые, где карий доминирует, может, зеленый и зеленый, потомство будет с зелеными глазами. А может, зеленый и голубой, тогда потомство будет с зелеными глазами. И вот это множество комбинаций и называется комбинативной изменчивостью.
Модификационная изменчивость носит групповой характер, а комбинативная- индивидуальный.
| А | Б | В | Г | Д | Е |
| 1 | 2 | 2 | 2 | 1 | 1 |
Ответ: 122211.
Норма реакции
[su_note note_color=”#defae6″]
Установите соответствие между признаком и диапазоном его нормы реакции: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
| ПРИЗНАК | ДИАПАЗОН |
|
А) строение глаза насекомого Б) удойность коровы В) урожайность пшеницы Г) масса тела человека Д) количество пальцев на руках |
1) узкая норма реакции 2) широкая норма реакции |
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам
| А | Б | В | Г | Д |
[/su_note]
Норма реакции — это та максимальная доля от фенотипического значения признака, на которую может изменить признак среда.
Глаз одного вида насекомых не будет кардинально отличаться, норма реакции узкая.
Корова может дать в один день много молока, а в другой- всего ничего, норма широкая.
Один год может быть урожайным, а другой – нет, норма широкая.
Человек весил 55 кг, набрал до 100кг, норма широкая.
Количество пальцев в норме 5, бывают аномалии, но +-1, норма узкая.
| А | Б | В | Г | Д |
| 1 | 2 | 2 | 2 | 1 |
Ответ: 12221.
Онтогенез. Жизненный цикл растений и зародышевые листки.
Стадии развития
Каждый зародышевый листок дает начало чему-то
| Зародышевый листок | Системы органов |
| Эктодерма | Кожа, нервная система, органы чувств |
| Мезодерма | Пищеварительный канал, печень, поджелудочная железа, легкие, хорда |
| Энтодерма | Мышцы, кровь, сосуды, кости, хрящи, гонады |
[su_note note_color=”#defae6″]
Установите соответствие между процессами, происходящими на разных стадиях развития зародыша трёхслойных животных, и стадиями, на которых эти процессы происходят: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
Запишите в таблицу выбранные цифры под соответствующими буквами
| А | Б | В | Г | Д | Е |
[/su_note]
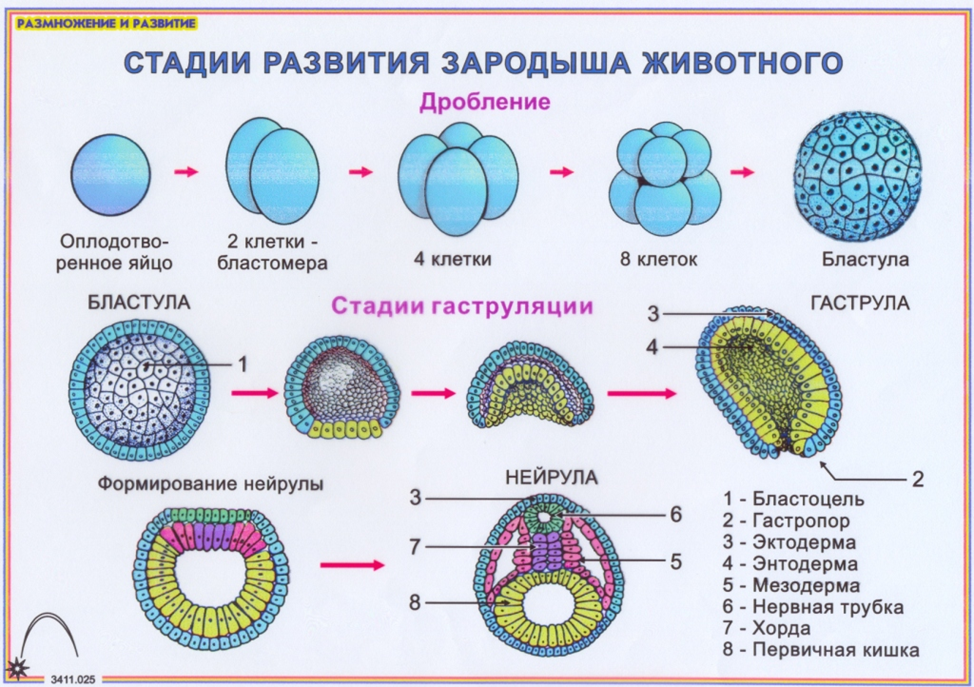
Однослойный зародыш – это бластуляция.
Образование мезодермы происходит в стадию нейрулы.
Двуслойный зародыш – гаструляция.
Вторичная полость – нейрула.
Первичный пузырек – бластула.
Органогенез – нейрула.
| А | Б | В | Г | Д | Е |
| 1 | 3 | 2 | 3 | 1 | 3 |
Ответ: 132313.
Жизненный цикл мха
[su_note note_color=”#defae6″]
Все приведённые ниже признаки, кроме двух, — диплоидные стадии развития мха. Определите два признака, «выпадающих» из общего списка, и запишите в ответ цифры, под которыми они указаны.
- спорофит
- спорангий
- листья
- спора
- зигота
[/su_note]
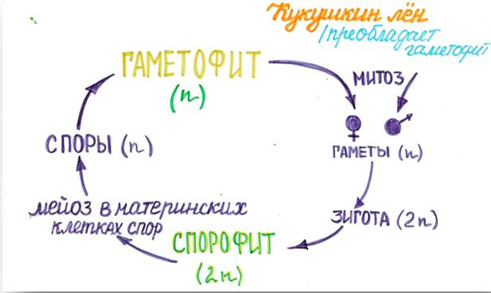
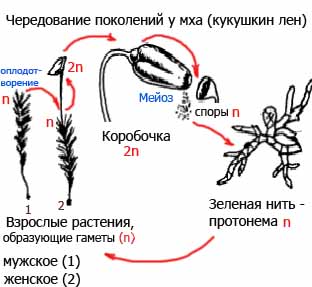
Жизненные циклы остается только выучить.
