Пройти тестирование по этим заданиям
Вернуться к каталогу заданий
Версия для печати и копирования в MS Word
1
В настоящее время на пути биологического регресса находятся
2
В настоящее время на пути биологического прогресса находятся
3
Появление у древних млекопитающих четырёхкамерного сердца, теплокровности, развитой коры головного мозга — пример
3) биологического прогресса
4) биологического регресса
4
Появление большого разнообразия видов насекомых на Земле — следствие развития их по пути
3) биологического регресса
4) идиоадаптации
5
Какой из перечисленных ароморфных признаков позволил млекопитающим освоить разнообразные среды обитания
4) развитие коры больших полушарий
Пройти тестирование по этим заданиям
Ароморфозы, идиоадаптации, общие дегенерации – всё это способы достижения биологического прогресса. Разберем их подробнее.
Ароморфоз — масштабное, значительное эволюционное изменение, приводящее к общему подъему организации, усиливающее интенсивность жизнедеятельности. Нужно понимать, что ароморфоз не является каким-то мелким приспособлением к условиям среды при их резком изменении, это действительно глобальная перемена.
Результаты ароморфозов
1. Повышение организации – крупные, значительные изменения.
2. Развитие приспособлений широкого значения. Следствие – рост выживаемости особей, снижение смертности в популяции.
3. Расширение среды обитания.
4. Образование новых популяций.
5. Ускорение процесса формирования новых видов.
Примеры ароморфозов у животных в целом
1. Живорождение.
2. Поддержание стабильной температуры тела.
3. Формирование замкнутой кровеносной системы.
Примеры ароморфозов у беспозвоночных
1. Половая дифференциация.
2. Билатеральная организация.
3. Появление трахейной системы дыхания.
4. Цефализация ЦНС.
Примеры ароморфозов у птиц
1. Появление крыла в качестве органа полета.
2. Возникновение совершенного четырехкамерного сердца.
3. Развитие отделов мозга, координирующих движения в полете.
Примеры ароморфозов у растений
1. Выход на сушу.
2. Возникновение пыльцевой трубки у голосеменных.
3. Появление цветка.
4. Появление сосудистой системы.
5. Способность к поддержанию и регулированию газообмена в листьях.
Казалось бы, мы перечислили очень разные процессы. Но их, все эти ароморфозы, объединяет то, что в ходе дальнейшей эволюции они не исчезают, а закрепляются и сохраняются, что ведет к появлению больших систематических групп: типов, отделов, классов, некоторых отрядов (у млекопитающих).
Идиоадаптация — способ достижения биологического прогресса, при которых организмы развивают некие частные приспособления, позволяющие им освоить дифференцированные (узкие) условия окружающей среды. Таким образом организмы могут прогрессировать без перехода на более высокий уровень морфофизиологической организации.
Приведем пример. Первые млекопитающие занимали маленькое место на планете – они робко шуршали в подлеске, опасаясь рептилий. Но сейчас, спустя миллионы лет, млекопитающие распространены по всей Земле, они научились жить не только на суше (в холодных снегах и жарких пустынях), но и в почве, и в воде (как пресной, так и соленой). Достигли этого млекопитающие благодаря череде идиоадаптаций – то есть частных изменений в строении и функциях разных органов без изменения уровня общей организации. Идиоадаптации ощутимо снижают уровень конкуренции между видами – им больше не нужно отчаянно бороться за пропитание и территорию (можно отрастить более плотный мех – и ты уже осваиваешь холодное предгорье, оставив конкурентов на равнине; или заполучить длинные, крепкие корни и доставать воду из глубоких слоев почвы).
Здесь есть тонкость. Неверно представлять, что частные приспособления при идиоадаптации – это всегда усложнение. Возможно и упрощение организма, «отбрасывание» органов и умений за ненадобностью. Этот путь называется общей дегенерацией. Если организмы попадают в более простую среду, чем та, в которой они жили до сих пор, у них упрощается строение.
Итак, общая дегенерация приводит к морфофизиологическому регрессу, который связан с сидячим, неподвижным, образом жизни, однородной, простой и скучной, средой обитания, с паразитическим существованием.
1. Например, рыбы, заплывшие в пещеру по ручью и оставшиеся жить в подземном озере, в кромешном мраке, утрачивают глаза и пигментацию чешуи.
