Вы уже знаете о том, что гены могут взаимодействовать друг с другом по типу полного и неполного доминирования. Однако,
в генетике встречается масса других примеров взаимодействия генов. В этой статье мы затронем те, которые ранее
не обсуждались.
Кодоминирование
Кодоминирование — взаимодействия аллельных генов, при котором в гетерозиготном состоянии могут оказаться два доминантных
гена одновременно, при этом каждый ген отвечает за свой признак.
Наиболее распространенным примером кодоминирования является наследование групп крови у человека.

Решим пару задач, которые укрепят понимание темы.
Пример решения задачи №1
«Родители имеют II и III группы крови, гетерозиготны. Какие группы крови можно ожидать у их детей?»
Гетерозиготный генотип матери — IAi0 и генотип отца — IBi0. Составим схему
решения для такого случая.

Итак, в результате такого брака может получиться ребенок с любой группой крови, в чем мы убедились.
Пример решения задачи №2
«Дигетерозиготная по B (III) группе и положительному резус-фактору вступила в брак с таким же мужчиной. Какое расщепление
по фенотипу можно ожидать у детей?»
Сходу понятно, что гетерозиготы по III (B) группе крови будут записаны IBi0. Резус-фактор для нас
новое понятие — это белок, находящийся на поверхности эритроцита (тогда говорят, что резус-фактор положителен), или
отсутствующий (тогда у человека резус-фактор считается отрицательным). Генотипы записываются так:
- Резус-фактор положителен: Rh+Rh+, Rh+rh—
- Резус-фактор отрицателен: rh—rh—
В данной задаче сказано, что «дигетерозиготна по … и положительному резус-фактору» — значит, резус фактор будет записывать
в генотипе — Rh+rh—.

Обратите внимание, что ошибкой является записать рецессивный ген перед доминантным. За такое могут снять балл на экзамене:
aA, bB, i0IA, rh—Rh+. Правильный вариант записи: Aa, Bb, IAi0,
Rh+rh—.
Каждая особь образует 4 гаметы, поэтому потомков получается 16. Подсчитает расщепление по фенотипу:
- 9 потомков : положительный резус-фактор, III (B) группа крови
- 3 потомка : положительный резус-фактор, I (0) группа крови
- 3 потомка : отрицательный резус-фактор, III (B) группа крови
- 1 потомок : отрицательный резус-фактор, I (0) группа крови
Расщепление по фенотипу в данном случае получилось: 9:3:3:1. Здесь проявляется III закон Менделя — закон независимого
наследования, так как гены, отвечающие за группу крови и резус-фактор, находятся в разных хромосомах.
Комплементарность
Комплементарность — тип взаимодействия неаллельных генов, при котором развитие признака определяется не одной, а двумя или более парами
неаллельных генов, располагающихся в разных хромосомах.
Неаллельные гены — это гены, расположенные в разных локусах хромосом, которые отвечают за разные признаки. В генетике
случается такое, что один неаллельный ген может влиять на другой (ген a подавляет действие гена B). В этом разделе статьи мы
подробно разберемся с подобным взаимодействием и рассмотрим задачи, которые могут встретиться.
Таким образом, развитие признака определяется именно сочетанием генов друг с другом. Здесь логичнее подчеркнуть
совместное действие генов, нежели чем сказать, что доминантный ген подавляет рецессивный — при комплементарности
это не совсем так.
В каждой задаче свой случай комплементарного взаимодействия генов. Чтобы успешно их решать, надо помнить, что
такое явление, как комплементарность, в принципе, возможно, и быть внимательным при написании генотипов особей и
их гамет.
Пример решения задачи №3
Наследование слуха у человека определяется двумя доминантными генами из разных аллельных пар, один из которых детерминирует развитие слухового нерва,
а другой – улитки. Определить вероятность рождения глухих детей, если оба родителя глухие, но по разным генетическим причинам (у одного отсутствует
слуховой нерв, у другого улитка). По генотипу оба родителя являются дигомозиготными.

Здесь проявляется I закон Менделя — закон единообразия гибридов первого поколения. Возможен только один вариант генотипа ребенка от такого брака.
У ребенка будет развит и слуховой нерв, и улитка — ребенок не будет глухим, в отличие от родителей.
Эпистаз
Эпистаз (противоположное действие генов) — явление, при котором один ген аллельной пары (супрессор) в доминантном (доминантный эпистаз) или
рецессивном (рецессивный эпистаз) состоянии может подавлять развитие признака, за развитие которого отвечает другая пара генов.
Широко известным примером рецессивного эпистаза является Бомбейский феномен, названный так в результате зафиксированного случая в
индийском городе Бомбеи. Доктор Бхенде обнаружил, что у людей рецессивных по гену h (hh) на поверхности эритроцитов не синтезируются
агглютиногены — в результате этого они могут быть универсальными донорами.
Говоря проще о Бомбейском феномене: у людей с генотипом hh всегда обнаруживается первая группа крови при любом генотипе — IAIA,
IBIB, IAIB. Ген h подавляет гены IA и IB — на поверхности эритроцитов не
образуются агглютиногены A и B.
Пример решения задачи №4
«Редкий рецессивный ген (h) в гомозиготном состоянии обладает эпистатическим действием по отношению к генам IA, IB и изменяет
их действие до I группы крови (бомбейский феномен). Определите возможные группы крови у детей, если у мужа II гомозиготная, у жены IV и оба родителя
гетерозиготны по эпистатическому гену»
Вероятность рождения детей с i(0) группой крови в данном случае равна 2/8, или 1/4 (25%). Генотипами, у которых будет i(0) группа крови являются:
IAIAhh и IAIBhh. Эпистатический рецессивный ген hh в гомозиготном состоянии всегда приводит к i(0)
группе крови.
Полимерия
Полимерией называют зависимость определенного признака организма от нескольких пар аллельных генов, обладающих схожим действием. Такие гены
называются полимерными. Часто выраженность признака зависит от соотношения доминантных и рецессивных аллелей — то есть чем больше доминантных генов,
тем более выражен признак.

У человека полимерное действие генов заложено в наследовании количественных признаков (вес, рост, цвет кожи, давление).
Пример решения задачи №5
«Цвет кожи у мулатов наследуется по типу полимерии. При этом данный признак контролируется 2 аутосомными несцепленными генами.
Сын белой женщины и негра женился на белой женщине. Может ли этот ребенок быть темнее своего отца?»
В данном случае полимерия проявляется в том, что чем больше доминантных генов в генотипе (A и B), тем более темный цвет кожи имеет человек. Это
правило мы и применим для решения.
В результате первого брака (вспоминаем закон единообразия Менделя) получается AaBb — средний мулат. По условиям задачи он берет в жены белую женщину
aabb. Очевидно, что в этой семье ребенок не может быть темнее своего отца: дети могут быть средними мулатами (AaBb), как отец, светлыми мулатами (aaBb, Aabb), либо белыми,
как мать (aabb).
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Взаимодействие генов
Проверка закономерностей независимого наследования на различных объектах уже в начале XX века показала, что, например, у ночной красавицы при скрещивании растений с красным и белым венчиком у гибридов первого поколения венчики окрашены в розовый цвет, тогда как во втором поколении имеются особи с красными, розовыми и белыми цветками в соотношении 1:2:1. Это навело исследователей на мысль, что аллельные гены могут оказывать определенное влияние друг на друга. Впоследствии было также установлено, что и неаллельные гены способствуют проявлению признаков других генов или подавляют их. Данные наблюдения стали основой представления о генотипе как о системе взаимодействующих генов. В настоящее время различают взаимодействие аллельных и неаллельных генов.
Ко взаимодействию аллельных генов относят полное и неполное доминирование, кодоминирование и сверхдоминирование. Полным доминированием считают все случаи взаимодействия аллельных генов, при которых в гетерозиготе наблюдается проявление исключительно доминантного признака, как, например, окраска и форма семени у гороха.
Неполное доминирование — это тип взаимодействия аллельных генов, при котором проявление рецессивного аллеля в большей или меньшей степени ослабляет проявление доминантного, как в случае окраски венчика ночной красавицы (белая + красная = розовая) и шерсти у крупного рогатого скота.
Кодоминированием называют такой тип взаимодействия аллельных генов, при котором оба аллеля проявляются, не ослабляя эффектов друг друга. Типичным примером кодоминирования является наследование групп крови по системе AB0.
Наследование групп крови по системе AB0
| Группа крови | Фенотип | Генотип | |
| агглютиногены | агглютинины | ||
| I (0) | — | $α$, $β$ | ii |
| II (A) | А | $β$ | IAIA или IAi |
| III (B) | В | $α$ | IBIB или IBi |
| IV (AB) | А и В | — | IAIB |
Как видно из таблицы, I, II и III группы крови наследуются по типу полного доминирования, тогда как IV (АВ) группа (генотип — IAIB) является случаем кодоминирования.
Сверхдоминирование — это явление, при котором в гетерозиготном состоянии доминантный признак проявляется намного сильнее, чем в гомозиготном; сверхдоминирование часто используется в селекции и считается причиной гетерозиса — явления гибридной силы.
Особым случаем взаимодействия аллельных генов можно считать так называемые летальные гены, которые в гомозиготном состоянии приводят к гибели организма чаще всего в эмбриональном периоде. Причиной гибели потомства является плейотропное действие генов серой окраски шерсти у каракулевых овец, платиновой окраски у лис и отсутствие чешуи у зеркальных карпов. При скрещивании двух гетерозиготных по этим генам особей расщепление по исследуемому признаку в потомстве будет равняться 2:1 вследствие гибели 1/4 потомства.
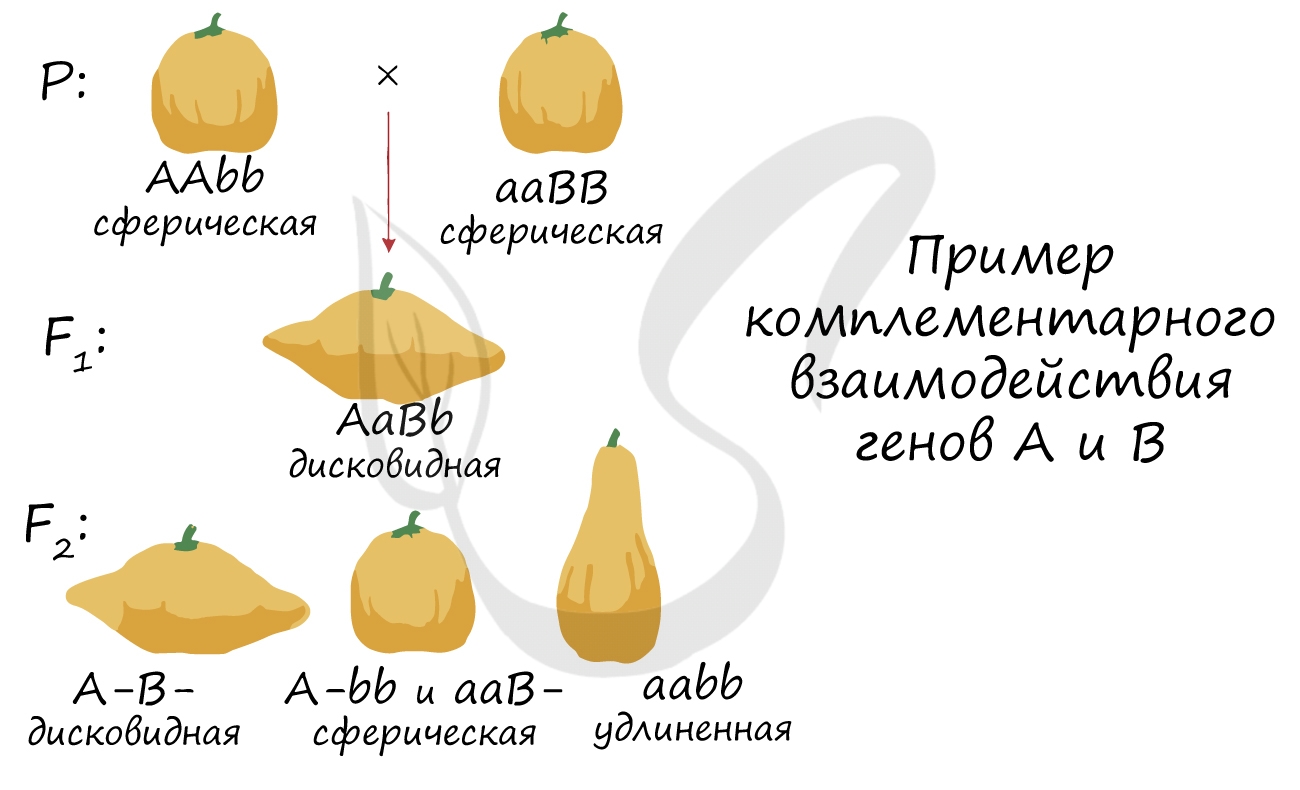
Основными типами взаимодействия неаллельных генов являются комплементарность, эпистаз и полимерия. Комплементарность — это тип взаимодействия неаллельных генов, при котором для проявления определенного состояния признака необходимо присутствие как минимум двух доминантных аллелей разных пар. Например, у тыквы при скрещивании растений со сферическими (ААbb) и длинными (aaBB) плодами в первом поколении появляются растения с дисковидными плодами (AaBb).
К эпистазу относят такие явления взаимодействия неаллельных генов, при которых один неаллельный ген подавляет развитие признака другого. Например, у кур окраска оперения определяется одним доминантным геном, тогда как другой доминантный ген подавляет развитие окраски, в результате чего большинство кур имеет белое оперение.
Полимерией называют явление, при котором неаллельные гены оказывают одинаковое влияние на развитие признака. Таким образом чаще всего кодируются количественные признаки. Например, цвет кожи человека определяется как минимум четырьмя парами неаллельных генов — чем больше доминантных аллелей в генотипе, тем темнее кожа.
Генотип как целостная система
Генотип не является механической суммой генов, поскольку возможность проявления гена и форма его проявления зависят от условий среды. В данном случае под средой понимается не только окружающая среда, но и генотипическая среда — другие гены.
Проявление качественных признаков редко зависит от условий окружающей среды, хотя, если у горностаевого кролика выбрить участок тела с белой шерстью и прикладывать к нему пузырь со льдом, то со временем на этом месте вырастет черная шерсть.
Развитие количественных признаков намного сильнее зависит от условий окружающей среды. Например, если современные сорта пшеницы возделывать без применения минеральных удобрений, то ее урожайность будет существенно отличаться от генетически запрограммированных 100 и более центнеров с гектара.
Таким образом, в генотипе записаны лишь «способности» организма, однако проявляются они только во взаимодействии с условиями окружающей среды.
Кроме того, гены взаимодействуют друг с другом и, оказавшись в одном генотипе, могут сильно влиять на проявление действия соседних генов. Таким образом, для каждого отдельно взятого гена существует генотипическая среда. Возможно, что развитие любого признака связано с действием многих генов. Кроме того, выявлена зависимость нескольких признаков от одного гена. Например, у овса окраска цветочных чешуй и длина их ости определяются одним геном. У дрозофилы ген белой окраски глаза одновременно влияет на цвет тела и внутренних органов, длину крыльев, снижение плодовитости и уменьшение продолжительности жизни. Не исключено, что каждый ген одновременно является геном основного действия для «своего» признака и модификатором для других признаков. Таким образом, фенотип — это результат взаимодействия генов всего генотипа с окружающей средой в онтогенезе особи.
В связи с этим известный российский генетик М. Е. Лобашев определил генотип как систему взаимодействующих генов. Сложилась эта целостная система в процессе эволюции органического мира, при этом выживали лишь те организмы, у которых взаимодействие генов давало наиболее благоприятную реакцию в онтогенезе.
Взаимодействие неаллельных генов егэ биология
Взаимодействие неаллельных генов
Неаллельные гены также могут взаимодействовать между собой. При этом их принцип взаимодействия несколько иной, чем доминантно-рецессивные отношения как в случае аллельных генов.
Правильнее говорить не о взаимодействии генов, а о взаимодействии их продуктов, т. е. взаимодействии белков, которые синтезируются на основе генов.
Комплементарное взаимодействие неаллельных генов — это такое их взаимодействие, при котором их продукты дополняют действие друг друга.
Примером комплиментарного взаимодействия генов является цвет глаз у мушки дрозофилы. У мушек с генотипом S-B — обычные красные глаза, ssbb — белые, S-bb — коричневые, ssB — — ярко-алые. Таким образом, если оба неаллельных гена рецессивны, то никакой пигмент не синтезируется, и глаза становятся белыми. При наличии только доминантного гена S появляется коричневый пигмент, а только доминантного B — ярко-алый. Если же есть два доминантных гена, то их продукты взаимодействуют между собой, образуя красный цвет.
При комплиментарном взаимодействии генов при скрещивании гетерозигот (AaBb) возможны разные расщепления по фенотипу (9:6:1, 9:3:3:1, 9:3:4, 9:7).
Эпистаз — это такое взаимодействие неаллельных генов, когда действие одного гена подавляет действие другого. Эпистатичным (подавляющим) действием на другой ген может обладать как доминантный, так и рецессивный аллель данного гена. Расщепление по фенотипу при доминантном эпистазе, отличается от рецессивного. Эпистатичный ген обычно обозначают буквой I.
Примером эпистаза может служить появление цветного оперения во втором поколении при скрещивании белых кур разных пород. У одних генотип IIAA, у других — iiaa. F1 — IaAa. В F2 происходит обычное расщепление по генотипу: 9I-A — : 3I-aa : 3iiA — : 1iicc. При этом птицы с генотипом iiA — оказываются окрашенными, что определяет доминантный ген A, который у одного родителя был подавлен доминантным геном-ингибитором I, а у другого присутствовал только в рецессивной форме.
При Полимерном взаимодействии неаллельных генов степень выраженности признака (его количество) зависит от количества доминантных аллельных и неаллельных генов. Чем больше генов участвуют в полимерном взаимодействии, тем больше различных степеней выраженности признака. Это происходит при комулятивной полимерии, когда все гены участвуют в накоплении признака. При некомулятивной полимерии количество доминантных генов не влияет на степень выраженности признака, достаточно хотя бы одного; а отличная по фенотипу форма наблюдается только у особей, у которых все полимерные гены рецессивны.
Полимерией, например, определяется цвет кожи человека. Влияние оказывают четыре гена (или четыре пары аллелей по другим источникам). Рассмотрим ситуацию с двумя парами. Тогда A1A1A2A2 определит самый темный цвет, a1a1a2a2 — самый светлый. Средний цвет кожи проявится, если два любых гена будут доминантны (A1a1A2a2 , A1A1a2a2, a1a1A2A2). Наличие одного доминантного гена приведет к цвету кожи близкому к светлому, но темнее, а трех доминантных — близкого к темному, но светлее.
Бывает, что один ген определяет несколько признаков. Такое действие гена называется Плейотропией. Понятно, что здесь речь идет не о взаимодействии генов, а с множественным действием одного гена.
«Дигетерозиготная по B (III) группе и положительному резус-фактору вступила в брак с таким же мужчиной. Какое расщепление по фенотипу можно ожидать у детей?»
Два средних мулата имеют двух детей-близнецов: черного и белого ребенка. Можно ли установить генотипы родителей?
Пример решения задачи №1
Составим схему решения для такого случая.
Biology. su
14.08.2017 16:35:22
2017-08-14 16:35:22
Взаимодействие генов
Вы уже знаете о том, что гены могут взаимодействовать друг с другом по типу полного и неполного доминирования. Однако, в генетике встречается масса других примеров взаимодействия генов. В этой статье мы затронем те, которые ранее не обсуждались.
Кодоминирование
Кодоминирование — взаимодействия аллельных генов, при котором в гетерозиготном состоянии могут оказаться два доминантных гена одновременно, при этом каждый ген отвечает за свой признак.
Наиболее распространенным примером кодоминирования является наследование групп крови у человека.
Решим пару задач, которые укрепят понимание темы.
Пример решения задачи №1
«Родители имеют II и III группы крови, гетерозиготны. Какие группы крови можно ожидать у их детей?»
Гетерозиготный генотип матери — I A i 0 и генотип отца — I B i 0 . Составим схему решения для такого случая.
Итак, в результате такого брака может получиться ребенок с любой группой крови, в чем мы убедились.
Пример решения задачи №2
«Дигетерозиготная по B (III) группе и положительному резус-фактору вступила в брак с таким же мужчиной. Какое расщепление по фенотипу можно ожидать у детей?»
Сходу понятно, что гетерозиготы по III (B) группе крови будут записаны I B i 0 . Резус-фактор для нас новое понятие — это белок, находящийся на поверхности эритроцита (тогда говорят, что резус-фактор положителен), или отсутствующий (тогда у человека резус-фактор считается отрицательным). Генотипы записываются так:
- Резус-фактор положителен: Rh + Rh + , Rh + rh — Резус-фактор отрицателен: rh — rh —
В данной задаче сказано, что «дигетерозиготна по. и положительному резус-фактору» — значит, резус фактор будет записывать в генотипе — Rh + rh — .
Обратите внимание, что ошибкой является записать рецессивный ген перед доминантным. За такое могут снять балл на экзамене: aA, bB, i 0 I A, rh — Rh + . Правильный вариант записи: Aa, Bb, I A i 0 , Rh + rh — .
- 9 потомков : положительный резус-фактор, III (B) группа крови 3 потомка : положительный резус-фактор, I (0) группа крови 3 потомка : отрицательный резус-фактор, III (B) группа крови 1 потомок : отрицательный резус-фактор, I (0) группа крови
Расщепление по фенотипу в данном случае получилось: 9:3:3:1. Здесь проявляется III закон Менделя — закон независимого наследования, так как гены, отвечающие за группу крови и резус-фактор, находятся в разных хромосомах.
Комплементарность
Комплементарность — тип взаимодействия неаллельных генов, при котором развитие признака определяется не одной, а двумя или более парами неаллельных генов, располагающихся в разных хромосомах.
Неаллельные гены — это гены, расположенные в разных локусах хромосом, которые отвечают за разные признаки. В генетике случается такое, что один неаллельный ген может влиять на другой (ген a подавляет действие гена B). В этом разделе статьи мы подробно разберемся с подобным взаимодействием и рассмотрим задачи, которые могут встретиться.
Таким образом, развитие признака определяется именно сочетанием генов друг с другом. Здесь логичнее подчеркнуть совместное действие генов, нежели чем сказать, что доминантный ген подавляет рецессивный — при комплементарности это не совсем так.
В каждой задаче свой случай комплементарного взаимодействия генов. Чтобы успешно их решать, надо помнить, что такое явление, как комплементарность, в принципе, возможно, и быть внимательным при написании генотипов особей и их гамет.
Пример решения задачи №3
Наследование слуха у человека определяется двумя доминантными генами из разных аллельных пар, один из которых детерминирует развитие слухового нерва, а другой – улитки. Определить вероятность рождения глухих детей, если оба родителя глухие, но по разным генетическим причинам (у одного отсутствует слуховой нерв, у другого улитка). По генотипу оба родителя являются дигомозиготными.
Здесь проявляется I закон Менделя — закон единообразия гибридов первого поколения. Возможен только один вариант генотипа ребенка от такого брака. У ребенка будет развит и слуховой нерв, и улитка — ребенок не будет глухим, в отличие от родителей.
Эпистаз
Эпистаз (противоположное действие генов) — явление, при котором один ген аллельной пары (супрессор) в доминантном (доминантный эпистаз) или рецессивном (рецессивный эпистаз) состоянии может подавлять развитие признака, за развитие которого отвечает другая пара генов.
Широко известным примером рецессивного эпистаза является Бомбейский феномен, названный так в результате зафиксированного случая в индийском городе Бомбеи. Доктор Бхенде обнаружил, что у людей рецессивных по гену h (hh) на поверхности эритроцитов не синтезируются агглютиногены — в результате этого они могут быть универсальными донорами.
Говоря проще о Бомбейском феномене: у людей с генотипом hh всегда обнаруживается первая группа крови при любом генотипе — I A I A, I B I B, I A I B. Ген h подавляет гены I A и I B — на поверхности эритроцитов не образуются агглютиногены A и B.
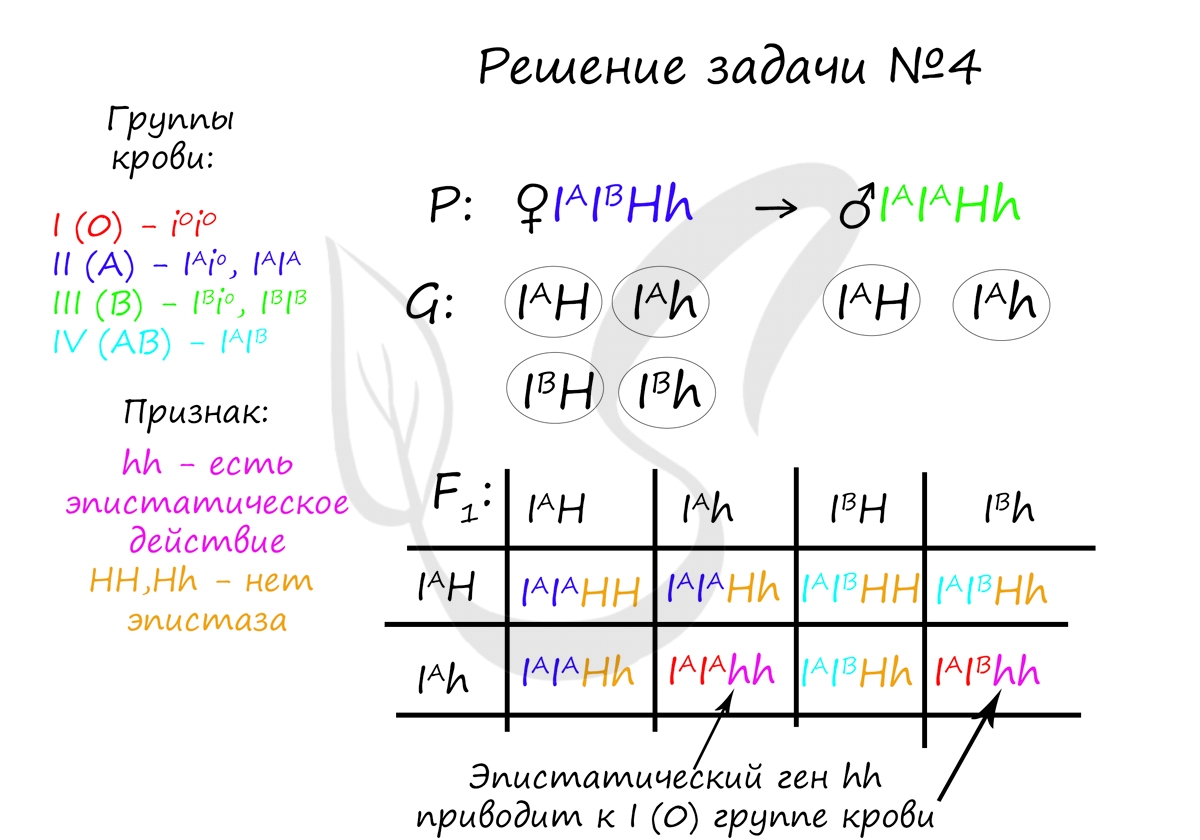
Пример решения задачи №4
«Редкий рецессивный ген (h) в гомозиготном состоянии обладает эпистатическим действием по отношению к генам I A, I B и изменяет их действие до I группы крови (бомбейский феномен). Определите возможные группы крови у детей, если у мужа II гомозиготная, у жены IV и оба родителя гетерозиготны по эпистатическому гену»
Вероятность рождения детей с i(0) группой крови в данном случае равна 2/8, или 1/4 (25%). Генотипами, у которых будет i(0) группа крови являются: I A I A hh и I A I B hh. Эпистатический рецессивный ген hh в гомозиготном состоянии всегда приводит к i(0) группе крови.
Полимерия
Полимерией называют зависимость определенного признака организма от нескольких пар аллельных генов, обладающих схожим действием. Такие гены называются полимерными. Часто выраженность признака зависит от соотношения доминантных и рецессивных аллелей — то есть чем больше доминантных генов, тем более выражен признак.
У человека полимерное действие генов заложено в наследовании количественных признаков (вес, рост, цвет кожи, давление).
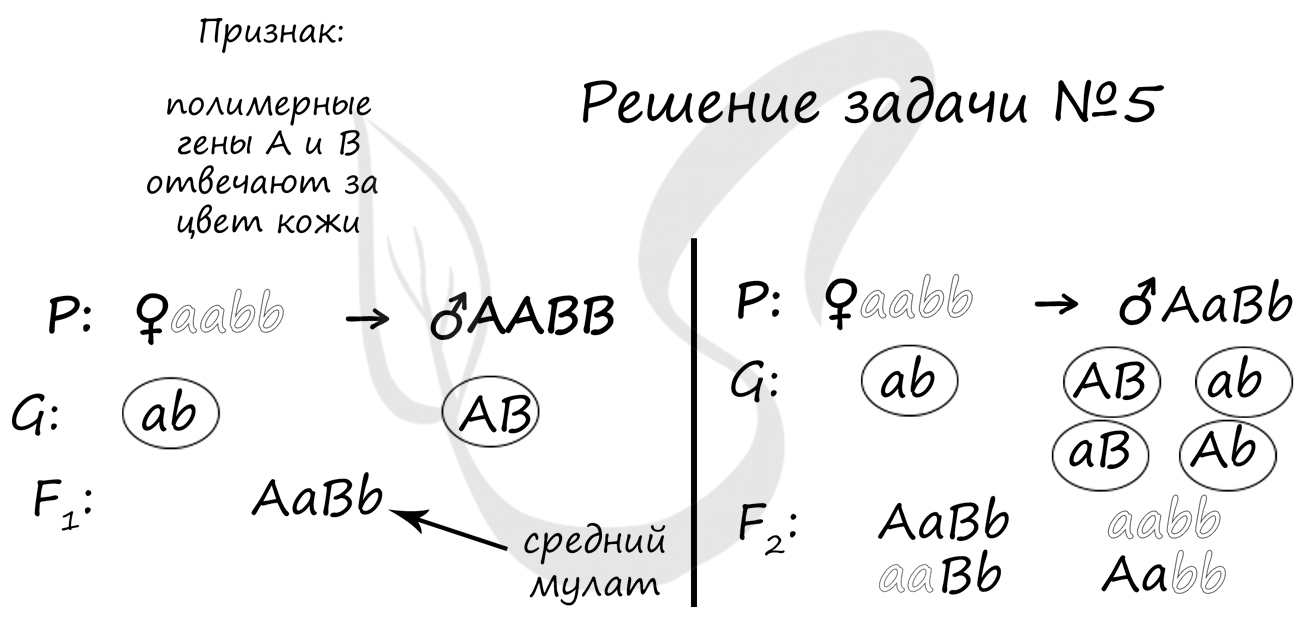
Пример решения задачи №5
«Цвет кожи у мулатов наследуется по типу полимерии. При этом данный признак контролируется 2 аутосомными несцепленными генами. Сын белой женщины и негра женился на белой женщине. Может ли этот ребенок быть темнее своего отца?»
В данном случае полимерия проявляется в том, что чем больше доминантных генов в генотипе (A и B), тем более темный цвет кожи имеет человек. Это правило мы и применим для решения.
В результате первого брака (вспоминаем закон единообразия Менделя) получается AaBb — средний мулат. По условиям задачи он берет в жены белую женщину aabb. Очевидно, что в этой семье ребенок не может быть темнее своего отца: дети могут быть средними мулатами (AaBb), как отец, светлыми мулатами (aaBb, Aabb), либо белыми, как мать (aabb).
© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
При скрещивании растений одного из сортов тыквы с белыми и желтыми плодами все потомство F 1 имело белые плоды. При скрещивании этого потомства между собой в их потомстве F 2 было получено:
204 растения с белыми плодами,
53 растения с желтыми плодами,
17 растений с зелеными плодами.
Определить возможные генотипы родителей и потомства.
AB Ab aB ab
Сын белой женщины и негра женится на белой женщине.
Studarium. ru
23.05.2018 3:32:35
2018-05-23 03:32:35
Консультация к ЕГЭ «Взаимодействие неаллельных генов»
Материал для подготовки к егэ (гиа) по биологии (11 класс)
Проявление одного признака может определяться двумя и более парами генов ( комплементарность и полимерное наследование ) и, наоборот, одна пара генов может влиять на проявление нескольких признаков ( множественное действие генов ). Кроме того, одни гены могут подавлять действие других ( эпистаз ). Все эти явления получили общее название взаимодействие генов.
При взаимодействии двух пар генов вероятность появления организмов каждого фенотипа определяется различными сочетаниями двух пар генов, участвующих в развитии признака. Так же, как и в случае дигибридного скрещивания, эти сочетания можно оценивать с помощью решетки Пеннета. В большинстве случаев различные соотношения фенотипов при скрещивании дигетерозигот образованы из соотношения 9:3:3:1 путем сложения отдельных элементов, например, расщепление 9:7 при комплементарном наследовании может быть представлено как 9:(3+3+1).
При решении задач такого типа следует обращать внимание на то, сколько классов фенотипов и в каком количественном соотношении образуется при скрещивании особей с различными генотипами.
2. Полимерное действие генов
Ответы и решения
Развитие признака может определяться не одной, а двумя или более парами неаллельных генов, располагающимися в разных хромосомах. Если хотя бы одна пара находится в гомозиготном рецессивном состоянии, то признак не развивается или отличен от доминантного.
С биохимической точки зрения зачастую это может быть связано с тем, что развитие признаков обычно представляет собой многостадийный процесс, каждый этап которого контролируется отдельным ферментом (информация о ферменте находится в определенном гене). Если хотя бы один ген находится в рецессивном состоянии, то синтезируется измененный фермент, реакция не идет, и конечный продукт не образуется:
Расщепление при скрещивании дигетерозигот при комплементарном наследовании обычно бывает в пропорции 9:7, 9:3:4, или 9:3:3:1, 9:6:1 (часть особей с минимальным выражением признака 7/16, 4/16 и 1/16).
У душистого горошка окраска цветов проявляется только при наличии двух доминантных генов А и В. Если в генотипе имеется только один доминантный ген, то окраска не развивается. Какое потомство F 1 и F 2 получится от скрещивания растений с генотипами ААbb и ааВВ?
Генотип исходных растений известен по условию задачи. Они гомозиготны и будут давать один тип гамет, которые можно объединить единственным образом. Потомство F 1 будет единообразно по генотипу (АаВb) и фенотипу (розовые цветы):