На схеме видно, что взрослые растения гаплоидны, а это значит, что гаплоидны и листья.
Также гаплоидны споры.
Ответ: 34.
Ксения Алексеевна | Просмотров: 7.4k
Индивидуальное развитие организма, или онтогенез, − это совокупность последовательных морфологических, физиологических и биохимических преобразований, претерпеваемых организмом от момента его зарождения до смерти. В онтогенезе происходит реализация наследственной информации, полученной организмом от родителей.
В онтогенезе выделяют два основных периода: эмбриональный и постэмбриональный. В эмбриональном периоде у животных формируется эмбрион, у которого закладываются основные системы органов. В постэмбриональном периоде завершаются формообразовательные процессы, происходит половое созревание, размножение, старение и смерть.
Эмбриональный период
Эмбриональный период начинается с образования зиготы и заканчивается рождением или выходом из яйцевых или зародышевых оболочек молодой особи. Он состоит из трех этапов: дробления, гаструляции и органогенеза.
Начальный этап развития оплодотворенного яйца носит название дробления. Через несколько минут или несколько часов (у разных видов по-разному) после внедрения сперматозоида в яйцеклетку образовавшаяся зигота начинает делиться митозом на клетки, называемые бластомерами. Этот процесс получил название дробления, так как в ходе него число бластомеров увеличивается в геометрической прогрессии, но они не вырастают до размеров исходной клетки, а с каждым делением становятся мельче. Бластомеры, образующиеся при дроблении, представляют собой ранние зародышевые клетки. Во время дробления митозы следуют один за другим, и к концу периода весь зародыш практически не отличается по размерам от зиготы.
Тип дробления яйца зависит от количества желтка и характера его распределения. Различают полное и неполное дробление. В бедных желтком яйцах наблюдается равномерное дробление. Полному дроблению подвергаются зиготы ланцетника и млекопитающих, так как они содержат мало желтка, и он распределен относительно равномерно.
В яйцах, богатых желтком, дробление может быть полным (равномерным и неравномерным) и неполным. Бластомеры одного полюса из-за обилия желтка всегда отстают в темпе дробления от бластомеров другого полюса. Полное, но неравномерное дробление характерно для амфибий. У рыб и птиц дробится лишь часть яйца, расположенная на одном из полюсов; происходит неполное дробление. Часть желтка остается вне бластомеров, которые располагаются на желтке в виде диска.
Рассмотрим более подробно дробление зиготы ланцетника. Дробление охватывает всю зиготу. Борозды первого и второго дробления проходят через полюса зиготы во взаимно перпендикулярных направлениях, в результате чего образуется зародыш, состоящий из четырех бластомеров (Рисунок 1).
Последующие дробления проходят попеременно в продольном и поперечном направлениях. На стадии 32 бластомеров зародыш напоминает ягоду шелковицы или малины. Он называется морулой.
При дальнейшем дроблении (примерно на стадии 128 бластомеров) зародыш расширяется, и клетки, располагаясь однослойно, образуют полый шар. Эта стадия называется бластулой. Стенка однослойного зародыша называется бластодермой, а находящаяся внутри полость — бластоцелью (первичной полостью тела).
Рисунок 1. Начальные стадии развития ланцетника: а — дробление (стадия двух, четырех, восьми, шестнадцати бластомеров); б — бластула; в — гаструляция; г — схематический поперечный разрез через зародыш ланцетника: 1 — эктодерма; 2 — вегетативный полюс бластулы; 3 — энтодерма; 4 — бластоцель; 5 — рот гаструлы (бластопор); 6,7 — спинная и брюшная губы бластопора; 8 — образование нервной трубки; 9 — образование хорды; 10 — образование мезодермы.
Следующий этап эмбрионального развития — образование двуслойного зародыша — гаструляция. После того как бластула ланцетника полностью сформировалась, дальнейшее дробление клеток особенно интенсивно происходит на одном из полюсов. Вследствие этого они как бы втягиваются (впячиваются) внутрь. В результате образуется двуслойный зародыш. На этой стадии зародыш похож на чашу и называется гаструлой. Наружный слой клеток гаструлы называется эктодермой, или наружным зародышевым листком, а внутренний слой, выстилающий полость гаструлы — гастральную полость (полость первичного кишечника), носит название энтодермы, или внутреннего зародышевого листка. Полость гаструлы, или первичный кишечник, превращается у большинства животных на дальнейших этапах развития в пищеварительный тракт и открывается наружу первичным ртом, или бластопором. У червей, моллюсков и членистоногих бластопор превращается в рот взрослого организма. Поэтому их называют первичноротыми. У иглокожих и хордовых рот прорывается на противоположной стороне, а бластопор превращается в заднепроходное отверстие. Их называют вторичноротыми.
На стадии двух зародышевых листков заканчивается развитие губок и кишечнополостных. У всех остальных животных образуется третий — средний зародышевый листок, расположенный между эктодермой и энтодермой. Он называется мезодермой.
После гаструляции начинается следующий этап в развитии зародыша — дифференцировка зародышевых листков и закладка органов (органогенез). Вначале происходит формирование осевых органов — нервной системы, хорды и пищеварительной трубки. Стадия, на которой осуществляется закладка осевых органов, называется нейрулой.