2. Кроты, живущие под землей, имеют очень слабое зрение, их маленькие глазки плотно закрыты веками – вероятнее всего, кроты постепенно тоже утрачивают органы зрения.
3. Паразитическая лиана, питающаяся соками своего хозяина, теряет листву, у нее замедляется фотосинтез.
4. Повилика утрачивает хлорофилл.
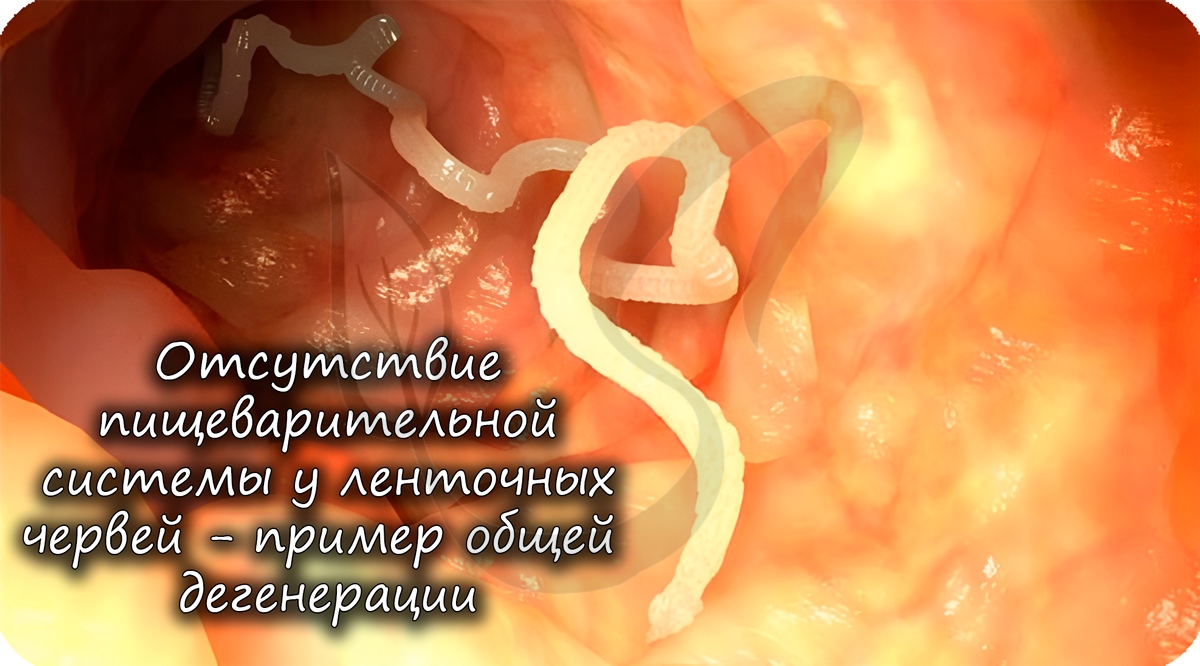
5. У ленточных червей, паразитирующих в организмах млекопитающих, редуцировался не только кишечник, но и нервная система.
6. У асцидий во взрослом состоянии редуцируется хорда.
Казалось бы, слепые рыбы, лианы без листьев и черви без нервов должны умереть. Но этого не происходит, напротив, организмы процветают – у них время биологического прогресса.
Пути эволюции
В своих работах советский ученый Северцов А.Н. выделил понятия биологического прогресса и регресса.
Биологический прогресс подразумевает победу вида в борьбе за существование. Биологический прогресс характеризуется следующими признаками:
- Численность вида увеличивается
- Ареал расширяется
- Смертность особей уменьшается
- Рождаемость увеличивается
- Происходит процветание вида
Основными направлениями биологического прогресса являются:
- Ароморфоз (греч. airomorphosis — поднимаю форму)
- Идиоадаптация (греч. ídios — свой, своеобразный, особый)
- Общая дегенерация (лат. degenero — вырождаться, перерождаться)
Ароморфоз представляет собой прогрессивное эволюционное преобразование, повышающее уровень организации организмов.
В результате ароморфоза становится возможным освоение новых, ранее недоступных для жизни, территорий. К примеру,
теплокровность птиц позволила им заселить места с холодным климатом.

Идиоадаптация подразумевает незначительные, частные изменения в строении и функциях организма, которые помогают
приспособиться к условиям среды обитания. Идиоадаптации существенно не повышают уровень организации.