Ab
aB
Дигетерозиготные потомки F 1 будут давать по 4 типа гамет, которые могут комбинироваться 16 способами. Построив решетку Пеннета, можно убедиться, что организмов, одновременно имеющих доминантные гены А и В (розовые цветы), будет примерно 9/16. То есть, в F 2 будет наблюдаться расщепление по цвету в пропорции 9:7.
AB Ab aB ab
AB Ab aB ab
A*bb aaB* aabb
Белый
7/16
В F 1 все потомство будет единообразным и будет иметь розовую окраску цветов. В F 2 9/16 растений будут иметь розовые, а 7/16 – белые цветы.
Окраска цветов душистого горошка в красный цвет обусловлена двумя парами генов. Если хотя бы одна пара находится в рецессивном состоянии, то окраска не развивается. Одновременное присутствие в генотипе обоих доминантных генов вызывает развитие окраски. Каков генотип растений с белыми цветами, если при их скрещивании друг с другом все растения получились красного цвета?
Поскольку потомство F 1 имело окрашенные цветки, то в его генотипе присутствуют оба доминантных гена А и В. Все потомство единообразно, следовательно, скрещивались гомозиготные особи. Родители имели неокрашенные цветки, значит, они не могут нести оба доминантных гена. Генотип родителей не может быть одинаковым (иначе потомство было бы таким же, то есть имело бы неокрашенные цветки). Этим условиям удовлетворяет только тот вариант, когда у одного родителя генотип ААbb, а у другого – ааВВ.
Ab
aB aB
При скрещивании двух растений тыквы со сферической формой плодов получено потомство, имеющее только дисковидные плоды. При скрещивании этих гибридов между собой были получены растения с тремя типами плодов:
9 частей – с дисковидными плодами,
6 частей – со сферической формой плодов,
1 часть – с удлиненными плодами.
Какая закономерность наблюдается в данном случае? Каковы генотипы родителей и потомства?
Потомство F 1 отличалось от родителей и было единообразным, следовательно, оно является гетерозиготным, а исходные особи были гомозиготными. Исходные особи должны отличаться по генотипу друг от друга, так как при одинаковом генотипе их потомство не отличалось бы от них. Это указывает на то, что в данном случае имеет место комплементарное взаимодействие генов. Генотип родительских особей – ААbb и ааВВ, а потомства F 1 – АаВb (см. решение задачи 5-2). Таким образом, форма плодов определяется двумя парами генов. Если обе пары находятся в доминантном состоянии (генотипы ААВВ, АаВb, ААВb или АаВВ), то форма плодов дисковидная. Если в доминантном состоянии находится только одна пара генов (генотипы ААbb, Ааbb, ааВВ или ааВb), – форма плодов сферическая. Расщепление в потомстве F 2 (9:6:1) отличается от расщепления 9:7, обычно наблюдаемого при комплементарном взаимодействии. 1/16 часть растений имела плоды удлиненной формы. Это может быть только в том случае, если генотип таких особей – ааbb, то есть оба гена находятся в рецессивном состоянии. Анализ решетки Пеннета подтверждает полученные результаты.
Среди ферментов, участвующих в образовании хлорофилла у ячменя, имеется два фермента, отсутствие которых приводит к нарушению синтеза этого пигмента. Если нет одного из них, то растение становится белым, если нет другого – желтым. При отсутствии обоих ферментов растение также белое. Синтез каждого фермента контролируется доминантным геном. Гены находятся в разных хромосомах. Какое потомство следует ожидать при самоопылении гетерозиготного по обоим генам ячменя?
У кукурузы нормальный рост определяется двумя доминантными неаллельными генами. Гомозиготность по рецессивным аллелям даже одной пары генов приводит к возникновению карликовых форм. При скрещивании двух карликовых растений кукурузы выросли гибриды нормальной высоты, а при скрещивании этих гибридов в их потомстве было получено 812 нормальных и 640 карликовых растений. Определить генотипы родителей и потомков.
У норки известно два рецессивных гена – р и i, гомозиготность по каждому из которых, или по обоим одновременно, обуславливает платиновую окраску меха. Дикая коричневая окраска получается при наличии обоих доминантных аллелей Р и I. При каком типе скрещивания двух платиновых норок все их потомство будет коричневым?
При скрещивании двух карликовых растений кукурузы получено потомство нормальной высоты. В F 2 от скрещивания потомства первого поколения было 452 растения нормальной высоты и 352 – карликовых. Предложите гипотезу, объясняющую эти результаты.
В двух цветоводческих хозяйствах, не связанных друг с другом, длительное время разводили чистые линии душистого горошка с белыми цветами. Какое потомство можно ожидать от скрещивания этих двух чистых линий?
Окраска шерсти у кроликов определяется двумя парами генов, расположенных в разных хромосомах. При наличии доминантного гена С доминантный ген А другой пары обуславливает серую окраску шерсти, рецессивный ген а – черную окраску. В отсутствии гена С окраска будет белая. Крольчата какого цвета получатся от скрещивания серых дигетерозиготных кроликов?
Окраска бобов может быть пурпурной, желтой и белой. Под действием гена А неокрашенное соединение переводится в пурпурный пигмент. Ген В вызывает превращение пурпурного вещества в желтое. Какое потомство получится от скрещивания растений с генотипами АаВb и ааВВ?
Для получения окрашенных луковиц необходимо наличие у растений лука доминантного гена С. При гомозиготности по рецессивному аллелю с получаются бесцветные луковицы. При наличии доминантного гена С вторая пара аллелей определяет цвет луковицы – красный (R) или желтый (r). Краснолуковичное растение было скрещено с белолуковичным. В потомстве были растения с красными, желтыми и бесцветными луковицами. Определить генотипы скрещиваемых растений. Какое расщепление по фенотипу произошло в потомстве? Какое расщепление было бы в потомстве, если бы обе исходные особи были красного цвета?
У лука ген R определяет красную окраску чешуй, а ген r – желтую. Любая окраска проявляется только при наличии в генотипе доминантного гена С, при его отсутствии чешуи имеют белую окраску. Определить генотипы исходных форм луковиц с белыми и красными чешуями, если все гибридные луковицы имели красную окраску чешуй.
От скрещивания белых и серых мышей в потомстве F 1 все особи были черными, а в F 2 было 77 черных, 37 серых и 45 белых мышей. Как наследуется окраска у этих мышей? Определить генотипы родителей и потомков.
Собаки породы кокер-спаниель при генотипе А*В* имеют черную масть, при генотипе А*bb – рыжую, при генотипе ааВ* – коричневую, а при генотипе ааbb – светло-желтую. При скрещивании черного кокер-спаниеля со светло-желтым родился светло-желтый щенок. Какое соотношение по масти следует ожидать от спаривания того же черного спаниеля с собакой одинакового с ним генотипа?
2. Полимерное действие генов
При полимерном наследовании развитие одного признака контролируется несколькими парами генов, расположенных в разных хромосомах. Чем больше генов находится в доминантном состоянии, тем ярче выражен признак.
Полимерное действие лежит в основе наследования количественных признаков и играет важную роль в эволюции.
Например, степень пигментации кожи определяется двумя парами (на самом деле – большим количеством) генов. В соответствии с этим по данному признаку людей можно условно разделить на 5 фенотипов: негры (ААВВ), темные мулаты (ААВb или АаВВ), средние мулаты (АаВb, ааВВ или ААbb), светлые мулаты (Ааbb или ааВb) и белые (ааbb).
Сын белой женщины и негра женится на белой женщине. Может ли ребенок от этого брака быть темнее своего отца?
Какой фенотип потомства будет:
А) от брака негра и светлой мулатки;
Б) от брака белого и темной мулатки?
Какое потомство получится от брака:
А) двух средних гетерозиготных мулатов;
Б) двух средних гомозиготных мулатов?
Два средних мулата имеют двух детей-близнецов: черного и белого ребенка. Можно ли установить генотипы родителей?
От брака среднего мулата и светлой мулатки родилось много детей, среди которых оказалось по 3/8 средних и светлых мулатов и по 1/8 – темных мулатов и белых. Каковы возможные генотипы родителей?
Может ли у одной пары родителей родиться двое детей-близнецов, один из которых белый, а другой – негр?
Эпистазом, или противоположным действием генов, называется явление, при котором ген одной аллельной пары (супрессор) в доминантном состоянии может подавлять развитие признака, контролируемого другой парой генов.
В случае эпистаза при скрещивании дигетерозигот в потомстве наблюдается расщепление в соотношении 13:3 или 12:3:1.
При скрещивании растений одного из сортов тыквы с белыми и желтыми плодами все потомство F 1 имело белые плоды. При скрещивании этого потомства между собой в их потомстве F 2 было получено:
204 растения с белыми плодами,
53 растения с желтыми плодами,
17 растений с зелеными плодами.
Определить возможные генотипы родителей и потомства.
Потомство F 1 единообразно. Это указывает на то, что родители были гомозиготны, и признак белой окраски доминирует. Гибриды первого поколения F 1 гетерозиготны (получены от родителей с разным генотипом и имеют расщепление в F 2 ). Во втором поколении имеется три класса фенотипов, но расщепление отличается от расщепления при кодоминировании (1:2:1) или при комплементарном наследовании (9:6:1, 9:3:4, 9:7 или 9:3:3:1). Предположим, что признак определяется противоположным действием двух пар генов, причем особи, у которых обе пары генов находятся в рецессивном состоянии (ccjj), отличаются по фенотипу от особей, у которых действие гена не подавляется. Расщепление в потомстве 12:3:1 подтверждает это предположение.
Генотипы родителей – ССjj и ссJJ, потомства F 1 – СсJj.
У кур породы леггорн окраска перьев обусловлена наличием доминантного гена С. Если он находится в рецессивном состоянии, то окраска не развивается. На действие этого гена оказывает влияние ген I, который в доминантном состоянии подавляет развитие признака, контролируемого геном С. Какое потомство получится от скрещивания дигетерозиготных по этим генам кур породы леггорн?
У кур породы леггорн окраска перьев обусловлена наличием доминантного гена С. Если он находится в рецессивном состоянии, то окраска не развивается. На действие этого гена оказывает влияние ген I, который в доминантном состоянии подавляет развитие признака, контролируемого геном С. Определить вероятность рождения окрашенного цыпленка от скрещивания кур с генотипом ССIi и ссIi.
При скрещивании чистых линий собак коричневой и белой масти все потомство имело белую окраску. Среди потомства полученных гибридов было 118 белых, 32 черных и 10 коричневых собак. Предложите гипотезу, объясняющую эти результаты.
У лошадей действие генов вороной (С) и рыжей масти (с) проявляется только в отсутствие доминантного гена D. Если он присутствует, то окраска белая. Какое потомство получится при скрещивании между собой белых лошадей с генотипом CcDd?
У овса черная окраска семян определяется доминантным геном А, а серая окраска – доминантным геном В. Ген А эпистатичен гену В, и последний в его присутствии не проявляется. При отсутствии в генотипе обоих доминантных генов окраска семян белая. При скрещивании двух серосеменных растений получили растения с серыми и белыми семенами в пропорции 3:1. Определить генотипы родителей. Какое расщепление в потомстве было бы получено, если бы скрещивались дигетерозиготные формы?
Свиньи бывают черной, белой и красной окраски. Белые свиньи несут минимум один доминантный ген I. Черные свиньи имеют доминантный ген Е и гомозиготны по рецессивной аллели i. Красные поросята (eeii) лишены доминантного гена-подавителя I и доминантного гена, определяющего черную окраску. Какое потомство можно ожидать от скрещивания черной гомозиготной свиньи и красного кабана?
Ответы и решения к задачам главы V
5-1. В F 1 все потомство будет единообразно и будет иметь розовую окраску цветов. В F 2 9/16 растений будут иметь розовые, 7/16 – белые цветы.
5-2. Генотип исходных растений – ААbb и ааВВ.
5-3. Объяснение в тексте.
5-4. 9/16 – зеленые (АаВb, ААВВ, ААВb и АаВВ),
3/16 – желтые (ААbb и Ааbb),
4/16 – белые (ааВВ, ааВb и ааbb).
5-5. Исходные организмы имели генотипы ААbb и ааВВ. Единообразное потомство F 1 несло генотип АаВb; в потомстве F 2 наблюдалось расщепление 9:7, характерное для комплементарного наследования.
5-6. Для скрещивания необходимо взять животных с генотипами ppII и PPii.
5-7. Наблюдаемое в потомстве F 2 расщепление в пропорции 9:7 может быть при комплементарном наследовании, когда развитие признака (нормальный рост) определяется двумя парами генов в доминантном состоянии. Для проверки этого предположения надо провести возвратное скрещивание потомка F 1 c одним из родителей. В этом случае должно быть расщепление по фенотипу в соотношении 1:1.
5-8. Возможные варианты: ААbb × ааВВ – окрашенные, ААbb × ААbb – белые, ааВВ × ааВВ – белые.
5-9. Получатся серые, черные и белые крольчата в соотношении 9:3:4.
5-10. Получится следующее потомство: 25% – АаВВ – пурпурные, 25% – АаВb – пурпурные, 25% – ааВВ – белые, 25% – ааВb – белые. Расщепление по фенотипу равно 1:1.
5-11. Генотипы исходных особей – CcRr и Ccrr. В потомстве от их скрещивания наблюдается расщепление по фенотипу в пропорции 3:3:2. Если бы обе исходные особи были красного цвета (генотип – CcRr), то было бы расщепление в соотношении 9:3:4.
5-12. Генотип исходных растений с белыми цветами – ссrr, с красными цветами – CCRR.
5-13. По-видимому, признак кодируется двумя парами комплементарных генов, которые у родителей находятся в разных состояниях (ААbb и ааВВ). Соотношения фенотипов потомства подтверждает это предположение:
А*В* (черных) – 9/16,
А*bb (серых) – 3/16,
АаВ* и ааbb (белых) – 4/16.
5-14. В потомстве следует ожидать следующее расщепление по масти:
А*В* (черная) – 9/16,
А*bb (рыжая) – 3/16,
АаВ* (коричневая) – 3/16,
Ааbb (светло-желтая) – 1/16.
5-15. Задача имеет три варианта решения. Во всех вариантах ребенок содержит меньше доминантных генов, чем отец, и, следовательно, будет светлее его.
5-16. а) От брака негра и светлой мулатки в равном соотношении будут рождаться темные и средние мулаты.
Б) От брака белого и темной мулатки с равной вероятностью будут рождаться светлые или средние мулаты.
5-17. а) От брака двух средних гетерозиготных мулатов будут рождаться негры (1/16), темные мулаты (4/16), средние мулаты (6/16), светлые мулаты (4/16) и белые (1/16).
Б) От брака средних гомозиготных мулатов, независимо от их генотипа, родятся только средние мулаты.
5-18. АаВb и АаВb. Только в этом случае в их потомстве будут одновременно встречаться дети с генотипами ааbb и ААВВ.
5-19. Генотип отца – АаВb, генотип матери – Ааbb или ааВb.
5-20. Может, если близнецы разнояйцовые, а родители являются дигетерозиготными мулатами (генотип АаВb). Тогда у них могут быть потомки с генотипом ААВВ (негр) и ааbb (белый).
5-21. Генотипы родителей – ССjj и ссJJ, потомства F 1 – СсJj.
5-22. В потомстве будет 3/16 окрашенных цыплят (С*ii) и 13/16 белых (генотипы **I* и ссii).
5-23. Вероятность рождения окрашенного цыпленка (Ccii) – 1/4.
5-24. Решение аналогично решению задачи 5-21.
5-25. СCDD, CcDD, CCDd, CcDd, ccDD и ccDd – белые (12/16), СCdd и Ccdd – вороные (3/16), ссdd – рыжие (1/16).
5-26. ааВb и ааВb. При скрещивании дигетерозигот было бы получено соотношение черных, серых и белых семян в пропорции 12:3:1.
5-27. Красный кабан по условию задачи имеет генотип eeii. Генотип свиньи не может быть ЕЕII, так как в этом случае она не была бы окрашенной. Следовательно, ее генотип ЕЕii. Каждое из животных дает один тип гамет (ei и Ei соответственно), которые можно объединить единственным образом. В результате этого получатся черные поросята с генотипом Eeii.
Вы уже знаете о том, что гены могут взаимодействовать друг с другом по типу полного и неполного доминирования. Однако, в генетике встречается масса других примеров взаимодействия генов. В этой статье мы затронем те, которые ранее не обсуждались.
«Редкий рецессивный ген (h) в гомозиготном состоянии обладает эпистатическим действием по отношению к генам I A, I B и изменяет их действие до I группы крови (бомбейский феномен). Определите возможные группы крови у детей, если у мужа II гомозиготная, у жены IV и оба родителя гетерозиготны по эпистатическому гену»
Тогда A 1 A 1 A 2 A 2 определит самый темный цвет, a 1 a 1 a 2 a 2 самый светлый.
Nsportal. ru
17.01.2018 9:32:43
2018-01-17 09:32:43
Источники:
Http://biology. su/genetics/non-allele
Http://studarium. ru/article/128
Http://nsportal. ru/shkola/biologiya/library/2020/11/22/konsultatsiya-k-ege-vzaimodeystvie-neallelnyh-genov
Взаимодействия неаллельных генов❗. | Биология ЕГЭ-2022 | ВКонтакте » /> » /> .keyword { color: red; } Взаимодействие неаллельных генов егэ биология
Взаимодействие неаллельных генов егэ биология
Взаимодействие неаллельных генов егэ биология