Нервная система у позвоночных формируется из эктодермы в виде нервной трубки. У хордовых первоначально она имеет вид нервной пластинки. Эта пластинка растет интенсивнее всех остальных участков эктодермы и затем прогибается, образуя желобок. Края желобка смыкаются, возникает нервная трубка, которая тянется от переднего конца к заднему. На переднем конце трубки затем формируется головной мозг. Одновременно с образованием нервной трубки происходит формирование хорды. Хордальный материал энтодермы выгибается, так что хорда выделяется из общей пластинки и превращается в обособленный тяж в виде сплошного цилиндра. Нервная трубка, кишечник и хорда образуют комплекс осевых органов зародыша, который определяет двустороннюю симметрию тела. Впоследствии хорда у позвоночных животных замещается позвоночником, и только у некоторых низших позвоночных ее остатки сохраняются между позвонками даже во взрослом состоянии.
Одновременно с образованием хорды происходит обособление третьего зародышевого листка — мезодермы. Способов образования мезодермы несколько. У ланцетника, например, мезодерма, как и все основные органы, образуется вследствие усиленного деления клеток с двух сторон первичной кишки. В результате образуются два энтодермальных кармана. Эти карманы увеличиваются, заполняя собой первичную полость тела, края их отрываются от энтодермы и смыкаются между собой, образуя две трубки, состоящие из отдельных сегментов, или сомитов. Это и есть третий зародышевый листок — мезодерма. В середине трубок находится вторичная полость тела, или целом.
Дальнейшая дифференцировка клеток каждого зародышевого листка приводит к образованию тканей (гистогенез) и формированию органов (органогенез). Кроме нервной системы из эктодермы развивается наружный покров кожи — эпидермис, и его производные (ногти, волосы, сальные и потовые железы), эпителий рта, носа, анального отверстия, выстилка прямой кишки, эмаль зубов, воспринимающие клетки органов слуха, обоняния, зрения и т. д. (Таблица 1).
Из энтодермы развиваются эпителиальные ткани, выстилающие пищевод, желудок, кишечник, дыхательные пути, легкие или жабры, печень, поджелудочную железу, эпителий желчного и мочевого пузыря, мочеиспускательного канала, щитовидную и околощитовидную железы.
Производными мезодермы являются соединительнотканная основа кожи (дерма), вся собственно соединительная ткань, кости скелета, хрящи, кровеносная и лимфатическая системы, дентин зубов, брыжейка, почки, половые железы, мускулатура.
Таблица 1. Органы и ткани, образующиеся из зародышевых листков
|
Эктодерма |
Энтодерма |
Мезодерма |
|
Эпидермис кожи |
Хорда |
Гладкая мускулатура всех органов |
|
Ногти |
Эпителий желудка, пищевода, кишечника |
Скелетная мускулатура |
|
Волосы |
Эпителий трахеи, бронхов, лёгких |
Сердечная мышца |
|
Потовые железы |
Печень |
Соединительная ткань |
|
Вся нервная система: головной мозг, спинной мозг, ганглии, нервы |
Поджелудочная железа |
Кости |
|
Рецепторные клетки органов чувств |
Эпителий желчного пузыря |
Хрящи |
|
Хрусталик глаза |
Щитовидная и паращитовидная железа |
Дентин зубов |
|
Зубная эмаль |
Эпителий мочевого пузыря |
Кровь |
|
Эпителий мочеиспускательного канала |
Кровеносные сосуды |
|
|
Брыжейка |
||
|
Почки |
||
|
Семенники и яичники |
Зародыш животных развивается как единый организм, в котором все клетки, ткани и органы находятся в тесном взаимодействии. При этом один зачаток оказывает влияние на другой, в значительной мере определяя путь его развития. Кроме того, на темпы роста и развития зародыша оказывают влияние внешние и внутренние условия.
Зародышевые оболочки
Эмбриональное развитие организмов протекает по-разному у разных типов животных, но во всех случаях необходимая связь зародыша со средой обеспечивается специальными внезародышевыми органами, функционирующими временно и называемыми провизорными. Примерами таких временных органов являются желточный мешок у личинок рыб, плацента у млекопитающих.
Развитие зародышей высших позвоночных животных, в том числе и человека, на ранних стадиях развития весьма похоже на развитие ланцетника, но у них, уже начиная со стадии бластулы, наблюдается появление специальных зародышевых органов — дополнительных зародышевых оболочек (хориона, амниона и аллантоиса), обеспечивающих защиту развивающегося зародыша от высыхания и различного рода воздействий среды.
Наружная часть сферического образования, развивающегося вокруг бластулы, называется хорионом (Рисунок 2). Эта оболочка покрыта ворсинками. У плацентарных млекопитающих хорион вместе со слизистой оболочкой матки образует детское место, или плаценту, обеспечивающую связь плода с материнским организмом.
Рисунок 2. Схема зародышевых оболочек: 1 — зародыш; 2 — амнион и его полость (3), заполненная амниотической жидкостью; 4— хорион с ворсинками, образующими детское место (5); 6 — пупочный или желточный пузырь; 7 — аллантоис; 8 — пуповина.
Второй зародышевой оболочкой является амнион (лат. amnion — околозародышевый пузырь). Амнион зародыша заполнен жидкостью. Амниотическая жидкость — водный раствор белков, сахаров, минеральных солей, содержащий также гормоны. Количество этой жидкости у шестимесячного зародыша человека достигает 2 л, а к моменту родов — 1 л. Стенка амниотической оболочки — производное экто- и мезодермы.
Аллантоис (лат. alios — колбаса, oidos — вид) — третья зародышевая оболочка. Это зачаток мочевого мешка. Появляясь в виде небольшого мешковидного выроста на брюшной стенке задней кишки, он выходит через пупочное отверстие и очень быстро разрастается и охватывает амнион и желточный мешок. У различных позвоночных животных его функции различны. У пресмыкающихся и птиц в нем накапливаются продукты жизнедеятельности зародыша до вылупливания из яйца. У зародыша человека он не достигает больших размеров и исчезает на третьем месяце эмбрионального развития.
Постэмбриональное развитие
Органогенез завершается в основном к концу эмбрионального периода развития. Однако дифференцировка и усложнение органов продолжается и в постэмбриональном периоде.