Общей дегенерацией называют упрощение организации, которое заключается в утрате отдельных органов и систем органов.
У многих этот пункт вызывает внутреннее противоречие: как общая дегенерация может относиться к биологическому прогрессу?
На самом деле, если орган или система органов не нужна организму в его условиях обитания — то зачем она? Эта система
может исчезнуть и освободить место для других, более полезных в данных условиях, органов.
У многих паразитов отсутствуют различные органы, к примеру, у ленточных червей нет пищеварительной системы. А зачем она
им, когда пища в кишке, где они обитают, уже переварена и расщеплена организмом хозяина?
Биологический регресс характеризуется признаками, противоположными биологическому прогрессу:
- Численность вида уменьшается
- Ареал сужается
- Смертность особей возрастает
- Рождаемость уменьшается
- Происходит вымирание вида
Главная причина биологического регресса в том, что скорость эволюции вида отстает от скорости изменения внешней среды, эволюции других видов: это несоответствие снижает приспособленность организмов. Часто деятельность человека молниеносно
меняет окружающую среду: далеко не все виды могут приспособиться к этому, происходит вымирание.

Сравнительно-анатомические доказательства эволюции
Изучение строения органов и их эволюционных изменений у различных групп организмов является основой выявления сравнительно-анатомических доказательств эволюции. Яркими примерами анатомических доказательств эволюции являются гомологичные