Биология ЕГЭ-2022 запись закреплена
Взаимодействия неаллельных генов
#биология_ЕГЭ #общая_биология #генетика
Неаллельные гены — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки.
1. Комплементарное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое
Проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9 : 6 : 1, 9 : 3 : 4, 9 : 7, иногда 9 : 3 : 3 : 1.
2. Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным.
Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I.
Эпистатическое взаимодействие неаллельных генов
Может быть доминантным и рецессивным.
3. Полимерия — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным.
14. От скрещивания растений льна с белыми и голубыми лепестками цветков получены растения с голубыми лепестками. В следующем поколении идет расщепление в отношении 33 голубых : 12 розовых : 18 белых. Какое явление иллюстрирует данный случай и каковы генотипы всех форм?
20 При скрещивании двух белозерных растений кукурузы F1 получилось белозерное, а в F2 получено 138 белых семян и 39 пурпурных. К какому типу наследования относится этот случай? Определите генотипы всех форм.
Теоретический вопрос ДНЯ
Во втором поколении на каждые 16 лошадей в среднем появилось 12 серых, 3 вороных,1 рыжая.
Vk. com
06.01.2020 0:24:28
2020-01-06 00:24:28
Задачи по генетике на взаимодействие неаллельных генов

Когда я заканчивал писать книгу «Как научиться решать задачи по генетике», то в заключении говорил, что прощаюсь с вами не надолго. Откуда у меня была такая уверенность?
Да всё просто. Даже знание самих азов генетики — этой завораживающей науки о наследственности, уже делает человека другим. Мудрее, что ли…. В отношении к жизни вообще, в отношении понимания своей собственной жизни.
А тема взаимодействия неаллельных генов бесспорно важна. Но в учебниках по биологии и генетике она преподносится настолько «сухарным» языком, что для многим остается совершенно непонятой.
В теоретической части этого сборника я постарался так изложить данный материал, чтобы было понятно «как и с чем его едят»:
Во-первых, надо понять, что общего в заданиях на взаимодействие неаллельных генов с моногибридным скрещиванием: и там, и там изучается наследование одного признака,
Во-вторых, что общего в заданиях на взаимодействие неаллельных генов с дигибридным скрещиванием: за призн ак (при взаимодействии неаллельных генов) и за призна ки (при дигибридном скрещивании) отвечают разные неаллельные гены.
Кому может быть полезен этот сборник:
1. Абитуриентам. В школьных учебниках по общей биологии даже базового уровня тема взаимодействия неаллельных генов рассматривается. Поэтому в любой год можно ожидать вопросов и задач этой тематики в контрольных заданиях ЕГЭ.
2. Студентам медицинских и биологических факультетов, изучающим общую генетику на первом курсе. Именно решения задач на взаимодействие неаллельных генов встречают у многих наибольшие затруднения.
3. Учителям и репетиторам по биологии. Здесь особый разговор. Можно самому быть, что называется «в теме», но часто бывает довольно сложным преподнести учащимся данный непростой материал в наиболее доступной форме.
В практической части этого сборника, я постарался подобрать такие задания, что бы мое разъяснение их решений могло помочь вам в решении любых подобных заданий.
Итак, предлагаю Вашему вниманию платную электронную книгу:
«Задачи по генетике на взаимодействие неаллельных генов»
* Ознакомившись с небольшой теоретической частью книги, Вам эта тема не покажется сложной
* Решения 47 задач по комплементарному, эпистатическому и полимерному взаимодействию генов даются с полным разъяснением.
Книга в формате. pdf. Стоимость 400 рублей.
Напишите мне письмо boris. sadykov@gmail. com о желании сделать покупку.
После моего ответа, что Вы приняли мудрое решение, Вы будете знать, что у меня есть Ваш адрес электронной почты, на который я в тот же день (иногда в тот же час) после оплаты вышлю купленные материалы.
ОПЛАТИТЬ МОЖНО:
1. На мою Электронную карту Сбербанка России : 2202 2008 1307 8159
2. электронные кошельки:

Яндекс Деньги 41001493555269
3. На ТЕЛЕФОН (оператор МЕГАФОН — Поволжский филиал): +7(927)32-32-052
Если возникнут вопросы по скачиванию и изучению материалов на вашем компьютере, обязательно напишите — без поддержки я Вас не оставлю.
Задачи, решения которых подробно разбираются в этом сборнике
Комплементарное взаимодействие генов
1. Нормальный темно-красный цвет глаз дрозофилы определяется двумя пигментами — красным и коричневым. Рецессивный аллель bw нарушает синтез красного пигмента и глаза у гомозигот коричневые. Другой рецессивный аллель st дает ярко-красный цвет глаз, так как отсутствует коричневый пигмент. У двойных гомозигот глаза белые, так как отсутствуют оба пигмента. Скрестили гомозиготных мух с коричневыми глазами и ярко-красными глазами. Какие потомки и в каком соотношении получатся в F1 и F2?
2. У охотничьих собак коккер-спаниель черная окраска шерсти (А) доминирует над бурой (а), сплошная окраска (S) — над пятнистой (s), гены не сцеплены с полом. От скрещивания собак, различающихся по окраске, в потомстве получено 3/8 черных, 3/8 бурых, 1/8 черно-белых, 1/8 буро-белых. Определите генотипы и фенотипы родителей.
3. У попугаев аллель А — желтый цвет перьев, а аллель В — голубой. При взаимодействии генов АВ — зеленый. Особи с генотипом ааbb — белые. При скрещивании гетерозиготных особей с желтыми и голубыми перьями, получено 20 попугаев. Сколько среди них белых?
4. У попугайчиков-неразлучников окраска оперения определяется двумя парами генов а) при спаривании между собой зелёных попугайчиков было получено 55 зелёных, 18 жёлтых, 17 голубых и 6 белых потомков. Определите генотипы родителей и потомков. Как наследуется окраска? б) спаривания зелёных и голубых попугайчиков не дало белых потомков. Определите генотипы родителей.
5. У кукурузы окраска алейрона у семян определяется комплементарным взаимодействием генов А и В, которые в доминантном состоянии обуславливают развитие в зерновке окрашенного алейронового слоя, а в рецессивном — неокрашенного. При скрещивании двух видов кукурузы, имеющих неокрашенный алейрон, в F1 было получено 24 растения с окрашенным алейроном, в F2 160 растений. Сколько растений в F2 имели окрашенный алейрон? Сколько разных фенотипов будет в F2? Сколько разных генотипов будет в F2? Сколько гомозиготных растений будут иметь неокрашенный алейроновый слой? Сколько гомозиготных растений будут иметь окрашенный алейроновый слой в зерновке?
6. Белая курица породы плимутрок скрещивалась со светлым петухом породы брама. В F1 все потомство имело черное оперение. При скрещивании гибридов F1 между собой получено 50 черных, 13 светлых и 23 белых птицы (всего 86). При скрещивании гибридов F1 с плимутроками получено 29 черных и 22 белых птицы (всего 51). При скрещивании гибридов F1 с брама получено 12 черных и 8 светлых (всего 20). Как наследуется окраска? Определите генотипы пород.
7. От скрещивания светло-бежевых норок с коричневыми в первом поколении все щенки оказались коричневыми, а во втором получилось расщепление: 114 коричневых, 11 светло-бежевых, 35 черных и 38 платиновых. Как наследуются эти окраски у норок? Определите генотип исходных животных. Что получится, если скрестить черных особей из второго поколения со светло-бежевыми? Черных и платиновых норок?
8. Аллель А у крыс обуславливает желтую окраску шерсти. Аллель В другого гена вызывает развитие черной окраски шерсти. У особей А-В — шерсть серого цвета, а у особей aabb – белого. Серого самца скрестили с желтой самкой и получили в первом поколении 3/8 желтых, 3/8 серых, 1/8 черных и 1/8 белых крысят. Определите генотипы родителей.
9. При скрещивании сортов перца, имеющих желтые и коричневые плоды, в первом поколении получены растения с красными плодами. При дальнейшем разведении гибридов в себе получено 322 растения. Из них 182 с красными плодами, 59 — с коричневыми, 61 — с желтыми, и 20 с зелеными плодами.
10. При скрещивании кур и петухов имеющих ореховидные гребни получили 279 цыплят с ореховидным гребнем, 115 – с гороховидным, 106 — с розовидным и 35 с простым. Объясните результаты, определите генотипы исходных кур и петухов. Какая часть потомков с розовидным гребнем из F2 гомозиготна?
11. От скрещивании черного хорька со светло-коричневым в первом поколении все щенки были черными. При скрещивании хорьков из первого поколения между собой наблюдалось расщепление по фенотипам: черные, серые (В), коричневые (D) и светло-коричневые. Расщепление было близко к 9 : 3 : 3 : 1 соответственно. Напишите все генотипы (родителей и потомства).
12. При скрещивании черного и коричневого хорьков было получено 10 щенков, 6 из которых были коричневыми, а 4 – черными. Определите генотипы родителей и потомства. Какое расщепление по фенотипам и генотипам следует ожидать при скрещивании черного и коричневого хорьков из первого поколения?
13. При скрещивании двух черных хорьков в потомстве были получены черные и серые хорьки. Предположите, как распределились эти признаки среди 12 щенков. Какое потомство следует ожидать при скрещивании черных и серых хорьков из первого поколения между собой?
14. С какой вероятностью может появиться светло-коричневый щенок у черных родителей? Свой ответ подтвердите генотипами родителей и предполагаемого потомства.
15. При скрещивании коричневого хорька с черным в первом поколении были получены 7 черных и 2 серых щенка. Определите генотипы родителей потомства. Какое расщепление по фенотипам и генотипам следует ожидать при скрещивании серых хорьков из первого поколения между собой.
16 При скрещивании рябого петуха, имеющего простой гребень, с рябой курицей с ореховидным гребнем было получено следующее потомство: Петухи: 23 рябых с розовидным гребнем; 19 рябых с ореховидным гребнем. Куры: 12 рябых с ореховидным гребнем; 8 рябых с розовидным гребнем; 11 нерябых с ореховидным гребнем; 10 нерябых с розовидным гребнем. Объясните результаты, определите генотипы родителей и потомков.
17. От скрещивания двух зеленых растений кукурузы было получено 78 зеленых и 24 белых растения (альбиносы). При самоопылении одного из исходных зеленых растений получили 318 зеленых и 102 белых растений, а при самоопылении другого исходного зеленого растения — 274 зеленых и 208 белых. Каковы генотипы исходных зеленых растений? Объясните результаты скрещивания
18. У льна наследование окраски венчика происходит по типу комплементарного взаимодействия генов. Аллель А обуславливает окрашенный венчик, аллель а-неокрашенный, аллель В-голубой, в-розовый. 10 гибридных растений, имеющих генотип АаВв, были скрещены с растениями, имеющими генотип аавв. Было получено 80 растений.1.Сколько растений имели голубую окраску венчика? 2.сколько разных генотипов могут обусловить белую окраску венчика. 3.сколько растений имели розовую окраску венчика 4.сколько растений с белым венчиком были двойными гомозиготами
Эпистатическое взаимодействие генов
1. Бомбейский феномен (очень редкий, обнаруженный только в одной из популяций людей в Индии) у человека связан с присутствием в генотипе рецессивного эпистатического гена h, подавляющего проявление доминантных аллелей генов группы крови I А и I В. Поэтому гомозиготы с аллелями hh всегда имеют I группу крови. В близкородственном браке, где муж был с IV (I А I В ) группой, жена с III (I В ) и они имели общую бабушку, носительницу аллеля h, родилось 12 детей со следующими группами крови: IV (I А I В ) — 4 ребенка, III (I В ) — 5 детей и I (I О ) — 3 ребенка. Определите генотипы родителей и детей.
2. При скрещивании растения лука с красными луковицами с растением, имеющим белые луковицы, все растения в F1 были с красными луковицами, а в F2 наблюдалось расщепление по окраске луковицы: 195 растений имели красные луковицы, 71 - желтые и 89 были белые. Определите генотипы родителей и гибридов F1.
3. От скрещивания двух растений лука с красными луковицами №1 и №2 в F1 получилось 94 растений с красными и 28 с желтыми луковицами. Для выяснения генотипов растений №1 и №2 они были подвергнуты самоопылению. В результате самоопыления растения номер 1 получили 130 растений с красными, 42 — с желтыми и 61 — с белыми луковицами. А в результате самоопыления растения номер 2 получили 129 растений с красными и 40 с желтыми луковицами. Объясните результаты, определите генотипы исходных растений.
4. От скрещивания растений тыквы с белыми и зелеными плодами в F1 получено расщепление : 86 белых, 39 желтых, 42 зеленых плода. Найти генотипы исходных растений тыквы.
5. Рожь с черным зерном скрестили с рожью с серым зерном. В F2 были потомки с черным, серым и белым зерном. Эти признаки проявлялись в соотношении 12:3:1. Определите взаимодействие генов.
6. Имея две формы пшеницы – безостую с черными колосьями и остистую с красными колосьями, – селекционер скрестил их, чтобы получить безостую красноколосую форму. В F1 от этого скрещивания все растения были безостыми с черными колосьями. В F2 произошло расщепление: 714 безостых с черными колосьями, 181 безостых с красными колосьями, 58 безостых с белыми колосьями,231 остистых с черными колосьями, 61 остистых с красными колосьями, 20 остистых с белыми колосьями. Объясните расщепление. Приведите схему наследования признаков.
7. При скрещивании тыкв с белыми плодами в первом поколении получили 67 растений с белыми, 19 – с желтыми и 6 – с зелеными плодами. Объясните результаты, определите генотипы исходных растений. Что получится, если скрестить исходные растения с зеленоплодными из первого поколения?
8. От скрещивания растений кукурузы с окрашенными и неокрашенными зернами в первом поколении все зерна оказались неокрашенными, а во втором произошло расщепление 562 неокрашенных и 120 окрашенных. Как наследуется признак? Определить генотипы исходных растений.
9. кукурузы пурпурная окраска зерен определяется присутствием в генотипе гена А и отсутствием гена С во всех остальных случаях — зерна белые. Какое расщепление в первом поколении при следующем скрещивании: АаСс х аасс?
10. При скрещивании лошадей серой и рыжей масти в первом поколении всё потомство оказалось серым. Во втором поколении на каждые 16 лошадей в среднем появилось 12 — серых, 3 — вороных, 1 — рыжая. Определить тип наследования масти у лошадей и установить генотипы указанных животных.
11. У садовода имеются две формы левкоя с красными и кремовыми цветками. При самоопылении красноцветковых растений он получил в потомстве 222 красноцветковых, 68 растений с кремовыми цветками и 94 – с белыми. При самоопылении растений с кремовыми цветками получилось 18 растений с кремовыми и 7 с белыми цветками. При скрещивании исходных форм между собой получено 29 красноцветковых растений, 31 растение с кремовыми цветками и 18 растений с белыми. Как наследуется окраска у левкоя и каковы генотипы исходных форм, имеющихся у садовода?
12. Скрещивая растения левкоя с фиолетовыми цветками с растением, имеющим белые цветки, садовод обнаружил в первом поколении 102 растения с фиолетовыми, 109 с красными, 231 с кремовыми и 420 с белыми цветками. От самоопыления родительского белоцветкового растения все растения оказались с белыми цветками. При самоопылении родительского растения с фиолетовыми цветками в F1 обнаружилось расщепление в отношении: 27/64 с фиолетовыми : 9/64 с красными : 12/64 с кремовыми : 16/64 с белыми цветками. Объясните расщепления. Как наследуется окраска цветков у левкоев? Определите генотипы всех растений, использованных в скрещиваниях.
13. При скрещивании тыкв с белыми плодами в F1 получили 67 растений с белыми, 19 с желтыми и 6 с зелеными плодами. Объясните результаты, определите генотипы исходных растений. Что получится, если скрестить исходные растения с зеленоплодным из F1?
14. От скрещивания растений льна с белыми и голубыми лепестками цветков получены растения с голубыми лепестками. В следующем поколении идет расщепление в отношении 33 голубых : 12 розовых : 18 белых. Какое явление иллюстрирует данный случай и каковы генотипы всех форм?
15. Красная окраска луковицы лука определяется доминантым геном, желтая — его рецессивным аллелем. Однако проявление гена окраски возможно лишь при наличии другого, несцепленного с ним доминантного гена, рецессивный аллель которого подавляет окраску, и луковицы оказываются белыми. Краснолуковичное растение было скрещено с желтолуковичным. В потомстве оказались особи с красными, желтыми и белыми луковицами. Определите генотипы родителей и потомства.
16. У кур ген С детерминирует окраску оперения, ген с — белое оперение. Ген I подавляет развитие окраски, его рецессивный аллель i не оказывает подавляющего действия. Белый самец (IICC) скрещен с белой самкой (iicc). Укажите процент особей с окрашенным оперением во втором поколении.
17. При скрещивании голубых гуппи с альбиносами, имеющими белую окраску и красные глаза, в первом поколении было получено 69 голубых рыб, а во втором — 68 голубых, 20 белых с темными глазами и 34 альбиноса. Объясните расщепление. Как наследуется окраска? Определите генотипы исходных рыб. Какая часть рыб второго поколения гомозиготна по окраске?
18. При скрещивании лошадей серой и рыжей масти в первом поколении всё потомство оказалось серым. Во втором поколении на каждые 16 лошадей в среднем появилось 12 — серых, 3 — вороных,1 — рыжая. Определить тип наследования масти у лошадей и установить генотипы указанных животных.
19. У лошадей серая масть (ген «C») доминирует над рыжей (ген «с»), вороная (ген «В») над рыжей (ген «в»). Ген «С» эпистатичен по отношению к гену «В». При скрещивании серого жеребца с серой кобылой получен рыжий жеребенок. Каковы генотипы родителей? Какое потомство можно ожидать при повторении этих спариваний? Покажите это на решетке Пеннета.
20 При скрещивании двух белозерных растений кукурузы F1 получилось белозерное, а в F2 получено 138 белых семян и 39 пурпурных. К какому типу наследования относится этот случай? Определите генотипы всех форм.
1. Если скрещиваются между собой особи с генотипами А1А1а2а2 и а1а1А2А2, то какая часть потомства в F2 будет гетерозиготной по одному гену?
2. Имеется сорт ячменя, дающий 6 г зерна на растение, и сорт, дающий 12 г. При скрещивании этих сортов в F1 наблюдается промежуточный фенотип, в среднем 9 г. У 250 растений F2 масса зерна варьирует от 6 до 12 г на растение. Четыре растения имели массу 6 г, четыре других – 12 г. Сколько генов определяют данный признак? Определите генотипы всех растений.
3. Имеется сорт ячменя, дающий 6 г зерна на растение, и сорт, дающий 12 г. Каков будет фенотип растений F1 от скрещивания этих сортов, и как распределятся растения по весу семян на растение в F2, если учесть тригенное отличие этих сортов, гены не сцеплены и взаимодействуют по типу некумулятивной полимерии? Какое будет распределение растений по весу семян на растение, если растение F1 скрестить с малоурожайным сортом?
4. У родителей с темным цветом кожи родился первый ребенок с темным цветом кожи, а второй — с белым цветом кожи.
5. У одной из пары белых лабораторных крыс длина хвоста контролируется тремя парами несцепленных генов, которые взаимодействуют по типу полимерии. Если пренебречь факторами среды, то можно допустить, что самый длинный хвост (10см) определяется у крыс, имеющих все доминантные гены, крысы гомозиготные по трем парам рецессивных генов имеют самый короткий хвост (4см). Определить длину хвоста у крыс, гетерозиготных по всем трем парам генов. Определить длину хвоста у потомства от скрещивания гетерозиготного самца с короткохвостой самкой.
6. У кукурузы длина початка контролируется 2 парами полимерных генов по типу кумулятивной полимерии. Гомозигота по рецессивным генам имеет длину початка 10 см. Скрещивались два сорта кукурузы различного происхождения с длиной початка 22 см. В первом поколении все растения также имели початки длиной 22 см. Для гибридов F1 затем проводилось возвратное скрещивание с одной из родительских форм. Какой ожидается длина початков в потомстве от этого скрещивания и какова будет доля растений с различными длинами початков? Ответ подтвердите схемой скрещивания.
7. Растение гомозиготное по трем парам рецессивных генов, имеет высоту 32 см, а гомозиготное по доминантным аллелям этих генов имеет высоту 50 см. Принимаем, что влияние отдельных доминантных генов на рост во всех случаях одинаково и их действие суммируется. В F2 от скрещивания этих растений получено 192 потомка. Сколько из них будет иметь генетически обусловленный рост в 44 см?
8. Сколько будет фенотипических классов по окраске зерна и в каком количестве среди 2600 гибридов второго поколения пшеницы и овса, полученных от скрещивания ди — и три-гетерозигот, если известно, что у пшеницы 2, а у овса 3 полимерных гена определяют этот признак от темно-красного до белого зерна по типу кумулятивной полимерии.
9. В браке темнокожего мужчины с белокожей женщиной в F1 все дети мулаты. Дети от мулатов могут появиться с пигментацией от белого до очень смуглого. Нужно рассчитать вероятность рождения белого ребенка в таких семьях.
«Биология отрицает законы математики: при делении происходит умножение» Валерий Красовский
1. Комплементарное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое
Проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9 : 6 : 1, 9 : 3 : 4, 9 : 7, иногда 9 : 3 : 3 : 1.
2. Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным.
Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I.
Эпистатическое взаимодействие неаллельных генов
Может быть доминантным и рецессивным.
3. Полимерия — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным.
На мою электронную карту Сбербанка России : 2202 2008 1307 8159
Книга в формате.
Www. biorepet-ufa. ru
02.04.2018 20:53:09
2018-04-02 20:53:09
Взаимодействие неаллельных генов егэ биология