После рождения или выхода из яйцевых оболочек наступает постэмбриональный, или послезародышевый, период онтогенеза. В постэмбриональном развитии можно выделить следующие периоды.
I. Ювенильный период (до полового созревания). Он протекает в зависимости от типа онтогенеза. В этом периоде различают прямое и непрямое развитие. Прямое развитие характерно для организмов с неличиночным и внутриутробным типом развития, непрямое – для организмов с личиночным типом развития.
При прямом развитии рождающийся организм имеет все органы, свойственные взрослому животному. Прямое (неличиночное) развитие характерно для рыб, пресмыкающихся и птиц, а также беспозвоночных, яйца которых богаты желтком, т. е. питательным материалом, достаточным для завершения онтогенеза. Прямое развитие осуществляется у высших млекопитающих (внутриутробный тип развития) и происходит не за счет питательных веществ яйцеклетки, а благодаря поступлению их из материнского организма. В связи с этим из тканей матери и зародыша образуются сложные провизорные органы, в первую очередь плацента.
Многим видам животных присуще непрямое развитие (развитие с превращением — метаморфозом). В этом случае эмбриональное развитие приводит к образованию личинки, которая значительно отличается по внешнему и внутреннему строению от взрослого организма, а затем куколки. Куколка, как правило, неподвижна, она не питается. Из нее развивается полностью сформировавшееся взрослое насекомое. В этом случае говорят о полном превращении (бабочки, мухи, комары, стрекозы). У насекомых с неполным превращением происходит постепенное изменение личинки, сходной со взрослым организмом, сопровождающееся линьками и увеличением размеров; стадия куколки отсутствует (кузнечик, саранча, клопы, вши, стрекозы).
Таблица 2. Насекомые с неполным и полным превращением
|
Насекомые с неполным превращением |
Насекомые с полным превращением |
|
Отряд Прямокрылые (кузнечики, сверчки, саранча, медведки) |
Отряд Сетчатокрылые (муравьиный лев, златоглазка) |
|
Отряд Таракановые (чёрный таракан, рыжий прусак, американский таракан) |
Отряд Жуки, или жесткокрылые (жужелицы, жуки, короеды, долгоносики) |
|
Отряд Богомолы (богомолы) |
Отряд Чешуекрылые, или бабочки (капустная белянка, «мёртвая голова», шелкопряд, комнатная моль, мучная моль) |
|
Отряд Привидениевые (палочник, листотелы) |
Отряд Двукрылые (мухи, комары, москиты, мошки) |
|
Отряд Термиты |
Отряд Блохи (собачья блоха) |
|
Отряд Стрекозы |
Отряд Перепончатокрылые (пчёлы, осы, муравьи, наездники) |
|
Отряд Пухоеды (собачья вошь) |
|
|
Отряд Вши (головная вошь, платяная вошь) |
|
|
Отряд Членистохоботные (цикады, листоблошки, червецы, тли) |
В подтипе позвоночных животных развитие с метаморфозом свойственно рыбам и амфибиям. Например, у лягушки из икринки развивается личинка (головастик), которая по строению, образу жизни и среде обитания резко отличается от взрослых животных. Так, у головастика имеются жабры, орган боковой линии, хвост, двухкамерное сердце, один, как у рыб, круг кровообращения. По достижении личинкой определенного уровня развития происходит ее метаморфоз, в процессе которого вырабатываются признаки взрослого организма. Так головастик превращается в лягушку. Наличие личиночной стадии в развитии земноводных обеспечивает им возможность жить в разной среде и использовать разные источники пищи: головастик живет в воде и питается растительной пищей, а лягушка ведет в основном наземный образ жизни и питается животной пищей. Такое явление наблюдается у многих насекомых. Смена среды обитания и, как следствие, смена образа жизни животного при переходе его от личиночной стадии к взрослому организму снижает интенсивность борьбы за существование внутри вида. Кроме того, у некоторых прикрепленных, малоподвижных или паразитических животных свободноплавающая личинка способствует расселению вида, расширению его ареала.
Переход функциональных систем на режим взрослого организма характеризуется интенсивным ростом органов и частей организма, установлением соответствующих пропорций тела. Выделяют два типа роста организма в процессе индивидуального развития:
1. Ограниченный, или определённый. Рост приурочен к определённым стадиям онтогенеза. Например, насекомые растут только в короткий период линек. У человека рост уже прекращается в возрасте около 15 лет. В период полового созревания наблюдается пубертатный скачок роста.
2. Неограниченный, или неопределённый. Такой тип роста наблюдается, например, у рыб, которые растут на протяжении всей жизни.
Рост, с одной стороны, контролируется генетически, а с другой, – зависит от условий, в которых протекает развитие.
II. Зрелый, или пубертатный, период. Характеризуется наибольшей самостоятельной активностью организма в окружающей среде.
III. Период старости. Старение – общебиологическая закономерность, свойственная всем живым организмам. Состояние старости в биологическом понимании достигается благодаря изменениям, составляющим содержание процесса старения. Эти изменения распространяются на все функциональные системы и обнаруживаются на любом структурно-функциональном уровне организации – будь то молекулярный, клеточный, тканевый, органный и организменный. Таким образом, биологический смысл старения заключается в том, что он делает неизбежной смерть. Смерть представляет собой универсальный механизм ограничения определённым пределом участия многоклеточных организмов в репродукции себе подобных. Без смерти не было бы смены поколений – одной из главных предпосылок эволюционного процесса.
Регенерация
Структуры организма образуются не только во время эмбриогенеза и роста организма, но и взамен утраченных участков тела. Регенерация – способность живых организмов со временем восстанавливать повреждённые ткани, а иногда и целые потерянные органы. Различают физиологическую и репаративную регенерацию.