и аналогичные органы.
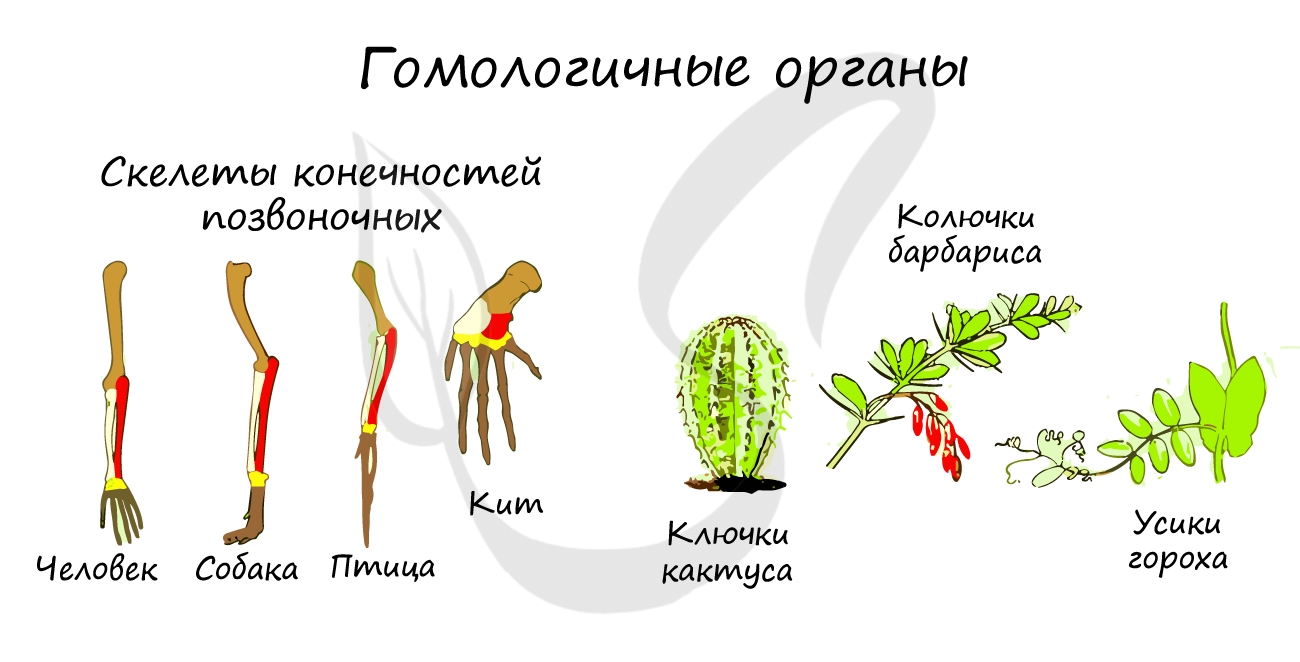
- Гомологичные органы (гомология, от греч. homo(s) — равный, одинаковый)
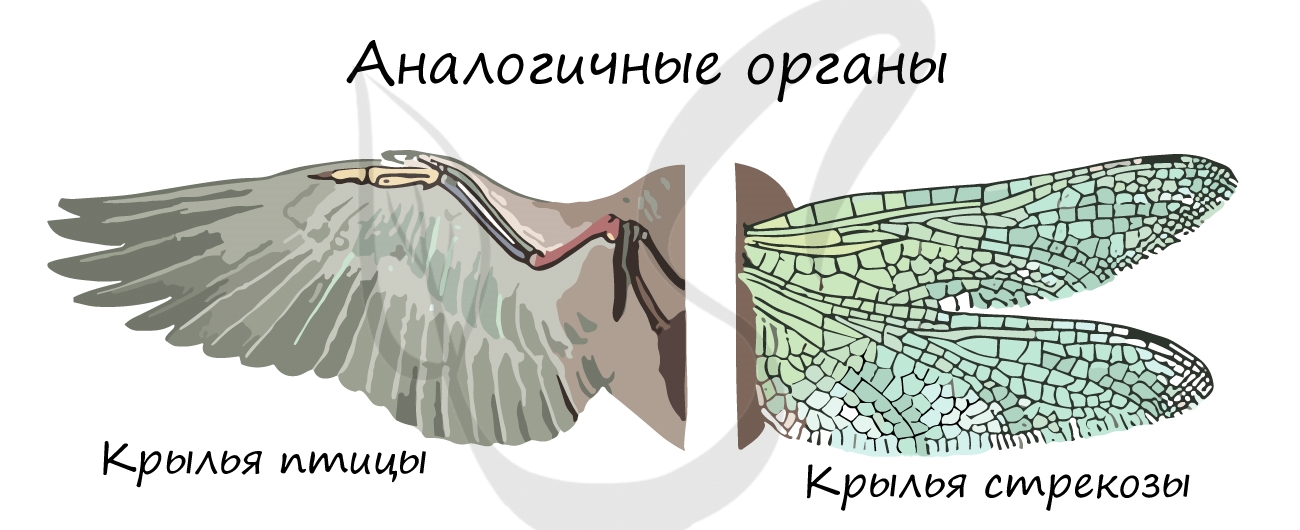
- Аналогичные органы (греч. análogos — соответственный)
Такие органы развиваются из одних и тех же зародышевых листков, имеют общий план строения, но выполняют разные функции.
Это связано с тем,
что животные освоили разные среды обитания, из-за чего происходит дивергенция (лат. divergo — отклоняюсь) —
расхождение признаков у первоначально близких животных в ходе эволюции.
Гомологичны между собой скелеты конечностей различных классов позвоночных: рука — ласт — крыло птицы, колючки кактуса
— усики гороха — листья растений.
Аналогичные органы развиваются из разных зародышевых листков, имеют различное строение, но выполняют схожие
функции. Такое сходство возникает в результате приспособления к одним и тем же условиям среды, из-за чего
происходит конвергенция (лат. convergo — сближаю) — схождение признаков у неблизкородственных видов в ходе эволюции.
Аналогичными органами являются крыло птицы — крыло бабочки, глаз человека — глаз кальмара, усики винограда — усики
гороха, жабры рака — жабры рыбы.
В строении нынешних животных можно найти признаки древних предковых форм, которые также свидетельствуют об эволюции. Сейчас
мы обсудим рудименты и атавизмы.