«Биология отрицает законы математики: при делении происходит умножение» Валерий Красовский
Взаимодействия аллельных и неаллельных генов

Теоретический вопрос ДНЯ
Кратко и понятно — схемы и конспекты (последние записи)
- Эффект бутылочного горлышка Теория симбиогенеза Клеточная теория Клетки человека Болото Тип Круглые черви — ароморфозы Тип Плоские черви — ароморфозы Правила эволюции Тип Кишечнополостные — ароморфозы Перья птиц
Последние обновления
- Класс Земноводные (Амфибии) Эффект бутылочного горлышка Головной мозг Спинной мозг Синапс Коферменты и их значение Тип Плоские черви Подцарство Простейшие Взаимодействия аллельных и неаллельных генов Цветок
Последние видео:
Подписывайся на обновления, обсуждай вопросы в соцсетях
17. От скрещивания двух зеленых растений кукурузы было получено 78 зеленых и 24 белых растения (альбиносы). При самоопылении одного из исходных зеленых растений получили 318 зеленых и 102 белых растений, а при самоопылении другого исходного зеленого растения — 274 зеленых и 208 белых. Каковы генотипы исходных зеленых растений? Объясните результаты скрещивания
Теоретический вопрос ДНЯ
- Эффект бутылочного горлышка Теория симбиогенеза Клеточная теория Клетки человека Болото Тип Круглые черви — ароморфозы Тип Плоские черви — ароморфозы Правила эволюции Тип Кишечнополостные — ароморфозы Перья птиц
- Класс Земноводные (Амфибии) Эффект бутылочного горлышка Головной мозг Спинной мозг Синапс Коферменты и их значение Тип Плоские черви Подцарство Простейшие Взаимодействия аллельных и неаллельных генов Цветок
Последние видео:
Подписывайся на обновления, обсуждай вопросы в соцсетях
6. У кукурузы длина початка контролируется 2 парами полимерных генов по типу кумулятивной полимерии. Гомозигота по рецессивным генам имеет длину початка 10 см. Скрещивались два сорта кукурузы различного происхождения с длиной початка 22 см. В первом поколении все растения также имели початки длиной 22 см. Для гибридов F1 затем проводилось возвратное скрещивание с одной из родительских форм. Какой ожидается длина початков в потомстве от этого скрещивания и какова будет доля растений с различными длинами початков? Ответ подтвердите схемой скрещивания.
ОПЛАТИТЬ МОЖНО:
Бомбейский феномен очень редкий, обнаруженный только в одной из популяций людей в Индии у человека связан с присутствием в генотипе рецессивного эпистатического гена h, подавляющего проявление доминантных аллелей генов группы крови I А и I В.
Biologyonline. ru
09.01.2019 7:50:21
2019-01-09 07:50:21
Источники:
Http://vk. com/wall-78321933_15718
Http://www. biorepet-ufa. ru/reshenie-geneticheskix-zadach/zadachi-po-genetike-na-vzaimodejstvie-neallelnyx-genov. html
Http://biologyonline. ru/index. php/2-uncategorised/230-vzaimodejstviya-allelnykh-i-neallelnykh-genov
Неаллельные гены. Селекция — Биология Егэ » /> » /> .keyword { color: red; } Взаимодействие неаллельных генов егэ биология
Взаимодействие неаллельных генов егэ биология
Задачи по генетике на Взаимодействие неаллельных генов. Комлементарность
Задача 1
У душистого горошка окраска цветов проявляется только при наличии двух доминантных генов А и В. Если в генотипе имеется только один доминантный ген, то окраска не развивается. Какое потомство F1 и F2 получится от скрещивания растений с генотипами ААbb и ааВВ?
Решение:
1. Генотип исходных растений известен по условию задачи. Они гомозиготны и будут давать один тип гамет, которые можно объединить единственным образом. Потомство F1 будет единообразно по генотипу (АаВb) и фенотипу (розовые цветы):
Схема первого скрещивания:
2. Дигетерозиготные потомки F1 будут давать по 4 типа гамет, которые могут комбинироваться 16 способами. Построив решетку Пеннета, можно убедиться, что организмов, одновременно имеющих доминантные гены А и В (розовые цветы), будет примерно 9/16. То есть, в F2 будет наблюдаться расщепление по цвету в пропорции 9:7.
Схема второго скрещивания
Ответ:
В F1 все потомство будет единообразным и будет иметь розовую окраску цветов. В F2 9/16 растений будут иметь розовые, а 7/16 – белые цветы.
Задача 2
При скрещивании двух растений тыквы со сферической формой плодов получено потомство, имеющее только дисковидные плоды. При скрещивании этих гибридов между собой были получены растения с тремя типами плодов:
• 9 частей – с дисковидными плодами,
• 6 частей – со сферической формой плодов,
• 1 часть – с удлиненными плодами.
Какая закономерность наблюдается в данном случае? Каковы генотипы родителей и потомства?
Решение:
1. Потомство F1 отличалось от родителей и было единообразным, следовательно, оно является гетерозиготным, а исходные особи были гомозиготными.
Схема первого скрещивания:
2. Исходные особи должны отличаться по генотипу друг от друга, так как при одинаковом генотипе их потомство не отличалось бы от них. Это указывает на то, что в данном случае имеет место комплементарное взаимодействие генов. Генотип родительских особей – BBcc и bbCC, а потомства F1 – BbCc.
3. Таким образом, форма плодов определяется двумя парами генов, В и С. Если обе пары находятся в доминантном состоянии (генотипы BBCC, BbCc, BBCc или BbCC), то форма плодов дисковидная. Если в доминантном состоянии находится только одна пара генов (генотипы BBcc, Bbcc, bbCC или bbCc), – форма плодов сферическая.
4. Расщепление в потомстве F2 (9:6:1) отличается от расщепления 9:7, обычно наблюдаемого при комплементарном взаимодействии. 1/16 часть растений имела плоды удлиненной формы. Это может быть только в том случае, если генотип таких особей – bbсс, то есть оба гена находятся в рецессивном состоянии.
Схема второго скрещивания:
5. Анализ решетки Пеннета подтверждает полученные результаты.
Задача 3
У норки известно два рецессивных гена – р и i, гомозиготность по каждому из которых, или по обоим одновременно, обуславливает платиновую окраску меха. Дикая коричневая окраска получается при наличии обоих доминантных аллелей Р и I. При каком типе скрещивания двух платиновых норок все их потомство будет коричневым?
Решение:
Платиновые норки должны быть обязательно гомозиготны по каждому из двух рецессивных генов — р и i или по обоим одновременно, т. е. генотип особей должен быть —Ppii, ppIi, ppII, PPii или ppii. Дикую коричневую окраску будут иметь особи с генотипом PpIi. В случае, если при скрещивании двух платиновых норок всё их потомство будет коричневым, генотип одного родителя должен быть гомозиготен по рецессивному гену р, а по гену i – гетерозиготен; другой родитель, наоборот, гомозиготен по гену i и гетерозиготен по гену р. Тогда для скрещивания необходимо взять животных с генотипами ppII и PPii. Что подтверждается схемой скрещивания.
Ответ:
Для скрещивания необходимо взять животных с генотипами ppII и PPii.
Задача 4
У кукурузы нормальный рост определяется двумя доминантными неаллельными генами. Гомозиготность по рецессивным аллелям даже одной пары генов приводит к возникновению карликовых форм. При скрещивании двух карликовых растений кукурузы выросли гибриды нормальной высоты, а при скрещивании этих гибридов в их потомстве было получено 812 нормальных и 640 карликовых растений. Определить генотипы родителей и потомков.
Решение:
1. Так как при скрещивании двух карликовых растений было получено потомство нормальной высоты. Потомство F1 отличалось от родителей и было единообразным, следовательно, оно является гетерозиготным, а исходные особи были гомозиготными. Таким образом, родители имели генотип ААbb и aaBB, а потомство — AaBb.
Схема первого скрещивания:
2. Дигетерозиготные потомки F1 будут давать по 4 типа гамет, которые могут комбинироваться 16 способами. Построив решетку Пеннета, можно убедиться, что организмов, одновременно имеющих доминантные гены А и В (нормальный рост), будет примерно 9/16. То есть, в F2 будет наблюдаться расщепление по карликовому росту в пропорции 9:7.
Схема второго скрещивания:
3. в потомстве F2 наблюдается расщепление 9:7, характерное для комплементарного наследования.
Ответ:
Исходные организмы имели генотипы ААbb и ааВВ. Единообразное потомство F1 несло генотип АаВb; в потомстве F2 наблюдалось расщепление 9:7, характерное для комплементарного наследования.
Задача 5
У лука ген R определяет красную окраску чешуй, а ген r – желтую. Любая окраска проявляется только при наличии в генотипе доминантного гена С, при его отсутствии чешуи имеют белую окраску. Определить генотипы исходных форм луковиц с белыми и красными чешуями, если все гибридные луковицы имели красную окраску чешуй.
Решение:
R — ген, определяющий красную окраску чешуи лука
R — ген, определяющий жёлтую окраску чешуи лука
C — ген, способствующий проявлению окраски
C — не влияет на окраску
1. Белая окраска луковиц может определяться генотипами ссRR, ccRr или ccrr, а красная — CcRR, CcRr или CCRR. Известно, что при скрещивании луковиц с белыми и красными чешуями получено потомство всё с красными луковицами, значит, в его генотипе присутствуют оба доминантных гена R и C.
2. Все потомство единообразно, следовательно, скрещивались гомозиготные особи. 3. Один родитель имел неокрашенные чешуи, значит, он мог нести оба рецессивных гена (c и r); другой родитель имел красную окраску чешуи, значит, он мог нести оба доминантных гена (C и R).
4. Этим условиям удовлетворяет только тот вариант, когда у одного родителя генотип ссrr, а у другого – CCRR.
Ответ:
Генотип исходных растений с белыми чешуями луковиц – ссrr, с красными чешуями луковиц – CCRR.
При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа I (например, обе формы неокрашены).
Широко известным примером рецессивного эпистаза является Бомбейский феномен, названный так в результате зафиксированного случая в индийском городе Бомбеи. Доктор Бхенде обнаружил, что у людей рецессивных по гену h (hh) на поверхности эритроцитов не синтезируются агглютиногены – в результате этого они могут быть универсальными донорами.
Эпистаз
Неаллельные гены это гены, расположенные в различных участках локусах хромосом и кодирующие неодинаковые белки.
Buzani. ru
03.09.2019 21:08:18
2019-09-03 21:08:18
Взаимодействие неаллельных генов егэ биология
Очень редко развитие того или иного признака определяется одним геном. Часто один ген обусловливает развитие нескольких признаков. Такое явление называетсяПлейотропным действием гена. Распространено и обратное — один признак контролируется несколькими генами.
Неаллельные гены — это гены, расположенные в различных участках (локусах) хромосом и кодирующие неодинаковые белки. Неаллельные гены могут взаимодействовать между собой, когда один признак проявляется под действием совокупности нескольких генов.
Выделяют три формы взаимодействия неаллельных генов:
Комплементарность
К комплементарным, или дополняющим друг друга, генам относятся такие неаллельные гены, которые при совместном проявлении обусловливают развитие нового признака. На примере наследования окраски цветков у душистого горошка можно понять сущность комплементарного действия генов. При скрещивании двух растений с белыми цветками у гибридов F1 цветки оказались пурпурными. При самоопылении растений из F1 в F2 наблюдалось расщепление растений по окраске цветков в отношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые у 7/16.

Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментом антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента — пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотипом ааВВ и ААbb имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором — есть пропигмент, но нет фермента, переводящего пропигмент в пигмент.

Проведем скрещивание двух растений душистого горошка с белыми цветками. В одной из линий, АAbb, есть доминантный аллель А, а у другой, ааBB, есть доминантный аллель В. У растений, имеющих доминантные гены А и В одновременно, есть и пропигмент (обеспечиваемый А), и фермент (обеспечиваемый В), необходимые для образования пурпурного пигмента. У гибридов F1 генотип AaBb, есть оба доминантных гена, поэтому они имеют пурпурные цветки. При самоопылении этих растений получаем F2. Результаты отражены в решётке Пеннета, розовым выделены генотипы тех растений, которые будут иметь пурпурные цветки.
| Гаметы | AB | Ab | AB | Ab |
| AB | AABB | AABb | AaBB | AaBb |
| Ab | AABb | AAbb | AaBb | Aabb |
| AB | AaBB | AaBb | AaBB | AaBb |
| Ab | AaBb | Aabb | AaBb | Aabb |
Комплементарное взаимодействие может давать не два, а три фенотипа. Это происходит в тех случаях, когда двойные рецессивные гомозиготы aabb по фенотипу отличаются от рецессивных гомозигот только по одному гену — aaBb, Aabb. В этом случае расщепление будет 9:6:1. Например, это наблюдается при наследовании формы плодов у тыквы:

Возможно и возникновение четырёх фенотипов. Например, при скрещивании кур с различной формой гребня.

Aabb — простой (листовидный)
Эпистаз
Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется Ингибитором и обозначается буквой I.
Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным.
ДОМИНАНТНЫЙ ЭПИСТАЗ
При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа I (например, обе формы неокрашены).
Наследование окраски шерсти у собак (пример доминантного эпистаза):
A — черная окраска, а — коричневая, I — подавляет окраску, i — не подавляет.
РЕЦЕССИВНЫЙ ЭПИСТАЗ
Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена в гомозиготном состоянии аллелей гипостатичного гена (ii > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа ii (например, обе формы неокрашены).
Рецессивный эпистаз проявляется при наследовании окраски шерсти у домовых мышей.
А — окраска агути (рыжевато-серая)
А — черная окраска
В — способствует проявлению окраски
B — супрессор (подавляет действие А и А)
Мыши с генотипом А-bb и ааbb имеют одинаковый фенотип — все белые.
Полимерия
Полимерия — взаимодействие неаллельных генов, при котором степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного гена имеют одинаковый нижний индекс.
Например, у пшеницы А — тёмно-красный цвет зёрен, а — белый цвет зёрен. За цвет отвечают два гена — 1 и 2. Первый ген может быть представлен аллелями А1 и а1, второй — аллелями А2 и а2.
В зависимости от того, каких аллелей больше — А или а, оттенок зёрен будет изменяться.

Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от числа доминантных аллелей всех генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1 для двух генов, 64:1 для трёх генов и т. д. Например, цвет кожи у людей зависит от четырёх генов, поэтому от брака двух мулатов практически никогда не рождаются дети с белой кожей.
Разберём решение задач с неаллельным взаимодействием генов.
При скрещивании двух божьих коровок с красными надкрыльями все гибриды первого поколения имели оранжевые надкрылья. Их скрестили между собой и получили во втором поколении 115 жуков с оранжевыми надкрыльями, 79 с красными надкрыльями и 13 с жёлтыми надкрыльями. Как наследуется цвет надкрылий?
РЕШЕНИЕ
Так как в первом поколении все потомки одинаковы, родители гомозиготны. Появление во втором поколении третьей окраски и численные соотношения указывают на то, что это дигибридное скрещивание. Значит, в потомстве имеется 16 генотипических классов. Общее число потомков второго поколения = 115 + 79 + 13 = 207. Размер генотипического класса = 207 : 16 = 13. С оранжевой окраской 115 : 13 = 9 генотипических классов. С красной окраской 79 : 13 = 6 генотипических классов. С жёлтой окраской 13 : 13 + 1 генотипический класс. Таким образом, во втором поколении расщепление 9:6:1, что соответствует комплементарному взаимодействию генов. Родительские формы ААbbи ааВВ, гибриды первого поколения АаВb. Во втором поколении 9 А__В_ : 6 (ааВ_ + А_bb) :1 ааbb.
Селекция — наука, разрабатывающая пути создания новых и улучшения существующих сортов растений, пород животных и штаммов микроорганизмов.
Создание новых сортов и пород основывается на таких важнейших свойствах живого организма, как наследственность и изменчивость. Именно поэтому генетика — наука об изменчивости и наследственности организмов — является теоретической основой селекции.
Имея свои собственные задачи и методы, селекция твердо опирается на законы генетики, является важной областью практического использования закономерностей, установленных генетикой. Вместе с тем селекция опирается и на достижения других наук. На сегодняшний день генетика вышла на уровень целенаправленного конструирования организмов с нужными признаками и свойствами.
Сорт, порода и штамм — устойчивая группа организмов, искусственно созданная человеком и имеющая определенные наследственные особенности.
Все особи внутри породы, сорта и штамма имеют сходные, наследственно закрепленные морфологические, физиолого-биохимические и хозяйственные признаки и свойства, а также однотипную реакцию на факторы внешней среды.
Основные направления селекции:
- высокая урожайность сортов растений, плодовитость и продуктивность пород животных; улучшение качества продукции (например, вкус, внешний вид плодов и овощей, химический состав зерна — содержание белка, клейковины, незаменимых аминокислот и т. д.); физиологические свойства (скороспелость, засухоустойчивость, зимостойкость, устойчивость к болезням, вредителям и неблагоприятным климатическим условиям).
- выведение стрессоустойчивых пород (для разведения в условиях большой скученности — на птицефабриках, фермах и т. п.); пушное звероводство; рыбоводство — разведение рыбы в искусственных водоемах.
ОТЛИЧИЕ КУЛЬТУРНЫХ ФОРМ ОТ ДИКИХ
| Культурные формы | Дикие формы |
| Развиты признаки, полезные для человека и часто вредные в естественных условиях | Наличие признаков, неудобных для человека (агрессивность, колючесть и т. п.) |
| Высокая продуктивность | Низкая продуктивность (мелкие плоды; низкая масса, яйценоскость, удойность) |
| Хуже адаптируются к меняющимся условиям среды | Высокая адаптивность |
| Не имеют средств защиты от хищников и вредителей (горьких или ядовитых веществ, шипов, колючек и т. п.) | Наличие естественных защитных приспособлений, повышающих жизнестойкость, но неудобных для человека |
Основные методы селекции
Основные методы селекции:
ПОДБОР РОДИТЕЛЬСКИХ ПАР
Данный метод применяется прежде всего в селекции животных, т. к. для животных характерно половое размножение и немногочисленное потомство.
Выведение новой породы — процесс длительный, требующий больших материальных затрат. Это может быть целенаправленное получение определенного Экстерьера(совокупности фенотипических признаков), повышение молочности, жирности молока, качества мяса и т. д.
Разводимые животные оцениваются не только по внешним признакам, но и по происхождению и Качеству потомства. Поэтому необходимо хорошо знать их родословную. В племенных хозяйствах при подборе производителей всегда ведется учёт родословных, в которых оцениваются экстерьерные особенности и продуктивность родительских форм в течение ряда поколений.
РАБОТЫ И. В. МИЧУРИНА
ОТБОР
Искусственный отбор — сохранение для дальнейшего размножения особей с интересующими селекционера признаками. Формы отбора: массовый и индивидуальный.
- Интуитивный (бессознательный) отбор — самая древняя форма отбора, используемая ещё древним человеком: отбор особей по фенотипу, т. е. с наиболее полезными сочетаниями признаков. Методический отбор — отбор для размножения особей с чётко определёнными признаками, согласно цели и с учетом их фенотипов и генотипов.
- Массовый отбор — устранение из размножения особей, не имеющих ценные признаки, либо имеющих нежелательные признаки (например, агрессивных).
Массовый отбор может быть эффективен в том случае, если отбираются качественные, просто наследуемые и легко определяемые признаки. Массовый отбор обычно проводят среди перекрестноопыляемых растений. При этом селекционеры отбирают растения по фенотипу с интересующими их признаками. Недостаток массового отбора заключается в том, что селекционер не всегда может определить лучший генотип по фенотипу.
- Индивидуальный отбор — выделение отдельных особей с интересующими человека признаками и получение от них потомства.
Индивидуальный отбор более эффективен при отборе особей по количественным, сложно наследуемым признакам. Этот вид отбора позволяет точно оценить генотип благодаря анализу наследования признаков у потомства. Индивидуальный отбор применяют по отношению к самоопыляемым растениям (сорта пшеницы, ячменя, гороха и др.).
ГИБРИДИЗАЦИЯ
В селекционной работе с животными применяют в основном два способа скрещивания:Инбридинг и Аутбридинг.
Инбридинг — скрещивание близкородственных форм: в качестве исходных форм используются братья и сестры или родители и потомство.
Результат: получение гомозиготных организмов → разложение исходной формы на ряд чистых линий.
Минусы: пониженная жизнеспособность (рецессивные гомозиготы зачастую несут наследственные заболевания).
Такое скрещивание в определённой степени аналогично самоопылению у растений, которое также приводит к повышению гомозиготности и, как следствие, к закреплению хозяйственно ценных признаков у потомков. При этом гомозиготизация по генам, контролирующим изучаемый признак, происходит тем быстрее, чем более близкородственное скрещивание используют при инбридинге. Однако гомозиготизация при инбридинге, как и в случае растений, ведет к ослаблению животных, снижает их устойчивость к воздействию среды, повышает заболеваемость.
В селекции инбридинг обычно является лишь одним из этапов улучшения породы. За ним следует скрещивание разных межлинейных гибридов, в результате которого нежелательные рецессивные аллели переводятся в гетерозиготное состояние и вредные последствия близкородственного скрещивания заметно снижаются.
Аутбридинг — неродственное скрещивание между особями одной породы или разных пород животных в пределах одного вида.
Результат: получение большого количества гетерозиготных организмов → поддержание полезных качеств и усиление их выраженности в ряду следующих поколений.
Отдалённая гибридизация — получение межвидовых и межродовых гибридов.
Отдалённая гибридизация в селекции животных применяется значительно реже, чем в селекции растений.
Межвидовые и межродовые гибриды животных и растений чаще всего бесплодны, так как нарушается мейоз и гаметогенез не происходит. При этом восстановление плодовитости у животных представляет более сложную задачу, поскольку получение полиплоидов на основе умножения числа хромосом у них невозможно.
Преодоление бесплодия межвидовых гибридов растений впервые удалось осуществить в начале 20-х годов ХХ века советскому генетику Г. Д. Карпеченко при скрещивании редьки и капусты. Это вновь созданное человеком растение не было похоже ни на редьку, ни на капусту. Стручки занимали как бы промежуточное положение и состояли из двух половинок, из которых одна напоминала стручок капусты, другая — редьки. Каждая из исходных форм имела в половых клетках по 9 хромосом. В этом случае клетки полученного от них гибрида имели 18 хромосом. Но некоторые яйцеклетки и пыльцевые зёрна содержали все 18 хромосом (диплоиды), а при их скрещивании создано растение с 36 хромосомами, которое оказалось плодовитым. Так была доказана возможность использования полиплоида для преодоления нескрещиваемости и бесплодия при отдалённой гибридизации.
Бывает, что бесплодны особи только одного пола. Например, у гибридов высокогорного быка яка и рогатого скота бесплодны (стерильны) самцы, а самки плодовиты(фертильны).
Но иногда гаметогенез у отдалённых гибридов протекает нормально, что позволило получить новые ценные породы животных. Примером являются архаромериносы, которые, как и архары (горные бараны), могут пастись высоко в горах, а как мериносы дают хорошую шерсть. Получены плодовитые гибриды от скрещивания местного (индийского) крупного рогатого скота с зебу. При скрещивании белуги и стерляди получен плодовитый гибрид — бестер, хорька и норки — хонорик, продуктивен гибрид между карпом и карасём.
В природе встречаются гибриды зебры и лошади (зеброид), бизона и зубра (зубробизон), тетерева и куропатки (межняк), зайца-русака и зайца-беляка (тумак), соболя и лисицы (кидус), а также тигра и льва (лигр).
В качестве примеров межродовых гибридов растений можно назвать гибрид пшеницы и ржи (тритикале), пшенично-пырейный гибрид, гибрид смородины и крыжовника (йошта), гибрид брюквы и кормовой капусты (куузика), гибриды озимой ржи и житняка, травянистого и древовидного томатов и др.
Гетерозис — явление повышенной жизнеспособности, урожайности, плодовитости гибридов первого поколения, превышающих по этим параметрам обоих родителей.
Уже со второго поколения гетерозисный эффект угасает. По-видимому, это происходит вследствие снижения числа гетерозиготных организмов и повышения доли гомозигот.
Классическими примерами проявления гетерозиса являются мул (гибрид кобылы и осла) и лошак (гибрид коня и ослицы) (рис. 1,2) . Это сильные, выносливые животные, которые могут использоваться в значительно более трудных условиях, чем родительские формы.