Физиологическая регенерация – это процессы обновления функционирующих тканей и органов. Например, замена слущивающихся поверхностных слоёв кожи новыми, регенерация эпителия матки в связи с менструациями, смена перьев или шерстяного покрова.
Репаративная регенерация – процессы восстановления частей тела, происходящие после различного рода повреждений. Например, восстановление органа на месте повреждённого, образование рубца на месте кожной раны.
Шпаргалка: Онтогенез понятие типы и основные атрибуты
Онтогенез – это индивидуальное развитие организма, в ходе которого происходит преобразование его морфофизиологических, физиолого-биохимических и цитогенетических признаков. Онтогенез включает две группы процессов: морфогенез и воспроизведение (репродукцию): в результате морфогенеза формируется репродуктивно зрелая особь. Онтогенез характеризуется устойчивостью – гомеорезом. Гомеорез – это стабилизированный поток событий, который представляет собой процесс реализации генетической программы строения, развития и функционирования организма.
С точки зрения эволюции рассматриваются следующие моменты онтогенеза: эмбриональные адаптации; филэмбриогенезы; автономизация онтогенеза; эмбрионизация онтогенеза.
Основные атрибуты онтогенеза
Исходная запрограммированность процессов. Наличие уникальной неизменной генетической программы развития, сформированной вследствие мейоза и оплодотворения
Необратимость онтогенеза. При реализации генетической программы невозможен возврат к предыдущим стадиям
Углубление специализации: по мере развития уменьшается вероятность смены траектории онтогенеза
Адаптивный характер: поливариантность онтогенеза обеспечивает возможность приспособления к различным условиям
Неравномерность темпов: скорость процессов роста и развития изменяется.
Целостность и преемственность отдельных этапов. Признаки, появляющиеся на более поздних стадиях, базируются на признаках, проявляющихся на ранних стадиях
Наличие цикличности: существует цикличность старения и омоложения
Наличие критических периодов, связанных с выбором пути в узловых точках (точках бифуркации) или с преодолением энергетических порогов.
Основные типы онтогенеза
1. Онтогенез организмов с бесполым размножением и/или при зиготном мейозе (прокариоты и некоторые эукариоты).
2. Онтогенез организмов с чередованием ядерных фаз при споровом мейозе (большинство растений и грибов).
3. Онтогенез организмов с чередованием полового и бесполого размножения без смены ядерных фаз. Метагенез – чередование поколений у Кишечнополостных. Гетерогония – чередование партеногенетического и амфимиктического поколений у червей, некоторых членистоногих и низших хордовых.
4. Онтогенез с наличием личиночных и промежуточных стадий: от первично-личиночного анаморфоза до полного метаморфоза. При недостатке питательных веществ в яйце личиночные стадии позволяют завершить морфогенез, а также в ряде случаев обеспечивают расселение особей.
5. Онтогенез с выпадением отдельных стадий. Утрата личиночных стадий и/или стадий бесполого размножения: пресноводные гидры, олигохеты, большинство брюхоногих моллюсков. Утрата конечных стадий и размножение на ранних этапах онтогенеза: неотения.
Таким образом, существует множество основных типов онтогенеза и еще большее число производных типов. В теории эволюции обычно рассматривается онтогенез на примере цветковых растений и позвоночных животных.
Рассматривая морфологическую сторону эволюции онтогенеза, наиболее существенную для палеонтолога, необходимо прежде всего иметь в виду взаимосвязь между изменениями в индивидуальном развитии и эволюцией взрослого организма. Именно эта проблема составляет, например, суть всего учения о филэмбриогенезах А. Н. Северцова [926, 927, 928]. Хотя такое внимание к итогу развития иногда считают чем-то вроде пережитка геккелевского мышления [163, 540] или, в лучшем случае, допустимым методическим приемом [1661], на деле оно неизбежно, если нас интересует реальный эволюционный процесс. Только во взятом изолированно онтогенетическом цикле взрослая стадия может казаться наименее значимой, поскольку она полностью обусловлена предыдущим развитием, но сама уже не определяет ничего. В эволюционном же аспекте дело представляется совсем иначе. Взрослая стадия — единственная, на которой в норме (по крайней мере, у Metazoa) лежит функция размножения, создающего генетическое разнообразие, т. е. материал эволюционного процесса. От успеха взрослых организмов в борьбе за существование зависит, какого сорта гаметы послужат для создания следующего поколения популяции и, следовательно, с каким феноти-пическим материалом будет в дальнейшем иметь дело отбор. В процессе отбора вместе с неудачными фенотипами элиминируются и реализующие их онтогенезы.
Это означает, что какие бы удачные приспособления ни создавались на промежуточных стадиях развития, они не дают преимуществ, если в итоге онтогенез не обеспечивает осуществления нужного фенотипа. Взрослая стадия как бы диктует свои условия всему онтогенезу. Поэтому столь часто критикуемые слова Э. Геккеля о том, что филогения (понимаемая им как совокупность. взрослых стадий) есть причина онтогении, в действительности имеют глубокий смысл, хотя и не совсем тот, который подразумевался автором биогенетического закона.
То, что изменяются все стадии онтогенеза, ни у кого не вызывает сомнений. Вопрос в другом — как соотносятся их изменения? Эволюирует ли взрослая стадия самостоятельно или под действием одновременных морфологических изменений в раннем развитии, или же обоими путями сразу? Наконец, возможно ли в филогенезе обратное воздействие поздних изменений на ход предшествующих стадий — т. е. такое, которое немыслимо в пределах одного онтогенеза?