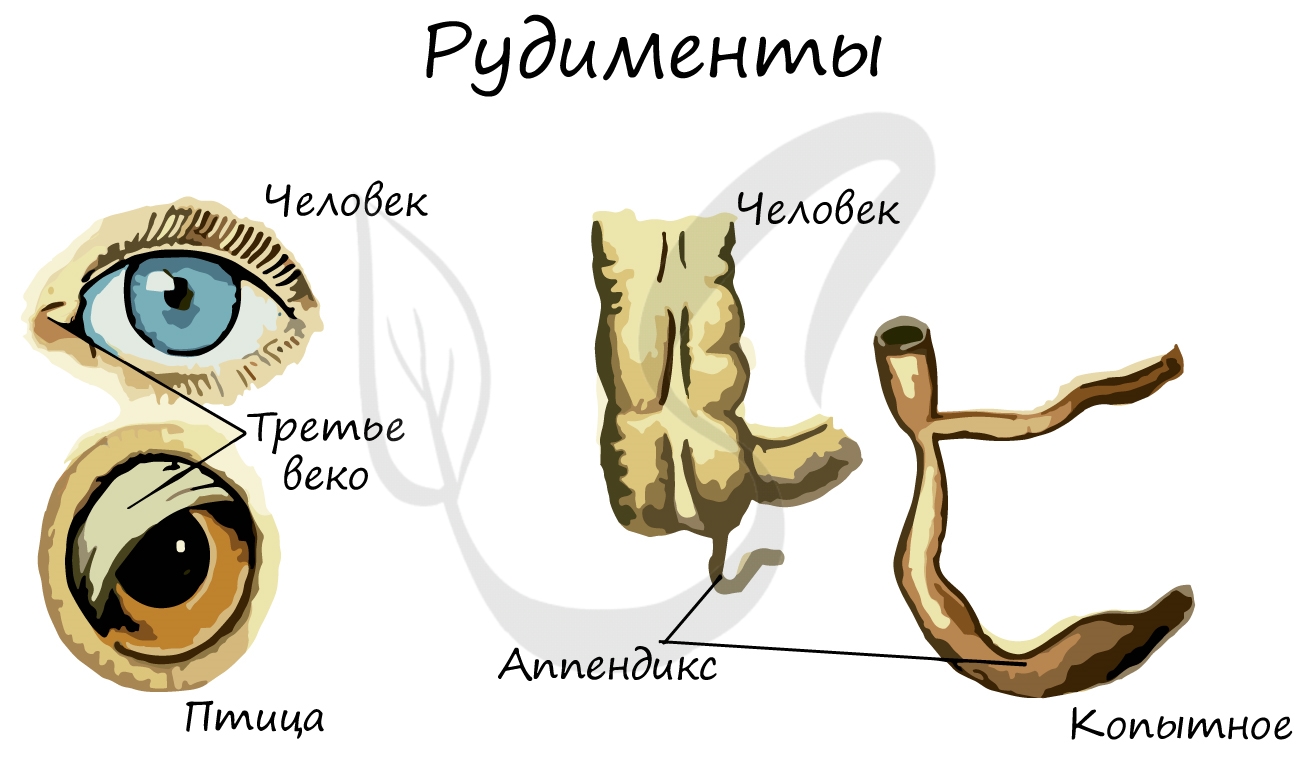
Рудименты (лат. rudimentum — зачаток) — органы, которые в ходе эволюции утратили свое функциональное значение. Они
сохраняются в течение всей жизни и в норме обнаруживаются у человека и животных.
У человека к рудиментарным органам относятся: зубы мудрости, копчик, ушные мышцы, аппендикс (червеобразный отросток),
третье веко.
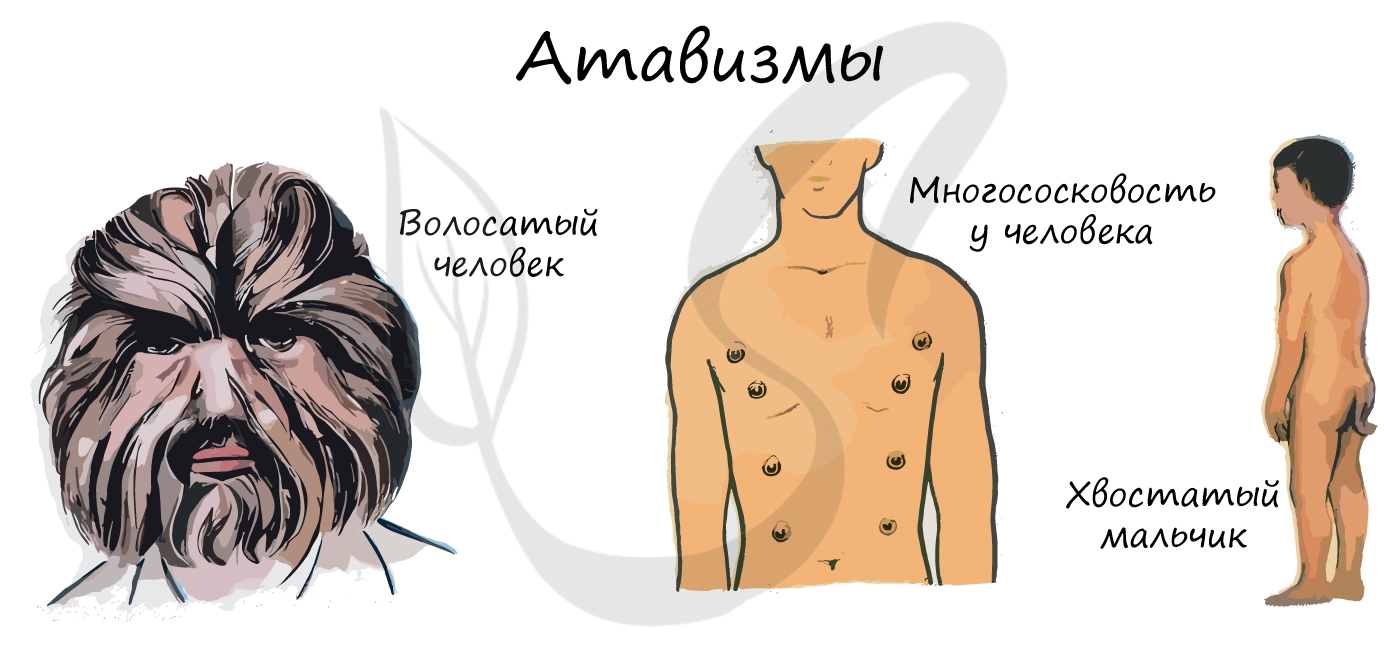
Атавизмы (лат. atavus — отдалённый предок) — случаи проявления у отдельных особей признаков дальних предков. Атавизмы