Рис. 1. Мул Рис. 2. Лошак
Продолжительность их жизни значительно выше, чем у родительских видов.
Лошак меньше мула ростом и строптив, поэтому менее удобен для использования в хозяйственной деятельности человека.
Гетерозис широко применяют в промышленном птицеводстве, например — бройлерные цыплята, отличающиеся очень быстрым ростом. Цыплёнок-бройлер — финальный гибрид, полученный в результате скрещивания нескольких линий разных пород кур (мясных родительских форм), проверенных на сочетаемость. Первоначально для такого скрещивания использовали породы корниш (в качестве отцовской формы) и белый плимутрок (в качестве материнской формы).
ИСКУССТВЕННЫЙ МУТАГЕНЕЗ
Искусственный мутагенез чаще всего используется как метод селекции растений. Он основан на применении физических и химических мутагенов для получения форм растений с выраженными мутациями. Такие формы в дальнейшем используются для гибридизации или отбора.
В селекции растений широко используется Полиплоидия.
Полиплоидия — увеличение числа наборов хромосом в клетках организма, кратное гаплоидному (одинарному) числу хромосом; тип геномной мутации.
Половые клетки большинства организмов гаплоидны (содержат один набор хромосом — n), соматические — диплоидны (2n). Организмы, клетки которых содержат более двух наборов хромосом, называются полиплоидами, три набора — триплоидами (3n), четыре — тетраплоидами (4n) и т. д. Наиболее часто встречаются организмы с числом хромосомных наборов, кратным двум, — тетраплоиды, гексаплоиды (6n) и т. д.
Полиплоиды с нечётным числом наборов хромосом (триплоиды, пентаплоиды и т. д.) обычно не дают потомства (стерильны), т. к. образуемые ими половые клетки содержат неполный набор хромосом — не кратный гаплоидному.
ПОЯВЛЕНИЕ ПОЛИПЛОИДИИ
Благодаря полиплоидии выведены высокоурожайные полиплоидные сорта сахарной свеклы, хлопчатника, гречихи и др. Полиплоидные растения часто более жизнеспособны и плодовиты, чем нормальные диплоиды. О их большей устойчивости к холоду свидетельствует увеличение числа видов-полиплоидов в высоких широтах и в высокогорьях.
Поскольку полиплоидные формы часто обладают ценными хозяйственными признаками, искусственную полиплоидизацию применяют в растениеводстве для получения исходного селекционного материала.
Получение полиплоидов в эксперименте тесно связано с искусственным мутагенезом. С этой целью используют специальные мутагены (например, алкалоид колхицин), нарушающие расхождение хромосом в митозе и мейозе.
Получены урожайные полиплоиды ржи, гречихи, сахарной свёклы и других культурных растений; стерильные триплоиды арбуза, винограда, банана популярны благодаря бессемянным плодам.
Применение отдалённой гибридизации в сочетании с искусственной полиплоидизацией позволило отечественным учёным получить плодовитые полиплоидные гибриды растений (Г. Д. Карпеченко, гибрид-тетраплоид редьки и капусты) и животных (Б. Л. Астауров, гибрид-тетраплоид тутового шелкопряда).
ШЕЛКОПРЯДЫ АСТАУРОВА
Биотехнология
Биотехнология — наука, изучающая возможность модификации биологических организмов для обеспечения потребностей человека.
Применение биотехнологии (рис. 3):
- производство лекарств, удобрений, средств биологической защиты растений; биологическая очистка сточных вод; восстановление ценных металлов из морской воды; коррекция и исправление генетических патологий.

Рис. 3. Возможности биотехнологии
Например, включение в геном кишечной палочки гена, ответственного за образование у человека инсулина, позволило наладить промышленное получение этого гормона (рис. 4).

Рис. 4. Биотехнология получения инсулина
В биотехнологии успешно применяются методы генной и клеточной инженерии.
ГЕННАЯ И КЛЕТОЧНАЯ ИНЖЕНЕРИЯ
Генная инженерия — искусственное, целенаправленное изменение генотипа микроорганизмов с целью получения культур с заранее заданными свойствами.
Исследования в области генной инженерии распространяются не только на микроорганизмы, но и на человека. Они особенно актуальны при лечении болезней, связанных с нарушениями в иммунной системе, в системе свертывания крови, в онкологии.
Основной метод генной инженерии: выделение необходимых генов, их клонирование и введение в новую генетическую среду. Например, введение определённых генов с помощью плазмиды в организм бактерии для синтеза ею определённого белка (рис. 5).

Рис. 5. Применение генной инженерии
Основные этапы решения генно-инженерной задачи следующие:
Получение изолированного гена. Введение гена в вектор (плазмиду) для переноса в организм. Перенос вектора с геном (рекомбинантной плазмиды) в модифицируемый организм. Преобразование клеток организма. Отбор генетически модифицированных организмов и устранение тех, которые не были успешно модифицированы.
Клеточная инженерия — это направление в науке и селекционной практике, которое изучает методы гибридизации соматических клеток, принадлежащих разным видам, возможности клонирования тканей или целых организмов из отдельных клеток.
Включает культивирование и клонирование клеток на специально подобранных средах, гибридизацию клеток, пересадку клеточных ядер и другие микрохирургические операции по «разборке» и «сборке» (реконструкции) жизнеспособных клеток из отдельных фрагментов.
На данный момент удалось получить гибриды между клетками животных, далёких по систематическому положению, например мыши и курицы. Соматические гибриды нашли широкое применение как в научных исследованиях, так и в биотехнологии.
Гибридные клетки, полученные от клеток человека и мыши и человека и китайского хомячка, участвовали в расшифровке генома человека.
Гибриды между опухолевыми клетками и лимфоцитами обладают свойствами обеих родительских клеточных линий: они неограниченно делятся и могут вырабатывать определённые антитела. Такие антитела применяют в лечебных и диагностических целях в медицине.
В эмбриологии для изучения процессов дифференцировки клеток и тканей в ходе онтогенеза используют организмы — Химеры, состоящие из клеток с разными генотипами . Их создают путём соединения клеток разных зародышей на ранних этапах их развития.
Клонирование животных — ещё один метод клеточной инженерии: ядро соматической клетки пересаживают в лишённую ядра яйцеклетку с последующим выращиванием зародыша во взрослый организм.
Преимущество клеточной инженерии в том, что она позволяет экспериментировать с клетками, а не с целыми организмами.
Методы клеточной инженерии часто применяют в сочетании с генной инженерией.
Работы Н. И. Вавилова
Николай Иванович Вавилов — российский генетик, растениевод, географ.
Н. И. Вавилов организовал 180 экспедиций (20−30 гг. ХХ века) по самым труднодоступным и зачастую опасным районам земного шара с целью изучения многообразия и географического распространения культурных растений. Им была собрана уникальная, самая крупная в мире коллекция культурных растений (к 1940 г. коллекция включала 300 000 образцов), которые ежегодно размножаются в коллекциях Всероссийского института растениеводства имени Н. И. Вавилова (ВИР) и широко используются селекционерами как исходный материал для создания новых сортов зерновых, плодовых, овощных, технических, лекарственных и других культур. Создал учение об иммунитете растений.
УЧЕНИЕ ОБ ИММУНИТЕТЕ РАСТЕНИЙ
ЦЕНТРЫ ПРОИСХОЖДЕНИЯ КУЛЬТУРНЫХ РАСТЕНИЙ
Центры происхождения культурных растений — географические области, являющиеся родиной дикорастущих предков культурных растений.
Центры происхождения важнейших культурных растений связаны с древними очагами цивилизации и местом первичного возделывания и селекции растений. Подобные очаги одомашнивания (центры Доместикации) выявлены и у домашних животных.
Было выделено восемь центров происхождения культурных растений (рис. 6):
1. Средиземноморский (спаржа, маслины, капуста, лук, клевер, мак, свекла, морковь).
2. Переднеазитский (инжир, миндаль, виноград, гранат, люцерна, рожь, дыня, роза).
3. Среднеазиатский (нут, абрикос, горох, груша, чечевица, лен, чеснок, мягкая пшеница).
4. Индо-Малайский (цитрусовые, хлебное дерево, огурец, манго, черный перец, кокосовая пальма, банан, баклажан).
5. Китайский (просо, редька, вишня, яблоко, гречиха, слива, соя, хурма).
6. Центральноамериканский (тыква, фасоль, какао, авокадо, махорка, кукуруза, батат, хлопчатник).
7. Южноамериканский (табак, ананас, томат, картофель).
8. Абиссинский центр (банан, кофе, сорго, твердая пшеница).
В поздних работах Н. И. Вавилова Переднеазиатский и Среднеазиатский центры объединяются в Юго-западноазиатский центр.

Рис. 6. Центры происхождения культурных растений
В настоящее время выделяют 12 первичных центров происхождения культурных растений.
При скрещивании растений с красными цветками (АА) и растений с белыми (аа) гибриды F1 имеют розовые цветки (Аа). Таким образом, имеет место неполное доминирование; в F2 наблюдается расщепление 1 : 2 : 1 как по фенотипу, так и по генотипу.
ОТЛИЧИЕ КУЛЬТУРНЫХ ФОРМ ОТ ДИКИХ
| Культурные формы | Дикие формы |
| Развиты признаки, полезные для человека и часто вредные в естественных условиях | Наличие признаков, неудобных для человека (агрессивность, колючесть и т. п.) |
| Высокая продуктивность | Низкая продуктивность (мелкие плоды; низкая масса, яйценоскость, удойность) |
| Хуже адаптируются к меняющимся условиям среды | Высокая адаптивность |
| Не имеют средств защиты от хищников и вредителей (горьких или ядовитых веществ, шипов, колючек и т. п.) | Наличие естественных защитных приспособлений, повышающих жизнестойкость, но неудобных для человека |
Основные методы селекции:
ПОДБОР РОДИТЕЛЬСКИХ ПАР
Данный метод применяется прежде всего в селекции животных, т. к. для животных характерно половое размножение и немногочисленное потомство.
Выведение новой породы — процесс длительный, требующий больших материальных затрат. Это может быть целенаправленное получение определенного Экстерьера(совокупности фенотипических признаков), повышение молочности, жирности молока, качества мяса и т. д.
Разводимые животные оцениваются не только по внешним признакам, но и по происхождению и Качеству потомства. Поэтому необходимо хорошо знать их родословную. В племенных хозяйствах при подборе производителей всегда ведется учёт родословных, в которых оцениваются экстерьерные особенности и продуктивность родительских форм в течение ряда поколений.
РАБОТЫ И. В. МИЧУРИНА
ОТБОР
Искусственный отбор — сохранение для дальнейшего размножения особей с интересующими селекционера признаками. Формы отбора: массовый и индивидуальный.
- Интуитивный (бессознательный) отбор — самая древняя форма отбора, используемая ещё древним человеком: отбор особей по фенотипу, т. е. с наиболее полезными сочетаниями признаков. Методический отбор — отбор для размножения особей с чётко определёнными признаками, согласно цели и с учетом их фенотипов и генотипов.
- Массовый отбор — устранение из размножения особей, не имеющих ценные признаки, либо имеющих нежелательные признаки (например, агрессивных).
Массовый отбор может быть эффективен в том случае, если отбираются качественные, просто наследуемые и легко определяемые признаки. Массовый отбор обычно проводят среди перекрестноопыляемых растений. При этом селекционеры отбирают растения по фенотипу с интересующими их признаками. Недостаток массового отбора заключается в том, что селекционер не всегда может определить лучший генотип по фенотипу.
- Индивидуальный отбор — выделение отдельных особей с интересующими человека признаками и получение от них потомства.
Индивидуальный отбор более эффективен при отборе особей по количественным, сложно наследуемым признакам. Этот вид отбора позволяет точно оценить генотип благодаря анализу наследования признаков у потомства. Индивидуальный отбор применяют по отношению к самоопыляемым растениям (сорта пшеницы, ячменя, гороха и др.).
ГИБРИДИЗАЦИЯ
В селекционной работе с животными применяют в основном два способа скрещивания:Инбридинг и Аутбридинг.
Инбридинг — скрещивание близкородственных форм: в качестве исходных форм используются братья и сестры или родители и потомство.
Результат: получение гомозиготных организмов → разложение исходной формы на ряд чистых линий.
Минусы: пониженная жизнеспособность (рецессивные гомозиготы зачастую несут наследственные заболевания).
Такое скрещивание в определённой степени аналогично самоопылению у растений, которое также приводит к повышению гомозиготности и, как следствие, к закреплению хозяйственно ценных признаков у потомков. При этом гомозиготизация по генам, контролирующим изучаемый признак, происходит тем быстрее, чем более близкородственное скрещивание используют при инбридинге. Однако гомозиготизация при инбридинге, как и в случае растений, ведет к ослаблению животных, снижает их устойчивость к воздействию среды, повышает заболеваемость.
В селекции инбридинг обычно является лишь одним из этапов улучшения породы. За ним следует скрещивание разных межлинейных гибридов, в результате которого нежелательные рецессивные аллели переводятся в гетерозиготное состояние и вредные последствия близкородственного скрещивания заметно снижаются.
Аутбридинг — неродственное скрещивание между особями одной породы или разных пород животных в пределах одного вида.
Результат: получение большого количества гетерозиготных организмов → поддержание полезных качеств и усиление их выраженности в ряду следующих поколений.
Отдалённая гибридизация — получение межвидовых и межродовых гибридов.
Отдалённая гибридизация в селекции животных применяется значительно реже, чем в селекции растений.
Межвидовые и межродовые гибриды животных и растений чаще всего бесплодны, так как нарушается мейоз и гаметогенез не происходит. При этом восстановление плодовитости у животных представляет более сложную задачу, поскольку получение полиплоидов на основе умножения числа хромосом у них невозможно.
Преодоление бесплодия межвидовых гибридов растений впервые удалось осуществить в начале 20-х годов ХХ века советскому генетику Г. Д. Карпеченко при скрещивании редьки и капусты. Это вновь созданное человеком растение не было похоже ни на редьку, ни на капусту. Стручки занимали как бы промежуточное положение и состояли из двух половинок, из которых одна напоминала стручок капусты, другая — редьки. Каждая из исходных форм имела в половых клетках по 9 хромосом. В этом случае клетки полученного от них гибрида имели 18 хромосом. Но некоторые яйцеклетки и пыльцевые зёрна содержали все 18 хромосом (диплоиды), а при их скрещивании создано растение с 36 хромосомами, которое оказалось плодовитым. Так была доказана возможность использования полиплоида для преодоления нескрещиваемости и бесплодия при отдалённой гибридизации.
Бывает, что бесплодны особи только одного пола. Например, у гибридов высокогорного быка яка и рогатого скота бесплодны (стерильны) самцы, а самки плодовиты(фертильны).
Но иногда гаметогенез у отдалённых гибридов протекает нормально, что позволило получить новые ценные породы животных. Примером являются архаромериносы, которые, как и архары (горные бараны), могут пастись высоко в горах, а как мериносы дают хорошую шерсть. Получены плодовитые гибриды от скрещивания местного (индийского) крупного рогатого скота с зебу. При скрещивании белуги и стерляди получен плодовитый гибрид — бестер, хорька и норки — хонорик, продуктивен гибрид между карпом и карасём.
В природе встречаются гибриды зебры и лошади (зеброид), бизона и зубра (зубробизон), тетерева и куропатки (межняк), зайца-русака и зайца-беляка (тумак), соболя и лисицы (кидус), а также тигра и льва (лигр).
В качестве примеров межродовых гибридов растений можно назвать гибрид пшеницы и ржи (тритикале), пшенично-пырейный гибрид, гибрид смородины и крыжовника (йошта), гибрид брюквы и кормовой капусты (куузика), гибриды озимой ржи и житняка, травянистого и древовидного томатов и др.
Гетерозис — явление повышенной жизнеспособности, урожайности, плодовитости гибридов первого поколения, превышающих по этим параметрам обоих родителей.
Уже со второго поколения гетерозисный эффект угасает. По-видимому, это происходит вследствие снижения числа гетерозиготных организмов и повышения доли гомозигот.
Классическими примерами проявления гетерозиса являются мул (гибрид кобылы и осла) и лошак (гибрид коня и ослицы) (рис. 1,2) . Это сильные, выносливые животные, которые могут использоваться в значительно более трудных условиях, чем родительские формы.
Рис. 1. Мул Рис. 2. Лошак
Продолжительность их жизни значительно выше, чем у родительских видов.
Лошак меньше мула ростом и строптив, поэтому менее удобен для использования в хозяйственной деятельности человека.
Гетерозис широко применяют в промышленном птицеводстве, например — бройлерные цыплята, отличающиеся очень быстрым ростом. Цыплёнок-бройлер — финальный гибрид, полученный в результате скрещивания нескольких линий разных пород кур (мясных родительских форм), проверенных на сочетаемость. Первоначально для такого скрещивания использовали породы корниш (в качестве отцовской формы) и белый плимутрок (в качестве материнской формы).
ИСКУССТВЕННЫЙ МУТАГЕНЕЗ
Искусственный мутагенез чаще всего используется как метод селекции растений. Он основан на применении физических и химических мутагенов для получения форм растений с выраженными мутациями. Такие формы в дальнейшем используются для гибридизации или отбора.
В селекции растений широко используется Полиплоидия.
Полиплоидия — увеличение числа наборов хромосом в клетках организма, кратное гаплоидному (одинарному) числу хромосом; тип геномной мутации.
Половые клетки большинства организмов гаплоидны (содержат один набор хромосом — n), соматические — диплоидны (2n). Организмы, клетки которых содержат более двух наборов хромосом, называются полиплоидами, три набора — триплоидами (3n), четыре — тетраплоидами (4n) и т. д. Наиболее часто встречаются организмы с числом хромосомных наборов, кратным двум, — тетраплоиды, гексаплоиды (6n) и т. д.
Полиплоиды с нечётным числом наборов хромосом (триплоиды, пентаплоиды и т. д.) обычно не дают потомства (стерильны), т. к. образуемые ими половые клетки содержат неполный набор хромосом — не кратный гаплоидному.
ПОЯВЛЕНИЕ ПОЛИПЛОИДИИ
Благодаря полиплоидии выведены высокоурожайные полиплоидные сорта сахарной свеклы, хлопчатника, гречихи и др. Полиплоидные растения часто более жизнеспособны и плодовиты, чем нормальные диплоиды. О их большей устойчивости к холоду свидетельствует увеличение числа видов-полиплоидов в высоких широтах и в высокогорьях.
Поскольку полиплоидные формы часто обладают ценными хозяйственными признаками, искусственную полиплоидизацию применяют в растениеводстве для получения исходного селекционного материала.
Получение полиплоидов в эксперименте тесно связано с искусственным мутагенезом. С этой целью используют специальные мутагены (например, алкалоид колхицин), нарушающие расхождение хромосом в митозе и мейозе.
Получены урожайные полиплоиды ржи, гречихи, сахарной свёклы и других культурных растений; стерильные триплоиды арбуза, винограда, банана популярны благодаря бессемянным плодам.
Применение отдалённой гибридизации в сочетании с искусственной полиплоидизацией позволило отечественным учёным получить плодовитые полиплоидные гибриды растений (Г. Д. Карпеченко, гибрид-тетраплоид редьки и капусты) и животных (Б. Л. Астауров, гибрид-тетраплоид тутового шелкопряда).
ШЕЛКОПРЯДЫ АСТАУРОВА
В случае если в одной аллели находится два доминантных гена, то, как правило, их действие и проявление сильно хуже, чем в предыдущем варианте с одним доминантным и одним рецессивным геном.
Биология в лицее
Как наследуется цвет надкрылий.
Sites. google. com
04.05.2018 10:34:19
2018-05-04 10:34:19
Взаимодействие аллельных генов в организме – формы и способы, биологическое значение
Вы уже знаете о том, что гены могут взаимодействовать друг с другом по типу полного и неполного доминирования. Однако, в генетике встречается масса других примеров взаимодействия генов. В этой статье мы затронем те, которые ранее не обсуждались.
Кодоминирование
Кодоминирование – взаимодействия аллельных генов, при котором в гетерозиготном состоянии могут оказаться два доминантных гена одновременно, при этом каждый ген отвечает за свой признак.
Наиболее распространенным примером кодоминирования является наследование групп крови у человека.
Решим пару задач, которые укрепят понимание темы.
Пример решения задачи №1
“Родители имеют II и III группы крови, гетерозиготны. Какие группы крови можно ожидать у их детей?”
Гетерозиготный генотип матери – I A i 0 и генотип отца – I B i 0 . Составим схему решения для такого случая.
Итак, в результате такого брака может получиться ребенок с любой группой крови, в чем мы убедились.
Пример решения задачи №2
“Дигетерозиготная по B (III) группе и положительному резус-фактору вступила в брак с таким же мужчиной. Какое расщепление по фенотипу можно ожидать у детей?”
Сходу понятно, что гетерозиготы по III (B) группе крови будут записаны I B i 0 . Резус-фактор для нас новое понятие – это белок, находящийся на поверхности эритроцита (тогда говорят, что резус-фактор положителен), или отсутствующий (тогда у человека резус-фактор считается отрицательным). Генотипы записываются так:
- Резус-фактор положителен: Rh + Rh + , Rh + rh – Резус-фактор отрицателен: rh – rh –
В данной задаче сказано, что “дигетерозиготна по. и положительному резус-фактору” – значит, резус фактор будет записывать в генотипе – Rh + rh – .
Обратите внимание, что ошибкой является записать рецессивный ген перед доминантным. За такое могут снять балл на экзамене: aA, bB, i 0 I A, rh – Rh + . Правильный вариант записи: Aa, Bb, I A i 0 , Rh + rh – .
Каждая особь образует 4 гаметы, поэтому потомков получается 16. Подсчитает расщепление по фенотипу:
- 9 потомков : положительный резус-фактор, III (B) группа крови 3 потомка : положительный резус-фактор, I (0) группа крови 3 потомка : отрицательный резус-фактор, III (B) группа крови 1 потомок : отрицательный резус-фактор, I (0) группа крови
Расщепление по фенотипу в данном случае получилось: 9:3:3:1. Здесь проявляется III закон Менделя – закон независимого наследования, так как гены, отвечающие за группу крови и резус-фактор, находятся в разных хромосомах.
Комплементарность
Комплементарность – тип взаимодействия неаллельных генов, при котором развитие признака определяется не одной, а двумя или более парами неаллельных генов, располагающихся в разных хромосомах.
Неаллельные гены – это гены, расположенные в разных локусах хромосом, которые отвечают за разные признаки. В генетике случается такое, что один неаллельный ген может влиять на другой (ген a подавляет действие гена B). В этом разделе статьи мы подробно разберемся с подобным взаимодействием и рассмотрим задачи, которые могут встретиться.
Таким образом, развитие признака определяется именно сочетанием генов друг с другом. Здесь логичнее подчеркнуть совместное действие генов, нежели чем сказать, что доминантный ген подавляет рецессивный – при комплементарности это не совсем так.
В каждой задаче свой случай комплементарного взаимодействия генов. Чтобы успешно их решать, надо помнить, что такое явление, как комплементарность, в принципе, возможно, и быть внимательным при написании генотипов особей и их гамет.
Пример решения задачи №3
Наследование слуха у человека определяется двумя доминантными генами из разных аллельных пар, один из которых детерминирует развитие слухового нерва, а другой – улитки. Определить вероятность рождения глухих детей, если оба родителя глухие, но по разным генетическим причинам (у одного отсутствует слуховой нерв, у другого улитка). По генотипу оба родителя являются дигомозиготными.
Здесь проявляется I закон Менделя – закон единообразия гибридов первого поколения. Возможен только один вариант генотипа ребенка от такого брака. У ребенка будет развит и слуховой нерв, и улитка – ребенок не будет глухим, в отличие от родителей.
Эпистаз
Эпистаз (противоположное действие генов) – явление, при котором один ген аллельной пары (супрессор) в доминантном (доминантный эпистаз) или рецессивном (рецессивный эпистаз) состоянии может подавлять развитие признака, за развитие которого отвечает другая пара генов.
Широко известным примером рецессивного эпистаза является Бомбейский феномен, названный так в результате зафиксированного случая в индийском городе Бомбеи. Доктор Бхенде обнаружил, что у людей рецессивных по гену h (hh) на поверхности эритроцитов не синтезируются агглютиногены – в результате этого они могут быть универсальными донорами.
Говоря проще о Бомбейском феномене: у людей с генотипом hh всегда обнаруживается первая группа крови при любом генотипе – I A I A, I B I B, I A I B. Ген h подавляет гены I A и I B – на поверхности эритроцитов не образуются агглютиногены A и B.
Пример решения задачи №4
“Редкий рецессивный ген (h) в гомозиготном состоянии обладает эпистатическим действием по отношению к генам I A, I B и изменяет их действие до I группы крови (бомбейский феномен). Определите возможные группы крови у детей, если у мужа II гомозиготная, у жены IV и оба родителя гетерозиготны по эпистатическому гену”
Вероятность рождения детей с i(0) группой крови в данном случае равна 2/8, или 1/4 (25%). Генотипами, у которых будет i(0) группа крови являются: I A I A hh и I A I B hh. Эпистатический рецессивный ген hh в гомозиготном состоянии всегда приводит к i(0) группе крови.
Полимерия
Полимерией называют зависимость определенного признака организма от нескольких пар аллельных генов, обладающих схожим действием. Такие гены называются полимерными. Часто выраженность признака зависит от соотношения доминантных и рецессивных аллелей – то есть чем больше доминантных генов, тем более выражен признак.
У человека полимерное действие генов заложено в наследовании количественных признаков (вес, рост, цвет кожи, давление).
Пример решения задачи №5
“Цвет кожи у мулатов наследуется по типу полимерии. При этом данный признак контролируется 2 аутосомными несцепленными генами. Сын белой женщины и негра женился на белой женщине. Может ли этот ребенок быть темнее своего отца?”
В данном случае полимерия проявляется в том, что чем больше доминантных генов в генотипе (A и B), тем более темный цвет кожи имеет человек. Это правило мы и применим для решения.
В результате первого брака (вспоминаем закон единообразия Менделя) получается AaBb – средний мулат. По условиям задачи он берет в жены белую женщину aabb. Очевидно, что в этой семье ребенок не может быть темнее своего отца: дети могут быть средними мулатами (AaBb), как отец, светлыми мулатами (aaBb, Aabb), либо белыми, как мать (aabb).
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Взаимодействие аллельных и неаллельных генов. Явление плейотропии
Аллельны е гены – это гены, расположенные в одинаковых местах (локусах) гомологичных хромосом, отвечающие за развитие альтернативных признаков. Взаимодействие аллельных генов происходит только в гетерозиготном состоянии (Аа).
Варианты взаимодействия аллельных генов:
А) полное доминирование,
Б) неполное доминирование,
Д) плейотропное действие гена.
1. Полное доминирование. Проявляется в тех случаях, когда один аллель
Гена (доминантный) полностью скрывает присутствие другого (рецессивного)
Аллеля. Например:
А – голубые глаза
Человек с генотипом Аа имеет карие глаза.
2. Неполное доминирование. При неполном доминировании фенотип
Гибридов первого поколения (Аа) внешне отличаются от родительских особей
(АА) и (аа). Проявление признака является промежуточным по сравнению с
Родительскими формами.
Например, при скрещивании гомозиготных растений с красными (АА) и белыми (аа) цветками у гибридов первого поколения цветки оказываются розовыми (Аа).
У человека по типу неполного доминирования наследуется признак, определяющий форму волос: ген кучерявых волос (А) неполностью доминирует над геном прямых волос (а), волнистые волосы определяются генотипом – Аа.
3. Кодоминирование – это взаимодействие двух доминантных аллельных генов. Например, каждый из аллельных генов кодирует определенный белок и у гетерозиготного организма синтезируются два вида белка. По типу кодоминирования у человека наследуется четвертая группа крови (I I ).
4. Наддоминир о вание – в гетерозиготном состоянии (Аа) доминантный аллель проявляется в большей степени, чем в гомозиготном (АА). Например, гибриды кукурузы отличаются более высоким ростом, урожайностью зерна по сравнению с гомозиготными растениями Такое явление називается гетерозисом или гибридной силой. У человека но типу наддоминирования проявляется акселерация.
5. Плейотролия – один ген влияет на проявление нескольких признаков, такое явление называется множественным действием одного гена. Например, у человека известная болезнь – синдром Марфана – арахнодактилия («паучьи пальцы») детерминируется доминантным геном, который отвечает за патологическое развитие соединительной ткани, вследствие этого проявляется комплекс патологических признаков – длинные, тонкие («паучьи») пальцы, дефекты развития сердечно-сосудистой системы и подвывих хрусталика (нарушение зрения). В основе таких патологических признаков лежит дефект развития соединительной ткани, обусловленным патологическим геном.
Взаимодействие неаллельных генов
Неаллельные гены – это гены, которые расположены в негомологичных хромосомах. Взаимодействие между неаллельными генами происходит в том случае, если признак проявляется только при их совместном присутствии.
Примерами взаимодействия неаллельных генов могут быть:
1. Комплементарность (или комплементарное взаимодействие генов)
Проявляется тогда, когда действие одного гена дополняется действием другого,
Т. е. для формирования признака необходимо наличие в генотипе двух
Доминантных неаллельных генов.
Например, у душистого горошка красная окраска венчика цветка обусловлена наличием двух доминантных генов А и В (генотип АаВв). В отсутствие одного из них цветки белые (ААвв, ааВВ).
Комплементарное взаимодействие генов у человека проявляется при формировании слуха: развитие нормального слуха обусловлено двумя доминантными неаллельными генами D и Е, из которых Один (D) определяет развитие улитки, а другой (Е)- слухового нерва. Нормальный слух имеют люди с генотипом: DDEE. DdEe, DDEe, DdEE Глухие люди имеют генотипы: Ddee, ddee, ddEE, ddEe.
2. Эпистаз. Явление, противоположное комплементарности. Эпистаз – это
Угнетение одним неаллельным геном (эпистаческим) действия другого
Неаллельного гена (гипостатического). Если эпистатическим действием
Обладает доминантный аллель, говорят о доминантном эпистазе (А > В). При
Рецессивном епистазе такое действие проявляют рецессивные аллели в
Гомозиготном состоянии (а > В). У человека изучен один вид рецессивного
Эпистаза – «бомбейский феномен».