В ответе на первый вопрос обнаруживается поразительное противоречие в существующих взглядах на роль онтогенеза в эволюции, объединяющих, по сути, две несовместимые концепции. С одной стороны, признается, что нормальное развитие есть зарегулированный процесс, устойчиво направленный к достижению конечного результата и стремящийся свести на нет все возникающие на этом пути уклонения [1192, 1196, 2439, 2441]. Отсюда следует, что эти вариации не должны непосредственно влиять на облик взрослого организма. Эволюция последнего должна быть связана с формообразовательными уклонениями, относящимися к концу развития, где возможности регуляции снижаются. И в то же самое время господствует убеждение, что изменения ранних или промежуточных стадий могут служить причиной немедленного отклонения (девиации) всего хода онтогенеза, радикально меняющего облик взрослой формы у потомков. Обычно этого противоречия даже не замечают. Обе концепции существуют давно и одинаково берут начало в классической морфологии, но лишь первая из них,. как будет показано, может быть согласована с требованиями эволюционной теории.
Первооснову для нее создали два эмпирических обобщения К. М. Бэра [1291]. Одно из них касается уменьшения эмбриональной изменчивости в последовательных стадиях (говорящее, по—мнению К. М. Бэра, о присутствии высшего целеполагающего контроля, регулирующего развитие). Второе составляет известный закон Бэра, или закон зародышевого сходства. В рамках эволюционной теории, как указывал уже Ч. Дарвин [309], это явление означает, что отбор стремится изменять организмы главным образом в позднем возрасте. В дальнейшем эти представления получили поддержку со стороны механики развития, экспериментально показавшей способность морфогенетических процессов к саморегуляции, в результате чего взрослый организм оказывается более устойчивым, чем способ его осуществления (в онтогенезе или при регенерации). Это обобщение, известное как «правило Ру» [921, 2184], или принцип эквифинальности [1524], подтвердило высокую консервативность ранних стадий развития, показывающую, что эволюция взрослой организации должна происходить преимущественно путем изменений в позднем онтогенезе [2185].
Однако главным источником таких представлений послужил основной биогенетический закон Э. Геккеля [1687], представляющий собой переложение на эволюционный язык старой идеи о параллелизме индивидуального развития и «лестницы существ». Выполнение этого закона (т. е. краткое повторение филогенеза в онтогенезе) невозможно представить себе иначе, как’на основе конечных изменений развития.
Поскольку Э. Геккеля интересовала в онтогенезе не столько его эволюция, сколько получение метода для филогенетических реконструкций, то о причинах такого хода изменений мы не находим у него специальных разъяснений, кроме многочисленных и мало что говорящих «законов» наследственности и приспособления. Тем не менее в биогенетическом законе содержится вполне целостная теория эволюции онтогенеза, подразумевающая два положения:
а) взрослая стадия эволюирует посредством прибавления к онтогенезу новых конечных стадий, что обеспечивает рекапитуляцию в онтогенезе облика взрослых предков (палингенез);
б) промежуточные стадии имеют собственную адаптивную эволюцию, искажающую онтогенетическую запись преобразований взрослой стадии (ценогенез).
Биогенетический закон обычно связывают с механоламарк-истскими взглядами Э. Геккеля [359, 565, 1196], иногда утверждая даже, что он был опровергнут самим открытием менделевских факторов и мутационного процесса [1661, 1892]. Однако в действительности вопрос о природе наследственности не играет решающей роли в оценке закона [1822]. В самом деле, независимо от того, связывается ли появление эволюционных новшеств с поздними соматическими изменениями, передаваемыми каким-то образом через половые клетки (ламаркистская концепция) или же с непосредственным изменением самих этих клеток (концепция наследственных факторов), в любом случае новый цикл развития неизбежно должен начинаться с преобразований зиготы, т. е. отличаться от родительского с самого начала.
Э. Геккель ясно понимал это обстоятельство и отнюдь не видел в нем препятствия для биогенетического закона, указывая, что изменения, возникшие в определенный момент развития и передаваемые через родительские гаметы, обнаруживают себя впервые у потомков на той же самой стадии («закон одновременного наследования» [1687]). В этом он прямо следовал Ч. Дарвину [310], сформулировавшему принцип «наследования в соответствующем возрасте» и подчеркивавшему, что появление наследственного уклонения в гаметах и его видимое проявление в индивидуальном развитии у потомства суть разные вещи. Поэтому неудивительно, что признание детерминантов или генов в качестве носителей наследственности не мешало многим исследователям признавать биогенетический закон [1380, 1692, 2472] или, по крайней мере, связь эволюционных изменений с поздними стадиями [503, 709].
Противоположный взгляд на эволюцию онтогенеза, допускающий изменение взрослых организмов путем уклонения на ранних стадиях развития, ведет свое начало от Э. Жоффруа Сент-Илера [2198] и Ф. Мюллера [719]. У них и у современных авторов он чаще всего имеет одно и то же обоснование — сравнение нормальных онтогенезов (или морфогенезов гомологических органов) и отождествление наблюдаемой конечной онтогенетической разницы (зародышевой дивергенции Бэра) с реальным эволюционным процессом. Если, например, морфогенезы гомологичных органов А и Б у двух разных форм совпадают до стадии х, а затем расходятся, то утверждают, что орган Б возник из А путем девиации на стадии х [2141]. Методологическая несостоятельность этой аргументации будет нами показана. Представления о возможности или неизбежности такого пути эволюции никогда не составляли в отличие от геккелевских какой-либо целостной теории. Тем не менее уже к началу XX в. они стали преобладающими.