сугубо индивидуальны и не являются нормой. Они также являются доказательством эволюции.
У человека атавизмами могут являться хвост, волосатое тело, добавочные молочные железы, незаращение межпредсердной перегородки.
Переходные формы
Переходные формы свидетельствуют о филогенетической преемственности, соединяя в своем строении черты высших и низших классов. Они —
наглядное, живое доказательство эволюции.
Такими формами являются, к примеру, утконос и ехидна из класса млекопитающих. При многих признаках млекопитающих, они откладывают яйца, тем самым подтверждают родство
млекопитающих с пресмыкающимися.

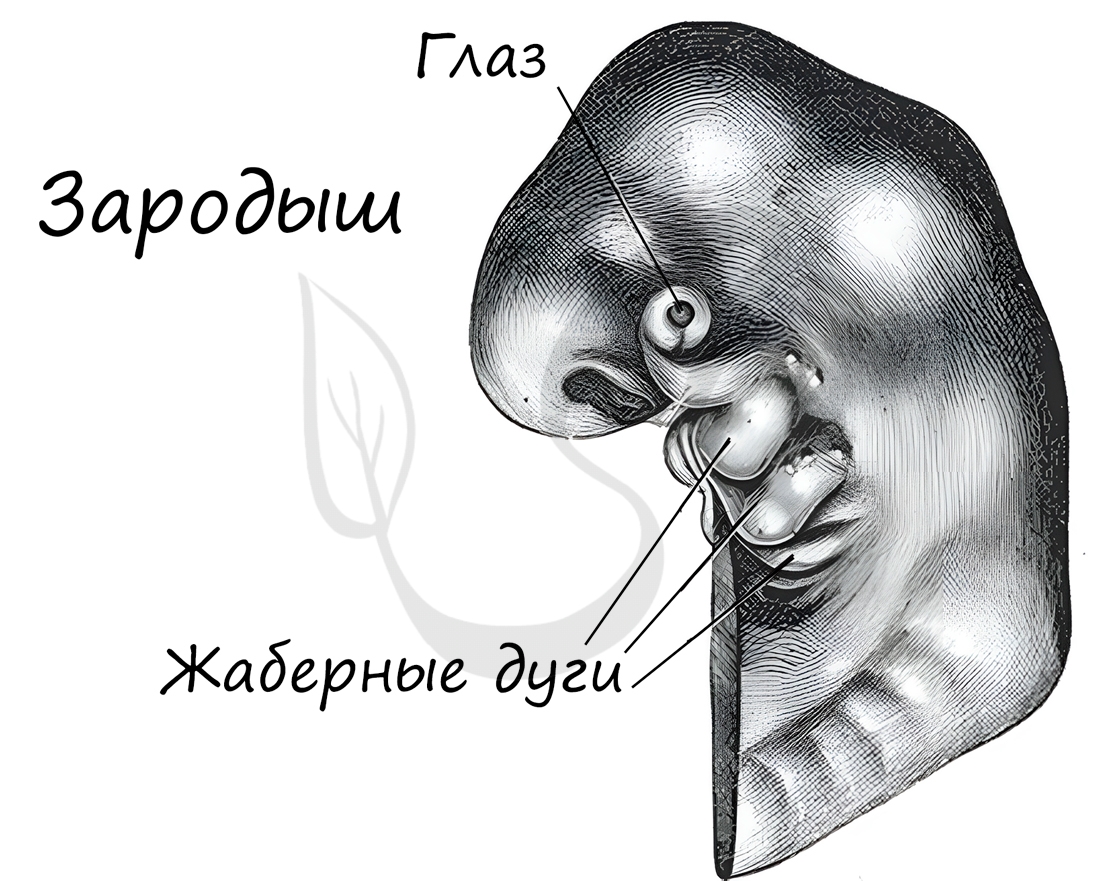
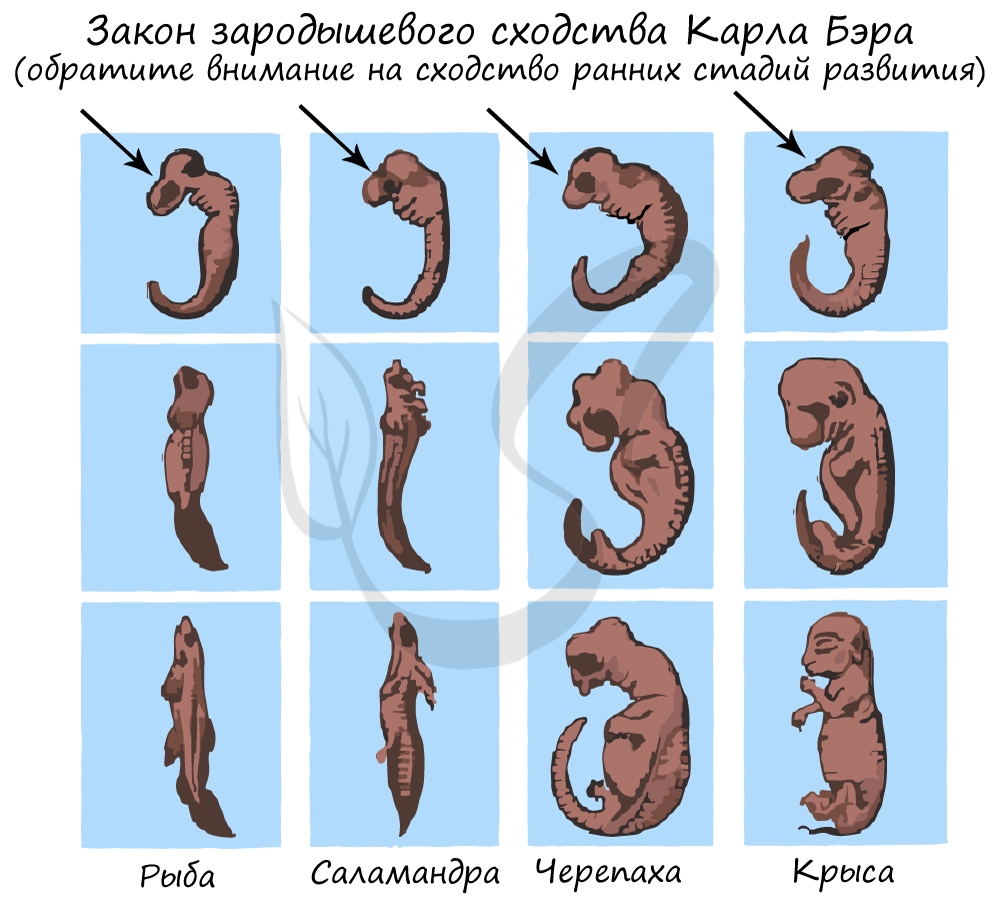
Эмбриологические доказательства
Эмбриология (греч. embryon — зародыш) — раздел биологии, изучающий строение эмбрионов. Только вдумайтесь: на этапе эмбриона,
через который мы с вами успешно прошли, у нас можно было найти закладку жаберных дуг, которые существуют непродолжительное время,
после чего исчезают.