Как известно, группы крови по система АВО обусловлены наследованием трех аллелей одного гена (I A, I B, i). В зависимости от их сочетания формируются четыре группы крови:
III – I B I B l B i
IV – I A 1 B
Однако, существует редкий эпистатический ген ( ) состоянии подавляет все доминантные аллели, определяющие группы крови. Вследствие этого у людей с генотипом – срср, фенотипически проявляется первая только группа крови.
Например, у людей с генотипом А 1 А А блокируется геном-супрессором (р, который проявляет свою активность в гомозиготном рецессивном состоянии ( A i
3. Полимерия – проявление одного признака в зависимости от суммарного действия нескольких неаллельных генов. Причем, чем больше доминантных генов, тем сильнее проявляется признак. Полимерные гены принято обозначать одной буквой латинского алфавита с указанием цифрового индекса (Аь А2).
Примером полимерного действия генов у человека является наследование цвета кожи. Несколько пар (около пяти пар) неалельных доминантных генов, отвечающих за синтез пигмента меланина, который обуславливает темный цвет кожи – А|, А2 и т. д. Генотипы людей с соответствующими оттенками цвегов кожи могут быть:
А|А|А А2 – черная кожа А^АгАг – темная aia, A2 A2 – смуглая а! а|А2а2 – светлая а! а,а2а2 – белая.
Кроме наследования цвета кожи, полимерными генами у человека определяются большинство количественных признаков, таких как рост, масса тела, интеллектуальные особенности, склонность к повышению артериального давления, устойчивость к инфекционным заболеваниям и другие.
Признаки, которые определяются несколькими парами неаллельных генов, называются полигенными.
Задание 1. Трансформируйте активные конструкции в пассивные:
Что заполняет что что скрывает что /присутствие наличие / что угнетает действие чего что подавляет что что блокирует активность чего.
Задание 2. Напишите предложения, раскрывая скобки:
1. Кодоминирование – это явление, когда (гетерозиготное состояние) проявляются признаки обоих генов.
2. Комплементарность проявляется тогда, когда действие одного гена дополняется (действие другого гена).
3. Существует редкий эпистатический ген (ф), который в гомозиготном рецессивном состоянии подавляет (все доминантные аллели групп крови).
4. Активность генов блокируется (эпистатический ген в гомозиготном состоянии).
Что обусловлено чем (тем, что. )
Задание 3. Трансформируйте простые предложения в сложные.
1. Нормальный слух обусловлен присутствием двух неаллельных генов ДиЕ.
2. Окраска венчиков цветка обусловлена наличием двух доминантных генов А и В.
3. Группы крови по системе АВО обусловлены наследованием трех аллелей одного типа (1 А, 1В, Ю).
Задание 4. Прочитайте информацию. Скажите, по каким признакам различаются аллельные и неаллельные гены:
А) аллельные гены – это гены, расположенные в одинаковых местах гомологичных хромосом. Взаимодействие происходит только в гетерозиготном состоянии;
Б) неаллельные гены – это гены, которые расположены в негомологичных хромосомах. Взаимодействие между неаллельными генами происходит в том случае, если они отвечают за развитие одного какого-либо признака.
Задание 5. Прочитайте текст 1 «Взаимодействие аллельных генов» и дайте определение каждого из вариантов взаимодействия аллельных генов.
Задание 6. Прочитайте часть текста «Эпистаз» и ответьте на вопрос: «Что понимают под. »
Б) доминантным эпистазом;
В) «бомбейским феноменом»?
Задание 7. Расскажите о полимерии по следующему плану.
1. Определение полимерии.
2. Зависимость степени проявления признака от количества доминантных генов.
3. Признаки, определяемые полимерными генами.
4. Пример полимерного действия генов.
Решение типовых задач
Взаимодействие аллельных генов (неполное доминирование)
I. У человека прямые волосы – рецессивный признак, а курчавые — неполностью доминируют над прямыми, у гетерозигот проявляются волнистые волосы. Какие дети могут быть у родителей с волнистыми волосами?
Обозначим гены:
А – курчавые волосы
А – прямые волосы
Аа – генотип человека с волнистыми волосами.
Курчавые волнистые прямые волосы волосы волосы
Ответ : 25% детей будут иметь курчавые волосы (ЛА), 25% – прямые волосы (аа) и 50% – волнистые волосы (Аа).
Взаимодействие аллельных генов (кодоминированис)
2. Мужчина со II группой крови (гомозиготный) женился на женщине с III группой крови (гомозигота). Обозначте:
Б) генотипы родителей;
В) напишите схему брака;
Д) определите, какие группы крови могут быть детей;
Е) какое взаимодействие аллельных генов проявилось в данной ситуации?
Обозначим гены, детерминирующие II и III группы крови – 1 А, 1 в. Генотипы родителей:
Взаимодействие аллельных генов в организме — формы и способы, биологическое значение
Главные закономерности наследования признаков у живых организмов были установлены Грегором Менделем. Как известно, все живые существа обладают большим количеством признаков (цвет семян и их форма у растений, тёмный или светлый окрас у животных и так далее), которые определённым образом наследуются от родительских особей. Взаимодействие аллельных генов определяет, какие признаки проявятся у потомка.

Полное доминирование
Одним из главных понятий в генетике является понятие признака — некоторой определённой особенности организма или его части, по которой особь отличается от другой. Признаком в генетическом понимании можно считать любую характеристику на уровне внешних черт, Физиологических или биохимических особенностей:
- вес; цвет; рост; набор веществ определённого типа в органах и клетках; совокупность определённых белков и так далее.

Все признаки одного организма называются фенотипом. Мендель осознал, что все характеристики, которые можно использовать в генетическом анализе, должны выявляться постоянно (независимо от возраста, внешних условий и другого) и должны иметь два или более альтернативных проявлений.
Аллельные гены (аллели) — это гены, отвечающие за один признак, но в разных его проявлениях (например, цвет семян — это один признак, который кодирует один ген, но проявляться может или жёлтый, или зелёный в зависимости от того, какой аллель является доминирующим).
Доминантные и рецессивные аллели одного гена обозначают, как правило, одинаковыми большой и маленькой буквой соответственно. Потомство от скрещивания двух особей с разными альтернативными проявлениями называют гибридным, а отдельного представителя такого потомства — гибридом.
Скрещивание, в котором проводится анализ одной пары альтернативных характеристик, называется моногибридным. Например, цвет семян гороха определяется геном sgr. Нормальный (доминантный) аллель этого гена будет обозначаться как А (он даёт функциональный продукт, который обуславливает разрушение хлорофилла и, соответственно, жёлтый цвет семян), а мутантный рецессивный аллель (ген не даёт функционального продукта, и семена остаются зелёными) как а. Именно поэтому такая мутация, когда теряется способность кодировать полноценный продукт, и обуславливает появление рецессивной формы этого гена.
Неполное доминирование
При полном доминировании и сверхдоминировании проявление признака не зависит от количества (два или один) доминантных аллелей в генотипе. Тем не менее в природе часто встречаются ещё и другие типы взаимодействия аллельных генов. Причина может состоять в том, что рецессивный аллель не даёт функционального продукта, но характеристика зависит от количества копий доминантного аллеля в генотипе. Например, окрас цветов в красный цвет у львиного зева Antirrhinum majus зависит от количества красного пигмента, а он Образуется вследствие активности фермента, который контролируется определённым геном:

- два гена АА обуславливают интенсивный красный окрас; гомозигота аа (отсутствие фермента и, соответственно, пигмента) характеризуется белым окрасом цветков; гетерозигота имеет в два раза меньшую интенсивность окраса в отличие от гомозиготы по доминантному аллелю (в два раза меньше фермента и пигмента) — цветы розового цвета.
Аналогично наследуется кучерявость перьев голубей: особь, гетерозиготная по гену кучерявости, имеет волнистые перья. Такой тип взаимодействия аллельных генов называется неполным доминированием. При неполном доминировании гетерозигота характеризуется фенотипом, отличным от обеих гомозигот — с этой поправкой первый закон Менделя остаётся без изменений. Что касается результатов скрещивания гетерозигот, то расщепление по генотипу и фенотипу будут совпадать и составлять 1:2:1.
Кодоминирование и множественный аллелизм

Аналогичные расщепления характерны и для кодоминирования с той разницей, что потомки от скрещивания гомозигот разных типов будут характеризоваться наличием фенотипических признаков обоих родителей одновременно. То есть в этом случае оба аллеля являются доминантными: образуют функциональные продукты, которые немного отличаются по некоторым характеристикам. Такая ситуация довольно часто наблюдается для ферментов, когда два аллеля обуславливают синтез двух белков — изоферментов — с практически одинаковой активностью, но при этом они отличаются молекулярной массой (один из белков имеет дополнительный структурный домен и так далее).
Эти примеры требуют важного уточнения. Поскольку в двух гомологичных хромосомах могут находиться только два разных аллеля одного гена, ещё не означает, что в группе особей ген обязательно имеет два аллеля. На самом деле, в большинстве случаев гены существуют в виде нескольких (теоретически неограниченное количество) разных аллелей. Это явление получило название множественный аллелизм.
Одним из примеров множественного аллелизма является система групп крови АВ0. Хорошо известны четыре группы крови — 0, А, В и АВ — определяются тремя аллелями одного гена: ІА, ІВ, і0. Аллели ІА и ІВ являются кодоминантными (отвечают за формирование на поверхности эритроцитов антигенов двух типов — А и В), но при этом доминируют над аллелем і0, который не продуцирует ни одного антигена.
При наличии трёх аллелей возможно шесть комбинаций между ними, которые можно получить, используя таблицу Пеннета. Но рецессивность і0 сводит количество фенотипов (групп крови) к четырём: группе 0 соответствует генотип і0і0, группе А — генотипы ІАІА и ІАі0, группе В — ІВІВ и ІВі0, группе АВ — генотип ІАІВ.
Причины отклонений от менделевских расщеплений
Менделевские расщепления в принципе могут быть реализованы только для видов живых организмов с нормальным половым процессом, диплоидностью обоих полов и при условии отсутствия нарушений в мейозе. Но и при выполнении указанных условий случаи отклонения от менделевских расщеплений достаточно многочисленны. При этом каких-либо исключений нет, а условия менделирования не нарушаются — Просто на них накладываются другие эффекты.

Три таких эффекта — взаимодействие неаллельных генов (комплементация, кумулятивная и некумулятивная полимерия), сцепление групп генов в одной хромосоме и расположение гена в половых хромосомах. Среди других причин выделяются такие:


На самом деле, в биологии все эти эффекты могут быть сведены к взаимодействию генов в сложной системе функционирования генома, хотя не всегда такое взаимодействие можно легко описать: часто нужно брать во внимание слишком много отдельных элементов, которые влияют один на другого.
Взаимодействие аллельных генов в организме – формы и способы, биологическое значение
Часто развитие признаков обусловлено сложным взаимодействием между несколькими генами. Понятие «взаимодействие генов» появилось задолго до выяснения механизма функционирования генов. На самом деле взаимодействуют не гены, а их продукты — белки-ферменты, обусловливающие развитие у организмов того или иного признака.
Взаимодействия аллельных генов
- Полное доминирование (доминантность) — полное преобладание в фенотипе гетерозиготного организма одного аллеля (доминантного) над другим (рецессивным) одного и того же гена. Неполное доминирование — у гетерозиготного организма доминантный аллель не до конца подавляет рецессивный того же гена. При неполном доминировании фенотип гетерозиготы Аа будет промежуточным между фенотипом доминантной АА и рецессивной аа гомозигот. Сверхдоминирование — доминантный аллель в гетерозиготном состоянии проявляется в фенотипе сильнее, чем в гомозиготном (Аа > АА). При этом типе имеет место действие летальных генов (серповидноклеточная анемия в условиях заражения малярией: здоровая особь АА гибнет от малярии, особь Аа — рецессивный аллель а (серповидноклеточная анемия) защищает организм от заболевания малярией, гомозиготы аа умирают от анемии). Кодоминирование — в фенотипе гетерозиготного организма проявляются оба аллеля одного и того же гена. Четвёртая группа крови человека (генотип I А I В ): в эритроцитах крови одновременно имеются антиген А (I А I А или I А I 0 ), характерный для II группы крови, и антиген В (аллель I B I B или I B I 0 ), свойственный III группе крови. Их отсутствие (I 0 I 0 ) в крови человека определяет формирование I группы крови. Аллели I А и I B — кодоминантные.
Кодоминирование и неполное доминирование фенотипически схожи, но имеют различные механизмы проявления. Кодоминирование — полноценное проявление двух аллелей; неполное доминирование — доминантный аллель не полностью подавляет рецессивный, генотипы различаются степенью выраженности признака.
Взаимодействие неаллельных генов
Эпистаз — взаимодействие неаллельных генов, при котором один ген подавляет действие гена из другой (неаллельной) пары. Подавляющий ген называется Эпистатическим, или Супрессором (ингибитором), подавляемый — Гипостатическим. Если ингибитор доминантный, эпистаз называют доминантным (А > В). Если ингибитор рецессивный, эпистаз — рецессивный (аа > В_ или аа > bb).
Плейотропия — явление множественного действия гена. Выражается в способности одного гена влиять на несколько фенотипических признаков. Один белок-фермент, образующийся под контролем одного гена, определяет развитие не только данного признака, но и воздействует на вторичные реакции биосинтеза других признаков и свойств, вызывая их изменения (синдром Марфана).
Полимерия — взаимодействие неаллельных генов, при котором один и тот же признак контролируется несколькими доминантными неаллельными генами, действующими на этот признак однозначно и в равной степени, усиливая его проявление. Такие однозначные гены называют Полигенами (полимерными) и обозначают одной буквой латинского алфавита, но с разными цифровыми индексами. Например, доминантные полимерные гены — A1, А2, A3 и т. д., рецессивные — а1, а2, а3 и т. д. Различают два вида полимерии: Кумулятивная (наследование, при котором с увеличением дозы доминантных генов усиливается степень выраженности признака, — цвет кожи) и Некумулятивная (степень выраженности признака не зависит от количества доминантных генов с однозначным фенотипическим проявлением, признак развивается в полной мере при наличии хотя бы одного гена).
Комплементарность — взаимодействие неаллельных генов, в результате которого проявляется новый признак, не определявшийся ни одним, ни другим геном.
Типы комплементарности
- Развитие нового признака организма определяется доминантными юнами из различных пар хромосом, каждый из которых не имеет самостоятельного фенотипического проявления. Один из доминантных генов способен к самостоятельному фенотипическому проявлению, Доминантные гены из разных пар хромосом, каждый из которых имеет собственное фенотипическое проявление.
ГЕНЕТИКА ЧЕЛОВЕКА
Законы наследственности применимы не только к растениям и животным, но и к человеку. От родителей ребенок получает набор генов, определяющих развитие всех признаков. У человека, как и у других организмов, есть доминантные и рецессивные признаки. Ф. Гальтон впервые выделил наследственность человека как самостоятельный предмет исследования в 1865 г.
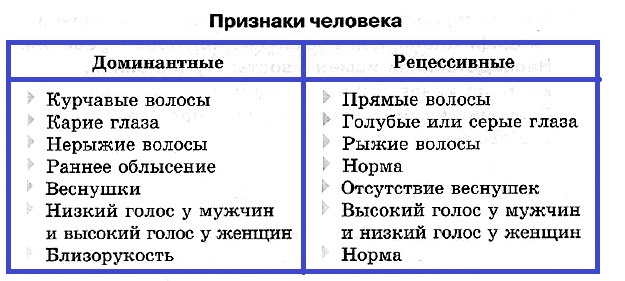
Генетика человека — раздел общей генетики, изучающий особенности наследования признаков у человека, наследственные заболевания (медицинская генетика), генетическую структуру популяций человека. Генетика человека является теоретической основой современной медицины и современного здравоохранения.
Главная цель: разработка путей сохранения, продления и оздоровления жизни человека, а также вы явление его истинных способностей.
Методы изучения генетики человека:
- генеалогический; близнецовый; цитогенетический; биохимический; популяционно-статистический; метод дерматоглифики; молекулярно-генетический; гибридизация ДНК; метод соматической гибридизации.
Генеалогический метод — изучение родословной человека для выявления распространения какого-либо признака или наследственного заболевания, установление наследственного характера определенных способностей человека.
Цитогенетический метод — исследование структуры хромосом. Хромосомы, окрашенные специальными красителями, рассматривают под микроскопом. Используют для изучения наследственных заболеваний, связанных с нарушениями структуры хромосом.
Близнецовый метод — сравнение характера проявления одних и тех же признаков у близнецов. Используется для выявления роли наследственности и среды в развитии тех или иных признаков.
Биохимический метод — выявление нарушений обмена веществ при некоторых наследственных заболеваниях.
Онтогенетический метод — используется для выяснения механизма развития наследственных заболеваний в онтогенезе, что важно для их лечения и профилактики.
Популяционный метод — изучение распространения отдельных генов или хромосомных аномалий в человеческих популяциях.
Особенности изучения генетики человека
- Невозможность произвольного скрещивания. Позднее наступление половой зрелости. Небольшое число детей у большинства супружеских пар. Невозможность уравнивания условий жизни для потомства.
Генотип как целостная система
Генотип — целостная система взаимодействующих генов организма. Гены могут наследоваться независимо, либо находиться в тесном взаимодействии друг с другом. Целостность генотипа сформировалась в процессе эволюции вида. Признаком системности генотипа является Взаимодействие генов.
Термин генотип предложен в 1909 г. датским генетиком Вильгельмом Иогансеном. Он же ввел термины: ген, аллель, фенотип, линия, чистая линия, популяция.
Это конспект для 10-11 классов по теме «Взаимодействие генов. Генетика человека». Выберите дальнейшее действие:
Что такое аллельные гены: свойства, способы взаимодействия и множественный аллелизм.
Что такое аллельные гены?
Аллельные гены – это гены, которые занимают одинаковое положение в локусах гомологичных хромосом и отвечают за развитие одного и того же признака.
Какие свойства есть у аллельных генов
Если рассмотреть генотип любого живого организма, то можно обнаружить, что он состоит из большого числа различных генов. Все вместе они образуют органическую совокупность и, являясь одним целым, выполняют общие функции.
Г. Менделем, которого считают основателем генетики, описана лишь одна возможность взаимодействия аллельных генов: когда одна полностью доминирует над другой. Аллель, которая подавляется, называют рецессивной.
Менделя считают отцом генетики как науки еще и потому, что он сформулировал все возможные закономерности наследования признаков. Сделал он это с помощью генетического метода, который и сегодня является наиболее перспективным. В основе метода лежит скрещивание организмов с определенными признаками и анализ проявлений этих признаков у потомства.
В этом месте нужно уточнить, что гены не всегда могут проявляться в виде признаков: при одинаковом генотипе у организмов могут быть фенотипические различия. Это объясняется тем, что на фенотип оказывает влияние то, как взаимодействуют генотип и окружающая среда.
Кроме того, фенотипическое проявление генов обусловлено не только одной парой генов: как минимум потому, что оно является результатом взаимодействия генотипической системы в целом.
Аллельное генотипическое взаимодействие — контакты белков и ферментов, а не генов.
Если принять и понять этот принцип, то можно избежать ошибок, проводя генетические исследования взаимодействия аллельных генов.
У методов Менделя есть определенные Преимущества:
- Обеспечение точного количественного учета проявления признаков. В качестве доказательства сам ученый приводил математические расчеты, что уже тогда позволило назвать такую работу синтезом биологии и математики; Обращение внимания на каждый признак в отдельности. Исследуя аллельный ген, и сегодня придерживаются этого пункта; Использование чистых линий растений. Случайные растения не рассматривались.
Чистая линия — это совокупность организмов, которые при длительном, на протяжении нескольких поколений, скрещивании друг с другом проявляют одинаковые признаки (расщепление отсутствует).
Ученый сформулировал 3 закона наследственности:
Первый закон — правило единообразия. Второй закон — закон расщепления признаков. Третий закон — закон о независимости наследования признаков.
Эти законы позволяют описывать различные закономерности изменчивости и наследственности. Принципы, лежащие в основе этих законов, применяются в биологии и сегодня.
Способы взаимодействия аллельных генов
Согласно основам генетики, есть 2 варианта генотипического взаимодействия:
Взаимодействие аллельных генов. Неаллельное взаимодействие.
Исходя из этого, все живые организмы обладают парными аллельными генами. Внутри организма гены взаимодействуют 3 различными способами:
- кодоминирование; сверхдоминирование; полное и неполное доминирование.
Кодоминирование
В случае такого взаимодействия аллельные гены проявляют свое действие независимо друг от друга.
Для варианта кодоминирования аллельных генов пример — это система групп крови ABO. Здесь гены A и B функционируют независимо.
Сверхдоминирование
В этой ситуации качество фенотипический проявлений доминантного гена увеличивается только тогда, когда он тесно связан с рецессивным.
В случае если в одной аллели находится два доминантных гена, то, как правило, их действие и проявление сильно хуже, чем в предыдущем варианте с одним доминантным и одним рецессивным геном.
Полное и неполное доминирование
При полном перекрытии доминантным геном рецессивного говорят о полном доминировании.
Неполное доминирование — вариант взаимодействия генов, когда рецессивный ген не подавляется полностью и может оказывать влияние (хотя бы минимальное) на фенотипическое проявление признака. В таком случае фенотипическое проявление признака является промежуточным — между родительскими формами.
Пример неполного доминирования — наследование окраски венчика цветка ночной красавицы. Здесь родительские формы имеют белый и красный цвета, а промежуточным будет розовый.
Множественный аллелизм
В генетике встречается такое явление как множественный аллелизм. В каждом организме есть два аллельных гена, при этом самих аллелей может быть больше двух. При таком раскладе только одна пара аллелей может проявлять фенотипические признаки: другие гены не задействуются. Гомологичные аллели, то есть одинаковые, «работают» над развитием одного и того же признака. При этом качество его проявления будет различаться.
При множественном аллелизме формы взаимодействия генов могут быть различными. Даже несмотря на то, что они отвечают за один и тот же признак. Дело в том, что проявляют они этот признак по-разному и при помощи различных способов (описанных выше).
Самый простой пример — окраска шерсти кролика. Здесь могут быть следующие варианты: белая, гималайская, шиншилловая, черная и коричневая. И это при том, что есть целая серия разных аллелей генов, ответственных за окрас. И таких примеров в биологии достаточно.
Несмотря на всю парадоксальность множественного аллелизма, в половую клетку живого существа проникает только одна пара гомологичных аллелей, и какая именно — вопрос случая. Так обеспечивается изменчивость каждого отдельного вида, играющая важнейшую роль в эволюции.
Благодаря изучению аллельных генов, становятся понятными закономерности наследования признаков. А еще это помогает исключить негативные последствия изменения наследственного набора организма.
29. Взаимодействие аллельных генов
Взаимодействие между аллельными генами осуществляется в виде трех форм: полное доминирование, неполное доминирование и независимое проявление (кодоминирование).
Полное доминирование – когда один доминантный аллель полностью подавляет проявление рецессивного аллеля, например, желтая окраска горошин доминирует над зеленой.
Неполное доминирование наблюдается в том случае, когда один ген из пары аллелей не обеспечивает образование в достаточном для нормального проявления признака его белкового продукта. При этой форме взаимодействия генов все гетерозиготы и гомозиготы значительно отличаются по фенотипу друг от друга. Примером расщепления при неполном доминировании может служить наследование окраски цветков Ночной красавицы.
При скрещивании растений с красными цветками (АА) и растений с белыми (аа) гибриды F1 имеют розовые цветки (Аа). Таким образом, имеет место неполное доминирование; в F2 наблюдается расщепление 1 : 2 : 1 как по фенотипу, так и по генотипу.
Кроме полного и неполного доминирования известны случаи отсутствия доминантно-рецессивных отношений или кодоминирования. При кодоминировании у гетерозиготных организмов каждый из аллельных генов вызывает формирование в фенотипе контролируемого им признака.
Примером этой формы взаимодействия аллелей служит наследование групп крови человека по системе АВ0, детерминируемых геном I. Существует три аллеля этого гена Io, Ia, Ib, определяющие антигены групп крови. Наследование групп крови иллюстрирует также явление множественного аллелизма: в генофондах популяций человека ген I существует в виде трех разных аллелей, которые комбинируются у отдельных индивидуумов только попарно. До этого примера мы говорили о генах, существующих только в двух разных аллельных формах. Однако многие гены состоят из сотен пар нуклеотидов, так что мутации могут проходить во многих участках гена и порождать множество различных его аллельных форм. Так как в каждой из гомологичной хромосом имеется по одному аллельному гену, то, разумеется, диплоидный организм имеет не более двух из серии аллелей генофонда популяции.
30. Неаллельные взаимодействия генов
Неаллельные гены— это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой.
При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы и взаимодействия неаллельных генов:
Комплементарное(дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9:6:1, 9:3:4, 9:7, иногда 9:3:3:1. Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлинённую. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдёт расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлинённые.
Эпистаз— взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3, 7:6:3. Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена аллелей гипостатичного гена (i > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7, 13:3.
Полимерия — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление F2 но фенотипу происходит в соотношении 1:4:6:4:1.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1.
Пример: цвет кожи у людей, который зависит от четырёх генов.
Биология в лицее
Сайт учителей биологии МБОУ Лицей № 2 г. Воронежа, РФ
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
Ген является структурной единицей наследственной информации. Материально ген представлен участком молекулы ДНК (в редких случаях – РНК). Гены контролируют элементарные признаки в процессе индивидуального развития организма. Первые исследования природы гена, проводившиеся в начале XX в., в основном были направлены на выяснение роли гена в передаче наследственных признаков. Не менее важной задачей стала расшифровка закономерностей действия генов. Ее решение имеет не только теоретическое, но и практическое значение, поскольку позволит предотвратить возможные вредные последствия этого действия.
Исследования генетиков установили дискретный характер генов, что подтверждается их независимым друг от друга наследованием: каждый из генов определяет развитие какого–то признака, независимого от других. Однако между различными генами существуют разные типы взаимодействия, обусловленные сложными отношениями как между аллельными, так и между неаллельными генами. Объединяясь в генотипе, они все вместе выступают как система взаимодействующих между собой генов особи.

Среди взаимодействий генов следует назвать прежде всего отношения доминантности и рецессивности, когда рецессивная аллель гена под влиянием доминантной аллели не проявляется в фенотипе. Кроме того, имеются факты, показывающие, что гены влияют на проявление действий других, неаллельных генов. Описаны также случаи, когда развитие того или иного признака организма находится под контролем не одного, а многих генов. Например, у человека не менее четырех генов определяют различие в цвете кожи представителей негроидной и европеоидной рас.
Взаимодействие генов
| Взаимодействие Аллельных генов |
Полное доминирование – одна аллель гена (рецессивная) под влиянием доминантной аллели не проявляется в фенотипе |
| Неполное доминирование наблюдается промежуточный характер наследования признаков у гетерозиготной особи с большим или меньшим отклонением в сторону доминантного или рецессивного состояния | |
| Кодоминирование – совместное участие обоих аллелей в определении признака у гетерозиготной особи | |
| Взаимодействие Неаллельных генов |
Комплементарное взаимодействие – доминантные или рецессивные аллели обусловливают развитие нового, отличного от родительских вариантов, признака |
| Множественное действие генов – действие одного гена влияет на развитие нескольких признаков | |
| Полимерия – степень развития одного и того же признака обусловлена влиянием целого ряда генов | |
| Эпистаз – аллели одного гена подавляют проявление аллелей других генов |

При скрещивании петуха с розовидным гребнем с курицей, имеющей гороховидный гребень, в F1 следует ожидать доминирования одного из признаков, а в F2 — расщепления 3:1.
В действительности результаты скрещивания оказываются резко отличными от ожидаемых. В F 1 все птицы имеют гребни, не похожие ни на одного из родителей, — эти гребни называются ореховидными. В F 2 , полученном от скрещивания петухов и кур с ореховидными гребнями, обнаруживаются четыре фенотипических класса: два с проявлением родительских признаков (розовидный и гороховидный гребни) — по 3 части от всего потомства, 9 частей с ореховидным гребнем и 1 часть с новым типом гребня (простым, или листовидным).
Наблюдаемое расщепление по фенотипу 9:3:3:1 ясно указывает, что имеет место дигибридное скрещивание. Для объяснения необычного характера наследования формы гребня была предложена гипотеза, согласно которой неаллельные гены R и Р , определяющие развитие гребня определенной формы, взаимодействуют друг с другом, причем результаты взаимодействия зависят от того, в какой форме находится каждый из генов — в доминантной или в рецессивной.
R — розовидный гребень, R — отсутствие розовидного гребня, P — гороховидный гребень, P — отсутствие гороховидного гребня. Если в обеих парах генов присутствуют доминантные аллели R_P_, то возникает новая форма гребня — ореховидный гребень, при генотипе Rrpp у кур развивается листовидный гребень.
Комплементарным называется взаимодействие, при котором действие генов из одной пары дополняется действием генов из другой пары таким образом, что в результате появляется новый признак.
Отличительная особенность комплементарного взаимодействия от закона независимого наследования признаков заключается в количестве учтённых признаков: в третьем законе рассматривается комбинирование двух признаков, а в комплементарном взаимодействии четыре вариации одного признака.

Рецессивный эпистаз. Скрещивание лука с красными и белыми луковицами даёт в F 1 растения только с красными луковицами, а в F 2 — расщепление: 9/16 — с красными, 3/16 — с жёлтыми и 4/16 — с белыми луковицами.
Обозначим ген, определяющий красный цвет луковиц, через Y ; ген, определяющий жёлтый цвет, — через y . Рецессивный ген с (от англ. colorless — бесцветный) в гомозиготном состоянии определяет образование белых луковиц, а доминантный С — окрашенных.
Взаимодействие генов, при котором один ген подавляет действие другого, называется эпистазом (от греч. эпи — над и стазис — стоять).
Если эпистатический ген проявляется в рецессивной форме, то такое взаимодействие называют рецессивным эпистазом. Расщепление по фенотипу при такой форме взаимодействия генов 9:3:4.

Доминантный эпистаз. В некоторых случаях наличие одного доминантного гена подавляет проявление другого неаллельного гена. Например, ген Y обуславливает жёлтый цвет плодов тыквы, а его рецессивная аллель Y — зелёный цвет плодов.
При наличии в генотипе доминантной аллели особого гена W действие гена Y (или Y) не проявляется и плоды имеют серовато-белую окраску. При генотипе wwY_ плоды тыквы имеют жёлтый цвет. Наконец, если оба гена представлены рецессивными аллелями ( wwyy ), плоды тыквы имеют зелёный цвет. При скрещивании тыкв с белыми ( WWYY ) и зелёными ( wwyy ) плодами все гибриды первого поколения будут иметь белые плоды ( WwYy ), а во втором поколении наблюдается расщепление по фенотипу 12 (белая кожура): 2 (жёлтая кожура): 1 (зелёная кожура).
Если эпистатический ген проявляется в доминантной форме, то такое взаимодействие называют доминантным эпистазом.

Полимерия. У пшеницы красный цвет зёрен определяется двумя генами: А1 и А2. Неаллельные гены обозначаются одной буквой алфавита потому, что определяют развитие одного признака. При генотипе А1А1А2А2 окраска зёрен наиболее интенсивная. При генотипе А1а1а2а2 зёрна имеют белый цвет. В зависимости от числа доминантных генов в генотипе можно получить все переходы между интенсивно красной и белой окраской.
Такая обусловленность определённого признака несколькими парами неаллельных генов, обладающих одинаковым действием, носит название полимерии (от греч. поли — много и мерос — часть).
При кумулятивной (от лат. кумуло — собираю, накапливаю), или накопительной, полимерии, как в случае с окраской зёрен пшеницы, степень проявления признака зависит от суммирующего проявления генов.

У человека интенсивность окраски кожи определяется несколькими парами генов. Учёные предполагают, что цвет кожи зависит от трёх пар генов. Самая тёмная кожа (у представителей негроидной расы) будет определяться генотипом А1А1А2А2А3А3; у мулатов будут присутствовать несколько рецессивных аллелей, а у коренного населения Европы генотип не будет иметь доминантных генов — А1а1а2а2а3а3.
В зависимости от числа доминантных генов в генотипе может формироваться более светлый или более тёмный цвет кожи.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1 для двух генов, 64:1 для трёх генов и т. д. Например, цвет кожи у людей зависит от четырёх генов, поэтому от брака двух мулатов практически никогда не рождаются дети с белой кожей.
Но иногда гаметогенез у отдалённых гибридов протекает нормально, что позволило получить новые ценные породы животных. Примером являются архаромериносы, которые, как и архары (горные бараны), могут пастись высоко в горах, а как мериносы дают хорошую шерсть. Получены плодовитые гибриды от скрещивания местного (индийского) крупного рогатого скота с зебу. При скрещивании белуги и стерляди получен плодовитый гибрид — бестер, хорька и норки — хонорик, продуктивен гибрид между карпом и карасём.
ОТЛИЧИЕ КУЛЬТУРНЫХ ФОРМ ОТ ДИКИХ
Наследование окраски шерсти у собак пример доминантного эпистаза.
3dnauka. ru
24.09.2020 18:23:25
2020-09-24 18:23:25
Источники:
Http://buzani. ru/zadachi/genetika/787-vzaimodejstvie-neallelnykh-genov-komlementarnost-z
Http://sites. google. com/site/biologiaege/neallelnye-geny
Http://3dnauka. ru/biologiya/vzaimodejstvie-allelnyh-genov-v-organizme-formy-i-sposoby-biologicheskoe-znachenie
Взаимодействие неаллельных генов
05-Июл-2019 | Нет комментариев | Лолита Окольнова
Неаллельные гены — это гены, расположенные в различных участках хромосом и кодирующие разные белки (признаки)
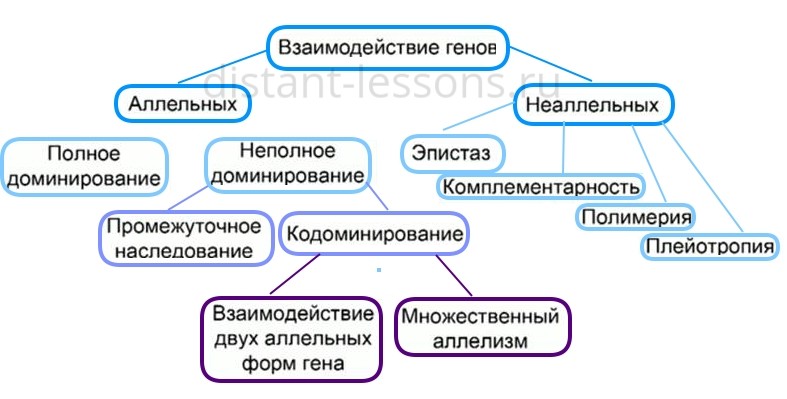
Эпистаз генов — аллели одного гена подавляют проявление аллелей другого гена или генов, при этом эпистазировать могут как доминантные, так и рецессивные аллели.
Рассмотрим на примере кур леггорнов:
- А — белое оперение
- а — цветное;
- В — черное оперение
- в — коричневое
При этом белая окраска подавляет проявление черной
P: АаВв х АаВв
G: АВ Ав аВ ав АВ Ав аВ ав
F1:
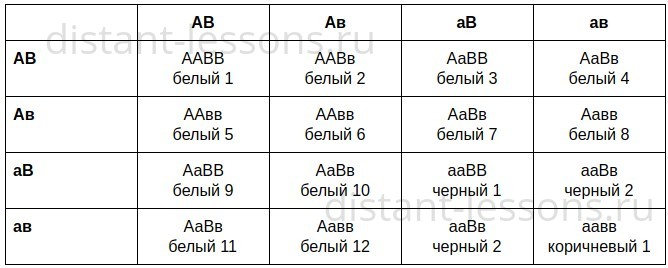
Получаем расщепление по фенотипу: 12:3:1
Полимерия — признак один, но неаллельных генов несколько. Степень проявления признака зависит от количества полимерных генов, действие таких генов суммируется, а фенотипическое проявление признака тем сильнее, чем больше генов участвует в его контролировании.
Тут возможны 2 варианта:
Кумулятивная (накопительная) полимерия — степень проявления признака зависит от числа доминантных генов, чем их больше, тем сильнее проявляется признак.
Количество пигмента меланина в коже человека определяется доминантными (A1A1A2A2) и рецессивными (а1а1а2а2) генами.
Интенсивность окраски кожи зависит от соотношения доминантных и рецессивных полимерных генов:
- негроидная раса — доминантный гомозиготный — A1A1A2A2
- европеоидная — рецессивный гомозиготный — а1а1а2а2
- мулат — дигетерозиготный — A1а1A2а2
- светлокожий негр — гетерозиготный по одному гену — A1A1A2а2
- светлокожий мулат -гетерозиготный по одному гену- A1а1а2а2
У детей мулатов возможны все варианты окраски кожи — от черной до белой включительно
Попробуем решить задачу:
Сын белой женщины и чернокожего мужчины женился на белокожей женщине. Может ли сын, рожденный от такого брака, оказаться темнее своего отца?
- отец по условию задачи будет мулатом (гетерозиготным): A1а1A2а2
- белокожая (рецессивный гомозиготный организм) мать: а1а1а2а2
Р: A1а1A2а2 х а1а1а2а2
G: A1A2 A1а2 а1A2 а1а2 а1а2
Чтобы сын был темнее, он должен быть доминантным гомозиготным ( негроидная раса), а такого варианта сочетания гамет у нас нет.
Некумулятивная полимерия — признак будет проявляться, если есть хотя бы один доминантный ген
Плейотропия — взаимодействие, при котором один ген контролирует развитие нескольких признаков, то есть один ген отвечает за формирование фермента, который влияет не только на свою реакцию, но и оказывает влияние на вторичные реакции биосинтеза
Пример — синдром Морфана — доминантный ген, вызывающий дефекты развития соединительной ткани, а следовательно, дефекты развития всех ее производных — вывих хрусталика, длинные вытянутые пальцы, проблемы с суставами, пороки сосудов и клапанов сердца.
Комплементарность — два неаллельных гена приводят к формированию нового (не промежуточного!) фенотипического признака