Одной из причин этого было накопление фактов сравнительной эмбриологии, не согласующихся, по мнению исследователей, с биогенетическим законом. Признание эволюции путем ранних уклонений развития устраняло тезис о первичной неизбежности рекапитуляции, правда, ценой того, что само их существование превратилось в загадку [565, 1869]. Другой момент в критике геккелевских взглядов был связан с растущим убеждением эмбриологов, что подлинное объяснение хода индивидуального развития следует искать в изучении его непосредственных причинных факторов, а не в конструировании умозрительных исторических принципов, подменяющих опытное знание. Этот подход и привел к появлению экспериментальной (каузальной) эмбриологии, которая выявила чрезвычайное разнообразие и специфичность структуры зародышевых клеток у различных организмов. Последнее с очевидностью показывало, что зигота является таким же продуктом эволюции, как и взрослый организм, и что начальная стадия развития высшего организма не есть повторение его одноклеточного предка. «Яйцо курицы не более соответствует начальному звену филогенетической цепи, чем сама курица» [1733]. Все это привело к убеждению, что эволюция происходит путем изменения всего онтогенетического цикла снизу доверху в каждом очередном поколении, а не за счет прибавления конечных стадий [1448, 1763, 2259]. Онтогения, таким образом, творит филогению, а не повторяет ее [1623]. При этом одни авторы имели в виду только постепенные преобразования, другие — возможность резких уклонений, скачкообразно меняющих взрослую форму.
В рамках этих взглядов исчезала, однако, всякая теоретическая возможность появления параллелей между онто- и филогенезом вопреки, казалось бы, очевидным фактам их существования. Это затруднение пытались обойти с помощью утверждения, что такие параллели имеют чисто морфогенетические, а не исторические причины, т. е. что анцестральная взрослая стадия может удерживаться в онтогенезе потомка лишь постольку, поскольку она составляет необходимую базу для реализации последующих стадий [1623]. Другое объяснение состояло в том, что большинство подобных примеров, используемых биогенетическим законом, в действительности относится не к повторению облика взрослых низших форм в онтогенезе высших, а просто к сохранению общих стадий развития у тех и у других, т. е. к проявлениям зародышевого сходства, на что указывал еще К. М. Бэр [1623, 1767, 1970 и др.]. Этот довод, постоянно повторяемый и позднее, например [1317, 1319], обычно считают решающим опровержением геккелевской модели эволюции. Но, как ни странно, при этом не замечают, что при перестройке онтогенеза каждый раз с начальной стадии сохранение зародышевого сходства становится таким же невероятным, как и появление рекапитуляции!
—PAGE_BREAK—
В целом, однако, позиция позднейших исследователей по отношению к биогенетическому закону выглядит более умеренной и допускает возможности разных путей онтогенеза. Соответственно выдвигаются различные классификации этих путей, или модусов [565, 926, 928, 1317, 1319, 1604, 2141, 2124], в которых геккелевский способ эволюции путем поздних надставок занимает то или иное место — от сравнительно большого, например у А. Н. Северцова, до ничтожно малого у Г. де Вира [1317]. Почему онтогенез меняется в разных случаях по-разному, этими взглядами не объясняется (если не считать обычного утверждения, что ранние изменения—это путь к созданию крупных систематических групп). А. Н. Северцов [927] прямо указывает, что ответ на этот вопрос вообще не входит в его задачу, которая состоит лишь в том, чтобы выяснить, как эволюция может идти. Он признает, что выдвигаемые им представления о филэмбриогенезах не зависят от принятия той или иной эволюционной теории.
Само разнообразие выделяемых модусов нередко приводит их авторов к выводу, что все логически мыслимые пути изменений индивидуального развития действительно реализуются в эволюции [2141]. Это равносильно признанию, что в преобразованиях онтогенеза нет сколько-нибудь общих закономерностей. Такой малоутешительный итог столетнего изучения проблемы после Э. Геккеля, в общем, неудивителен в условиях, когда оно основывалось на сравнении одних лишь нормальных онтогенезов, вне связи с анализом механизмов, реализующих материал эволюции — онтогенетическую изменчивость.
Очередным подтверждением этого служит работа С. Гулда [1661] о соотношении онто- и филогенеза, представляющая собой наиболее обширное исследование на эту тему за последние десятилетия. Методология здесь все та же — выделяются эволюционные модусы и оценивается, насколько они отражают ход филогенеза. Попытка связать их с двумя типами адаптивных стратегий (г и 7’С-отбором) не меняет традиционного характера подхода к проблеме. Эволюция онтогенеза остается суммой изолированных процессов, каждый из которых идет в своих особых условиях. Вопрос об их общих первопричинах по-прежнему остается в стороне.
ОНТОГЕНЕЗ И СТАБИЛИЗИРУЮЩИЙ ОТБОР
Морфологическая эволюция онтогенеза не может быть понята, исходя из одних лишь ее адаптивных результатов, т. е. из простого сопоставления «готовых» типов нормального развития. В ее основе лежит процесс эволюции формообразовательных механизмов, преобразующий типичную взрослую организацию (адаптивную норму [1184]), а вместе с ней и стандартный путь ее осуществления в онтогенезе. Каждое элементарное изменение взрослой нормы связано с отбором одного из ее уклонений, т. е. с сохранением любых индивидуальных вариантов развития, реализующих данное выгодное уклонение. Этот отбор варьирующих индивидуальных циклов по принципу однозначности их итога неминуемо должен перестраивать типичный ход развития в целом (см. 7.3), и главная задача эволюционной теории состоит в уяснении закономерностей этой морфогенетической перестройки. Морфологическая эволюция онтогенеза является лишь ее внешним выражением.
Основа для решения этой проблемы содержится в теории стабилизирующего отбора, или эпигенетической концепции эволюции (см. 7.3), согласно которой создание элементарного адаптивного изменения выражается в росте устойчивости онтогенетического осуществления исходной для него вариации.
В свете этих взглядов помехоустойчивость типичного развития, вскрываемая экспериментальной эмбриологией, рассматривается как результат отбора на максимальную защиту нормального формообразования. Механизмы этой защиты (саморегуляции) наиболее хорошо изучены для «регуляционных» онтогенезов (позвоночные, иглокожие и т. д.), где развитие на ранних стадиях идет путем индукционного взаимодействия зачатков, определяющего направление их дальнейшей дифференцировки. Устойчивый ход развития поддерживается за счет широкого раздвигания порогов нормального реагирования зачатков по самым различным параметрам: количеству и активности метаболитов, протяженности периодов компетенции (способности к нормальному взаимодействию) и т. д. Поэтому небольшие колебания в этих показателях, вызываемые генетическими или внешними нарушениями, в достаточно широких пределах забуфериваются и не влияют на дальнейший ход развития. Сильные же воздействия, необратимо нарушающие развитие, ведут либо к гибели, либо к дефинитивным аномалиям, снижающим жизнеспособность.
Такая же устойчивость характерна и для онтогенезов «мозаичного» типа (моллюски, членистоногие и др.), где органогенезы протекают в небольших группах клеток, и регуляция, очевидно, имеет место внутри них. Зачатки здесь в принципе так же муль-типотентны, как и в «регуляционных» онтогенезах (например, зачаток глаза насекомого способен развиваться в антенну или конечность), но нормальная детерминация осуществляется столь надежно, что на первый взгляд кажется единственно возможной. Между двумя типами развития нет четкой грани и в других отношениях: «мозаичные» фазы имеют место в «регуляционных» онтогенезах и наоборот. В целом, благодаря саморегуляции, нормальный онтогенез представляет собой канализированную последовательность событий (креод, или, буквально, «необходимый путь» [2441]), стремящуюся выправить все помехи на пути к осуществлению стандартной организации.
Рассматриваемая нами проблема соотношения эволюции взрослого организма и его онтогенеза вначале должна быть поставлена несколько иначе — каким образом становление новых взрослых особенностей связано с перестройкой их формообразовательных механизмов?
Согласно эпигенетической концепции [1178, 1180, 1181, 1192, 1196], в главных чертах эта связь сводится к следующему. Элементарное эволюционное изменение создается отбором как модификация прежней фенотипической нормы (см. рис. 26,. 27, см. 7.3), осуществляемая лишь в тех условиях, на фоне которых она имеет адаптивное преимущество. Это — примитивное (зависимое) формообразование, в котором среда выступает прямо или косвенно (через посредство функции) как детерминирующий фактор развития. По мере того как новый признак приобретает значение оптимальной адаптации для любого варианта нормальных условий, его развитие становится все более независимым от специфических внешних стимулов, благодаря установлению регуляторных взаимосвязей с другими формообразовательными процессами, составляющими нормальный онтогенез. Реализация адаптивного изменения становится, таким образом, неотъемлемой составной частью осуществления фенотипической нормы как целого. В итоге становление новой элементарной адаптации есть процесс замены внешних факторов ее развития внутренними.
Идея о таком ходе эволюционных изменений принадлежит, как известно, неоламаркизму, но лишь теория стабилизирующего отбора впервые дала ей рациональное обоснование. Адаптивная модификация не возникает одноактно, а создается путем отбора среди элементарных индивидуальных реакций (морфозов), представляющих собой видоспецифичные уклонения нормального развития (см. 7.3). Ее последующее фиксирование—результат селективного преобразования частного варианта нормы в безусловную норму, а не выражение «прямого унаследования». Признаки вообще делятся не на «наследственные» и «приобретенные», а лишь на устойчивые к внешним изменениям и зависимые от них [1194, 1196, 2501]. Унаследование новой особенности не означает ничего иного, кроме стабилизации ее онтогенетического осуществления [1180, 1181].
Прямым экспериментальным подтверждением описанного хода эволюционных преобразований являются опыты К. X. Уоддингтона [2441] по генетической ассимиляции морфозов у дрозофилы, в которых индуцированные внешними воздействиями изменения в ходе отбора превращались в стойкие наследственные признаки (подробнее см. 7.3). На такой путь создания адаптации указывает также и множество наблюдений сравнительного характера. Прежде всего это хорошо известные факты параллелизма между адаптивными модификациями у отдельных видов или рас и устойчивыми признаками у их ближайших родственников, что многократно отмечалось для растений, позвоночных, насекомых и т. д. [405, 1192, 1196]. Естественно предполагать, что мы наблюдаем здесь как бы последовательные стадии стабилизации этих признаков, тем более, что указанные модификации у первых («пластичных») видов обычно возникают при воздействии условий, свойственных в норме для вторых («стабильных»). Другие подобные доказательства дают эмбриология и палеонтология. Например, у позвоночных в позднем развитии очень велика формообразую-щая роль функции, в связи с чем еще В. Ру выделял в их онтогенезе дофункциональный (автономный) и функциональный периоды. Сравнение показывает, что граница между ними сдвигается в эволюции за счет нарастания автономности развития у более высокоорганизованных форм. Так, у водных хвостатых амфибий образование легочных альвеол обусловлено дыханием; у более наземных жаб оно индуцируется гормонально и лишь в конечных стадиях дифференцировки зависит от функции; наконец, у амниот оно протекает вполне автономно в раннем развитии [632].
Во многих случаях устойчивого формообразования первичная зависимость развития от внешних факторов самоочевидна. Например, образование мозолей у млекопитающих и бегающих птиц есть обычная адаптивная реакция кожи на трение. Однако в участках, подвергающихся трению постоянно (например, подошвы конечностей), утолщение эпидермиса наступает еще в эмбриональный период [1196]. Стертая поверхность зубов у дюгоня явно возникла первично на основе жевательных движений, однако в онтогенезе она развивается вне связи с жеванием, путем эмбриональной резорбции поверхности зубов [1317]. Еще один пример—развитие вдавленности на внутренней стороне оборотов раковины наутилоидей. Первоначально она возникла как пассивный результат плотного смыкания оборотов, так что у девонских форм ее еще не было на ранних стадиях развития раковины, предшествующих смыканию. Но уже у некоторых карбоновых видов, как и у современных, вогнутость появляется независимо от механического воздействия — на стадии несвернутой раковины [1776].