- примеры вопросов по теме
Обсуждение: «Взаимодействие неаллельных генов»
(Правила комментирования)
Так как в первом поколении все потомки одинаковы, родители гомозиготны. Появление во втором поколении третьей окраски и численные соотношения указывают на то, что это дигибридное скрещивание. Значит, в потомстве имеется 16 генотипических классов. Общее число потомков второго поколения = 115 + 79 + 13 = 207. Размер генотипического класса = 207 : 16 = 13. С оранжевой окраской 115 : 13 = 9 генотипических классов. С красной окраской 79 : 13 = 6 генотипических классов. С жёлтой окраской 13 : 13 + 1 генотипический класс. Таким образом, во втором поколении расщепление 9:6:1, что соответствует комплементарному взаимодействию генов. Родительские формы ААbbи ааВВ, гибриды первого поколения АаВb. Во втором поколении 9 А__В_ : 6 (ааВ_ + А_bb) :1 ааbb.
Селекция — наука, разрабатывающая пути создания новых и улучшения существующих сортов растений, пород животных и штаммов микроорганизмов.
Создание новых сортов и пород основывается на таких важнейших свойствах живого организма, как наследственность и изменчивость. Именно поэтому генетика — наука об изменчивости и наследственности организмов — является теоретической основой селекции.
Имея свои собственные задачи и методы, селекция твердо опирается на законы генетики, является важной областью практического использования закономерностей, установленных генетикой. Вместе с тем селекция опирается и на достижения других наук. На сегодняшний день генетика вышла на уровень целенаправленного конструирования организмов с нужными признаками и свойствами.
Сорт, порода и штамм — устойчивая группа организмов, искусственно созданная человеком и имеющая определенные наследственные особенности.
Все особи внутри породы, сорта и штамма имеют сходные, наследственно закрепленные морфологические, физиолого-биохимические и хозяйственные признаки и свойства, а также однотипную реакцию на факторы внешней среды.
Основные направления селекции:
- высокая урожайность сортов растений, плодовитость и продуктивность пород животных;
- улучшение качества продукции (например, вкус, внешний вид плодов и овощей, химический состав зерна — содержание белка, клейковины, незаменимых аминокислот и т. д.);
- физиологические свойства (скороспелость, засухоустойчивость, зимостойкость, устойчивость к болезням, вредителям и неблагоприятным климатическим условиям).
- выведение стрессоустойчивых пород (для разведения в условиях большой скученности — на птицефабриках, фермах и т. п.);
- пушное звероводство;
- рыбоводство — разведение рыбы в искусственных водоемах.
ОТЛИЧИЕ КУЛЬТУРНЫХ ФОРМ ОТ ДИКИХ
| Культурные формы | Дикие формы |
| развиты признаки, полезные для человека и часто вредные в естественных условиях | наличие признаков, неудобных для человека (агрессивность, колючесть и т. п.) |
| высокая продуктивность | низкая продуктивность (мелкие плоды; низкая масса, яйценоскость, удойность) |
| хуже адаптируются к меняющимся условиям среды | высокая адаптивность |
| не имеют средств защиты от хищников и вредителей (горьких или ядовитых веществ, шипов, колючек и т. п.) | наличие естественных защитных приспособлений , повышающих жизнестойкость, но неудобных для человека |
основные методы селекции
Основные методы селекции:
- подбор родительских пар
- отбор
- гибридизация
- искусственный мутагенез
ПОДБОР РОДИТЕЛЬСКИХ ПАР
Данный метод применяется прежде всего в селекции животных, т. к. для животных характерно половое размножение и немногочисленное потомство.
Выведение новой породы — процесс длительный, требующий больших материальных затрат. Это может быть целенаправленное получение определенного экстерьера(совокупности фенотипических признаков), повышение молочности, жирности молока, качества мяса и т. д.
Разводимые животные оцениваются не только по внешним признакам, но и по происхождению и качеству потомства. Поэтому необходимо хорошо знать их родословную. В племенных хозяйствах при подборе производителей всегда ведется учёт родословных, в которых оцениваются экстерьерные особенности и продуктивность родительских форм в течение ряда поколений.
РАБОТЫ И. В. МИЧУРИНА
ОТБОР
Искусственный отбор — сохранение для дальнейшего размножения особей с интересующими селекционера признаками. Формы отбора: массовый и индивидуальный.
- Интуитивный (бессознательный) отбор — самая древняя форма отбора, используемая ещё древним человеком: отбор особей по фенотипу, т.е. с наиболее полезными сочетаниями признаков.
- Методический отбор — отбор для размножения особей с чётко определёнными признаками, согласно цели и с учетом их фенотипов и генотипов.
- Массовый отбор — устранение из размножения особей, не имеющих ценные признаки, либо имеющих нежелательные признаки (например, агрессивных).
Массовый отбор может быть эффективен в том случае, если отбираются качественные, просто наследуемые и легко определяемые признаки. Массовый отбор обычно проводят среди перекрестноопыляемых растений. При этом селекционеры отбирают растения по фенотипу с интересующими их признаками. Недостаток массового отбора заключается в том, что селекционер не всегда может определить лучший генотип по фенотипу.
- Индивидуальный отбор — выделение отдельных особей с интересующими человека признаками и получение от них потомства.
Индивидуальный отбор более эффективен при отборе особей по количественным, сложно наследуемым признакам. Этот вид отбора позволяет точно оценить генотип благодаря анализу наследования признаков у потомства. Индивидуальный отбор применяют по отношению к самоопыляемым растениям (сорта пшеницы, ячменя, гороха и др.).
ГИБРИДИЗАЦИЯ
В селекционной работе с животными применяют в основном два способа скрещивания:инбридинг и аутбридинг.
Инбридинг — скрещивание близкородственных форм: в качестве исходных форм используются братья и сестры или родители и потомство.
Результат: получение гомозиготных организмов → разложение исходной формы на ряд чистых линий.
Минусы: пониженная жизнеспособность (рецессивные гомозиготы зачастую несут наследственные заболевания).
Такое скрещивание в определённой степени аналогично самоопылению у растений, которое также приводит к повышению гомозиготности и, как следствие, к закреплению хозяйственно ценных признаков у потомков. При этом гомозиготизация по генам, контролирующим изучаемый признак, происходит тем быстрее, чем более близкородственное скрещивание используют при инбридинге. Однако гомозиготизация при инбридинге, как и в случае растений, ведет к ослаблению животных, снижает их устойчивость к воздействию среды, повышает заболеваемость.
В селекции инбридинг обычно является лишь одним из этапов улучшения породы. За ним следует скрещивание разных межлинейных гибридов, в результате которого нежелательные рецессивные аллели переводятся в гетерозиготное состояние и вредные последствия близкородственного скрещивания заметно снижаются.
Аутбридинг — неродственное скрещивание между особями одной породы или разных пород животных в пределах одного вида.
Результат: получение большого количества гетерозиготных организмов → поддержание полезных качеств и усиление их выраженности в ряду следующих поколений.
Отдалённая гибридизация — получение межвидовых и межродовых гибридов.
Отдалённая гибридизация в селекции животных применяется значительно реже, чем в селекции растений.
Межвидовые и межродовые гибриды животных и растений чаще всего бесплодны, так как нарушается мейоз и гаметогенез не происходит. При этом восстановление плодовитости у животных представляет более сложную задачу, поскольку получение полиплоидов на основе умножения числа хромосом у них невозможно.
Преодоление бесплодия межвидовых гибридов растений впервые удалось осуществить в начале 20-х годов ХХ века советскому генетику Г. Д. Карпеченко при скрещивании редьки и капусты. Это вновь созданное человеком растение не было похоже ни на редьку, ни на капусту. Стручки занимали как бы промежуточное положение и состояли из двух половинок, из которых одна напоминала стручок капусты, другая — редьки. Каждая из исходных форм имела в половых клетках по 9 хромосом. В этом случае клетки полученного от них гибрида имели 18 хромосом. Но некоторые яйцеклетки и пыльцевые зёрна содержали все 18 хромосом (диплоиды), а при их скрещивании создано растение с 36 хромосомами, которое оказалось плодовитым. Так была доказана возможность использования полиплоида для преодоления нескрещиваемости и бесплодия при отдалённой гибридизации.
Бывает, что бесплодны особи только одного пола. Например, у гибридов высокогорного быка яка и рогатого скота бесплодны (стерильны) самцы, а самки плодовиты(фертильны).
Но иногда гаметогенез у отдалённых гибридов протекает нормально, что позволило получить новые ценные породы животных. Примером являются архаромериносы, которые, как и архары (горные бараны), могут пастись высоко в горах, а как мериносы дают хорошую шерсть. Получены плодовитые гибриды от скрещивания местного (индийского) крупного рогатого скота с зебу. При скрещивании белуги и стерляди получен плодовитый гибрид — бестер, хорька и норки — хонорик, продуктивен гибрид между карпом и карасём.
В природе встречаются гибриды зебры и лошади (зеброид), бизона и зубра (зубробизон), тетерева и куропатки (межняк), зайца-русака и зайца-беляка (тумак), соболя и лисицы (кидус), а также тигра и льва (лигр).
В качестве примеров межродовых гибридов растений можно назвать гибрид пшеницы и ржи (тритикале), пшенично-пырейный гибрид, гибрид смородины и крыжовника (йошта), гибрид брюквы и кормовой капусты (куузика), гибриды озимой ржи и житняка, травянистого и древовидного томатов и др.
Гетерозис — явление повышенной жизнеспособности, урожайности, плодовитости гибридов первого поколения, превышающих по этим параметрам обоих родителей.
Уже со второго поколения гетерозисный эффект угасает. По-видимому, это происходит вследствие снижения числа гетерозиготных организмов и повышения доли гомозигот.
Классическими примерами проявления гетерозиса являются мул (гибрид кобылы и осла) и лошак (гибрид коня и ослицы) (рис. 1,2) . Это сильные, выносливые животные, которые могут использоваться в значительно более трудных условиях, чем родительские формы.


Рис. 1. Мул Рис. 2. Лошак
Продолжительность их жизни значительно выше, чем у родительских видов.
Лошак меньше мула ростом и строптив, поэтому менее удобен для использования в хозяйственной деятельности человека.
Гетерозис широко применяют в промышленном птицеводстве, например — бройлерные цыплята, отличающиеся очень быстрым ростом. Цыплёнок-бройлер — финальный гибрид, полученный в результате скрещивания нескольких линий разных пород кур (мясных родительских форм), проверенных на сочетаемость. Первоначально для такого скрещивания использовали породы корниш (в качестве отцовской формы) и белый плимутрок (в качестве материнской формы).
ИСКУССТВЕННЫЙ МУТАГЕНЕЗ
Искусственный мутагенез чаще всего используется как метод селекции растений. Он основан на применении физических и химических мутагенов для получения форм растений с выраженными мутациями. Такие формы в дальнейшем используются для гибридизации или отбора.
В селекции растений широко используется полиплоидия.
Полиплоидия — увеличение числа наборов хромосом в клетках организма, кратное гаплоидному (одинарному) числу хромосом; тип геномной мутации.
Половые клетки большинства организмов гаплоидны (содержат один набор хромосом — n), соматические — диплоидны (2n). Организмы, клетки которых содержат более двух наборов хромосом, называются полиплоидами, три набора — триплоидами (3n), четыре — тетраплоидами (4n) и т. д. Наиболее часто встречаются организмы с числом хромосомных наборов, кратным двум, — тетраплоиды, гексаплоиды (6n) и т. д.
Полиплоиды с нечётным числом наборов хромосом (триплоиды, пентаплоиды и т. д.) обычно не дают потомства (стерильны), т. к. образуемые ими половые клетки содержат неполный набор хромосом — не кратный гаплоидному.
ПОЯВЛЕНИЕ ПОЛИПЛОИДИИ
Благодаря полиплоидии выведены высокоурожайные полиплоидные сорта сахарной свеклы, хлопчатника, гречихи и др. Полиплоидные растения часто более жизнеспособны и плодовиты, чем нормальные диплоиды. О их большей устойчивости к холоду свидетельствует увеличение числа видов-полиплоидов в высоких широтах и в высокогорьях.
Поскольку полиплоидные формы часто обладают ценными хозяйственными признаками, искусственную полиплоидизацию применяют в растениеводстве для получения исходного селекционного материала.
Получение полиплоидов в эксперименте тесно связано с искусственным мутагенезом. С этой целью используют специальные мутагены (например, алкалоид колхицин), нарушающие расхождение хромосом в митозе и мейозе.
Получены урожайные полиплоиды ржи, гречихи, сахарной свёклы и других культурных растений; стерильные триплоиды арбуза, винограда, банана популярны благодаря бессемянным плодам.
Применение отдалённой гибридизации в сочетании с искусственной полиплоидизацией позволило отечественным учёным получить плодовитые полиплоидные гибриды растений (Г. Д. Карпеченко, гибрид-тетраплоид редьки и капусты) и животных (Б. Л. Астауров, гибрид-тетраплоид тутового шелкопряда).
ШЕЛКОПРЯДЫ АСТАУРОВА
биотехнология
Биотехнология — наука, изучающая возможность модификации биологических организмов для обеспечения потребностей человека.
Применение биотехнологии (рис. 3):
- производство лекарств, удобрений, средств биологической защиты растений;
- биологическая очистка сточных вод;
- восстановление ценных металлов из морской воды;
- коррекция и исправление генетических патологий.

Рис. 3. Возможности биотехнологии
Например, включение в геном кишечной палочки гена, ответственного за образование у человека инсулина, позволило наладить промышленное получение этого гормона (рис. 4).

Рис. 4. Биотехнология получения инсулина
В биотехнологии успешно применяются методы генной и клеточной инженерии.
ГЕННАЯ И КЛЕТОЧНАЯ ИНЖЕНЕРИЯ
Генная инженерия — искусственное, целенаправленное изменение генотипа микроорганизмов с целью получения культур с заранее заданными свойствами.
Исследования в области генной инженерии распространяются не только на микроорганизмы, но и на человека. Они особенно актуальны при лечении болезней, связанных с нарушениями в иммунной системе, в системе свертывания крови, в онкологии.
Основной метод генной инженерии: выделение необходимых генов, их клонирование и введение в новую генетическую среду. Например, введение определённых генов с помощью плазмиды в организм бактерии для синтеза ею определённого белка (рис. 5).

Рис. 5. Применение генной инженерии
Основные этапы решения генно-инженерной задачи следующие:
- Получение изолированного гена.
- Введение гена в вектор (плазмиду) для переноса в организм.
- Перенос вектора с геном (рекомбинантной плазмиды) в модифицируемый организм.
- Преобразование клеток организма.
- Отбор генетически модифицированных организмов и устранение тех, которые не были успешно модифицированы.
Клеточная инженерия — это направление в науке и селекционной практике, которое изучает методы гибридизации соматических клеток, принадлежащих разным видам, возможности клонирования тканей или целых организмов из отдельных клеток.
Включает культивирование и клонирование клеток на специально подобранных средах, гибридизацию клеток, пересадку клеточных ядер и другие микрохирургические операции по «разборке» и «сборке» (реконструкции) жизнеспособных клеток из отдельных фрагментов.
На данный момент удалось получить гибриды между клетками животных, далёких по систематическому положению, например мыши и курицы. Соматические гибриды нашли широкое применение как в научных исследованиях, так и в биотехнологии.
Гибридные клетки, полученные от клеток человека и мыши и человека и китайского хомячка, участвовали в расшифровке генома человека.
Гибриды между опухолевыми клетками и лимфоцитами обладают свойствами обеих родительских клеточных линий: они неограниченно делятся и могут вырабатывать определённые антитела. Такие антитела применяют в лечебных и диагностических целях в медицине.
В эмбриологии для изучения процессов дифференцировки клеток и тканей в ходе онтогенеза используют организмы—химеры, состоящие из клеток с разными генотипами.Их создают путём соединения клеток разных зародышей на ранних этапах их развития.
Клонирование животных — ещё один метод клеточной инженерии: ядро соматической клетки пересаживают в лишённую ядра яйцеклетку с последующим выращиванием зародыша во взрослый организм.
Преимущество клеточной инженерии в том, что она позволяет экспериментировать с клетками, а не с целыми организмами.
Методы клеточной инженерии часто применяют в сочетании с генной инженерией.
работы Н. И. Вавилова
Николай Иванович Вавилов — российский генетик, растениевод, географ.
- Н. И. Вавилов организовал 180 экспедиций (20−30 гг. ХХ века) по самым труднодоступным и зачастую опасным районам земного шара с целью изучения многообразия и географического распространения культурных растений.
- Им была собрана уникальная, самая крупная в мире коллекция культурных растений (к 1940 г. коллекция включала 300 000 образцов), которые ежегодно размножаются в коллекциях Всероссийского института растениеводства имени Н. И. Вавилова (ВИР) и широко используются селекционерами как исходный материал для создания новых сортов зерновых, плодовых, овощных, технических, лекарственных и других культур.
- Создал учение об иммунитете растений.
УЧЕНИЕ ОБ ИММУНИТЕТЕ РАСТЕНИЙ
- Закон гомологических рядов наследственной изменчивости: у генетически близких видов и родов существуют гены, которые дают сходные признаки. Таким образом, можно предсказать наличие признаков у других видов известного рода.
- Установил, что наибольшее разнообразие форм вида сосредоточено в тех районах, где этот вид возник. Н. И. Вавилов выделил 8 центров происхождения культурных растений.
ЦЕНТРЫ ПРОИСХОЖДЕНИЯ КУЛЬТУРНЫХ РАСТЕНИЙ
Центры происхождения культурных растений — географические области, являющиеся родиной дикорастущих предков культурных растений.
Центры происхождения важнейших культурных растений связаны с древними очагами цивилизации и местом первичного возделывания и селекции растений. Подобные очаги одомашнивания (центры доместикации) выявлены и у домашних животных.
Было выделено восемь центров происхождения культурных растений (рис. 6):
1. Средиземноморский (спаржа, маслины, капуста, лук, клевер, мак, свекла, морковь).
2. Переднеазитский (инжир, миндаль, виноград, гранат, люцерна, рожь, дыня, роза).
3. Среднеазиатский (нут, абрикос, горох, груша, чечевица, лен, чеснок, мягкая пшеница).
4. Индо-Малайский (цитрусовые, хлебное дерево, огурец, манго, черный перец, кокосовая пальма, банан, баклажан).
5. Китайский (просо, редька, вишня, яблоко, гречиха, слива, соя, хурма).
6. Центральноамериканский (тыква, фасоль, какао, авокадо, махорка, кукуруза, батат, хлопчатник).
7. Южноамериканский (табак, ананас, томат, картофель).
8. Абиссинский центр (банан, кофе, сорго, твердая пшеница).
В поздних работах Н. И. Вавилова Переднеазиатский и Среднеазиатский центры объединяются в Юго-западноазиатский центр.

Рис. 6. Центры происхождения культурных растений
В настоящее время выделяют 12 первичных центров происхождения культурных растений.
Биология ЕГЭ Задание 4 (2022 год) проверяет знания о закономерностях наследственности и их цитологических основах. Чтобы выполнить такое задание, необходимо иметь чёткое представление о доминантных и рецессивных признаках, аллельных генах, анализирующем скрещивании, гетеро- и гомогаметности, знать основные законы генетики, уметь составлять схему скрещивания с указанием генотипов родителей, гамет, генотипов и фенотипов потомства.
Выбрать другое задание
Вариант ЕГЭ с пояснениями
Кодификатор ЕГЭ
Линия 4 ЕГЭ по Биологии. Моно- и дигибридное, анализирующее скрещивание. Решение биологической задачи. Коды проверяемых элементов содержания (КЭС): 3.5. Уровень сложности: Б. Максимальный балл: 1. Примерное время выполнения: 5 мин. Средний % выполнения: 64,7.
Задание представляет собой текстовую задачу по генетике. В ответе необходимо записать буквы, последовательность цифр в порядке убывания или конкретное число.
Алгоритм выполнения задания № 04 на ЕГЭ по биологии:
- Внимательно прочитайте задачу.
- Проанализируйте, о взаимодействии каких генов идёт речь.
- Составьте схему скрещивания.
- Если необходимо, выполните вычисления на черновике.
- Запишите ответ в виде последовательности цифр в порядке убывания (если надо определить соотношение получившихся фенотипов), букв (если необходимо определить гаметы), чисел (если нужно вычислить вероятность, процентное соотношение, количество) в поле ответа КИМ и бланк ответов № 1.
Примечание: Необходимо учитывать регистр начертания букв: ЗАГЛАВНЫЕ буквы — доминантный ген / признак (А, В), строчные — рецессивный ген / признак (а, b).
Задание 4 (пример выполнения с решением)
Линия 04. Пример № 1.
Определите соотношение фенотипов у потомства, полученного от скрещивания особей с генотипами АаВb и АаВВ. Ответ запишите в виде последовательности цифр, показывающих соотношение получившихся фенотипов, в порядке их убывания.
Правильный ответ: 6 : 2
Пояснение (решение).

Теория, которую необходимо повторить
В период подготовки к экзамену ПОВТОРЯЕМ теорию по конспектам:
| КЭС 3.5. Закономерности наследственности, их цитологические основы. Закономерности наследования, установленные Г. Менделем, их цитологические основы (моно- и дигибридное скрещивание). Законы Т. Моргана: сцепленное наследование признаков, нарушение сцепления генов. Генетика пола. Наследование признаков, сцепленных с полом. Взаимодействие генов. Генотип как целостная система. Генетика человека. Методы изучения генетики человека. Решение генетических задач. Составление схем скрещивания | Конспект 1, Конспект 2, Конспект 3, Конспект 4, Конспект 5, Конспект 6, Конспект 7 |
Нажмите на спойлер ниже, чтобы посмотреть основный теоретический материал к данной линии: основные генетические понятия, первый, второй и третий законы Менделя, анализирующее скрещивание.
Открыть краткий справочный материал к заданию № 6
Тренировочные задания
Выполните самостоятельно примеры задания № 4 и сверьте свой ответ с правильным (спрятан в спойлере).
Пример № 2.
Способность лучше владеть правой рукой у человека доминирует над леворукостью. Женщина-правша, отец которой был левшой, вышла замуж за мужчину-левшу. Какова вероятность того, что в этой семье родится леворукий ребёнок? В ответе запишите только цифры.
Посмотреть правильный ОТВЕТ и пояснение
Пример № 3.
У человека карий цвет глаз доминирует над голубым, а праворукость — над леворукостью. Гены не сцеплены. Определите соотношение фенотипов, если родители — кареглазые правши, гетерозиготные по двум парам признаков. Ответ запишите в виде последовательности цифр, показывающих соотношение получившихся фенотипов, в порядке их убывания.
Нажмите на спойлер, чтобы увидеть ОТВЕТ
Пример № 4.
Сколько типов гамет образует зигота АаВbСс, если гены не сцеплены? В ответ запишите только соответствующее число.
Нажмите на спойлер, чтобы увидеть ОТВЕТ
Пример № 5.
У отца отрицательный резус-фактор крови, а у матери — положительный (гетерозигота). Какова вероятность (в %) рождения детей с положительным резус-фактором? В ответе запишите только соответствующее число.
Нажмите на спойлер, чтобы увидеть ОТВЕТ
Пример № 6.
Сколько разных фенотипов образуется при скрещивании двух дигетерозигот, если гены находятся в разных хромосомах? Ответ запишите в виде цифры.
Нажмите на спойлер, чтобы увидеть ОТВЕТ
Вы смотрели: Биология ЕГЭ Задание 4. Что нужно знать и уметь, план выполнения, примеры с ответами и пояснениями (комментариями) специалистов, анализ типичных ошибок.
Выбрать другое задание
Вариант ЕГЭ с пояснениями
Кодификатор ЕГЭ
Биология ЕГЭ Задание 4