А у рыб, например, жаберные дуги не исчезают — из них развиваются жабры.
Немецкие ученые Ф. Мюллер и Э. Геккель во второй половине XIX века сформулировали биогенетический закон, гласящий, что
онтогенез (индивидуальное развитие) каждой особи есть краткое и быстрое повторение филогенеза (исторического развития вида).
Биогенетический закон Мюллера-Геккеля объясняет повторение этапов (на стадии зародыша), которые были свойственны нашим далеким
предкам. Таким образом, мы проходим их этапы, но, не останавливаясь на них, двигаемся дальше к более совершенным этапам.
У головастиков лягушек развивается плавник, есть жабры — это наглядное повторение признаков, которые характерны для их предков — рыб.
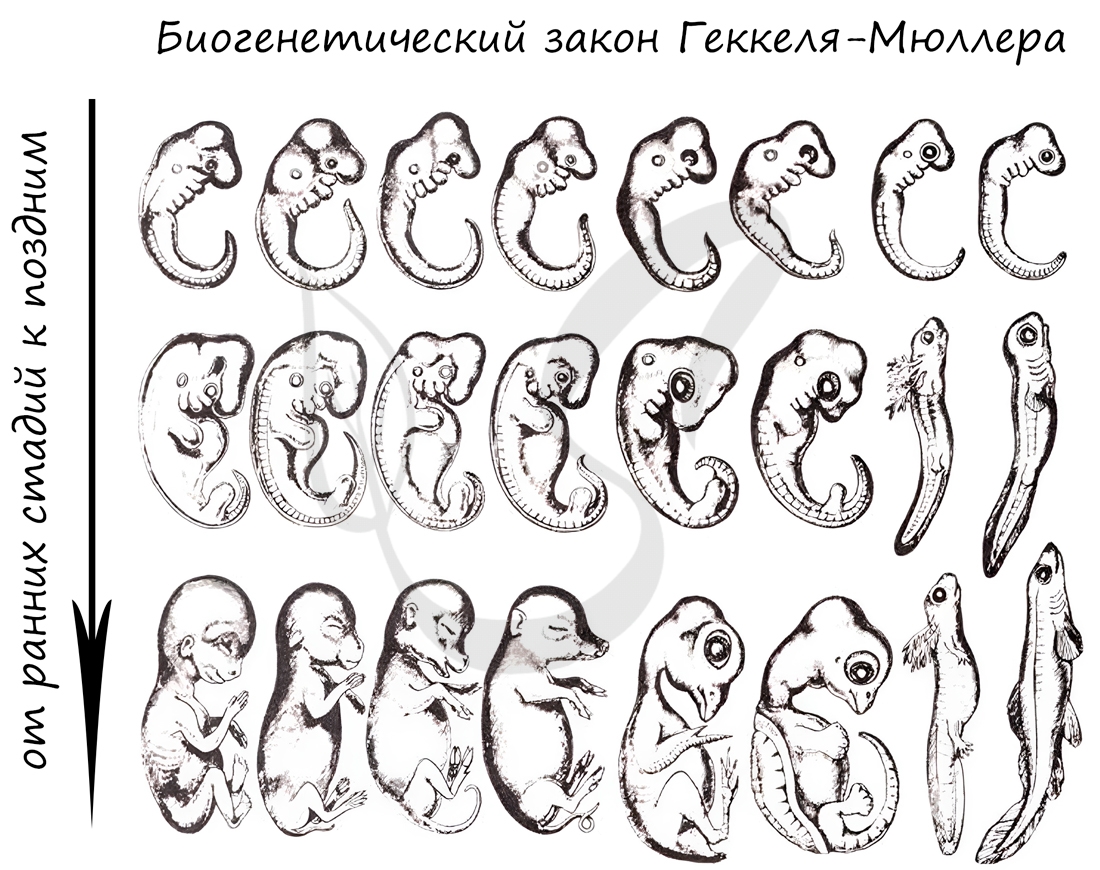
Карл Бэр сформулировал закон зародышевого сходства, который гласит, что на ранних стадиях развития зародыши позвоночных животных
настолько похожи друг на друга, что практически неразличимы между собой. Это также указывает и подтверждает единство происхождения
животного мира.
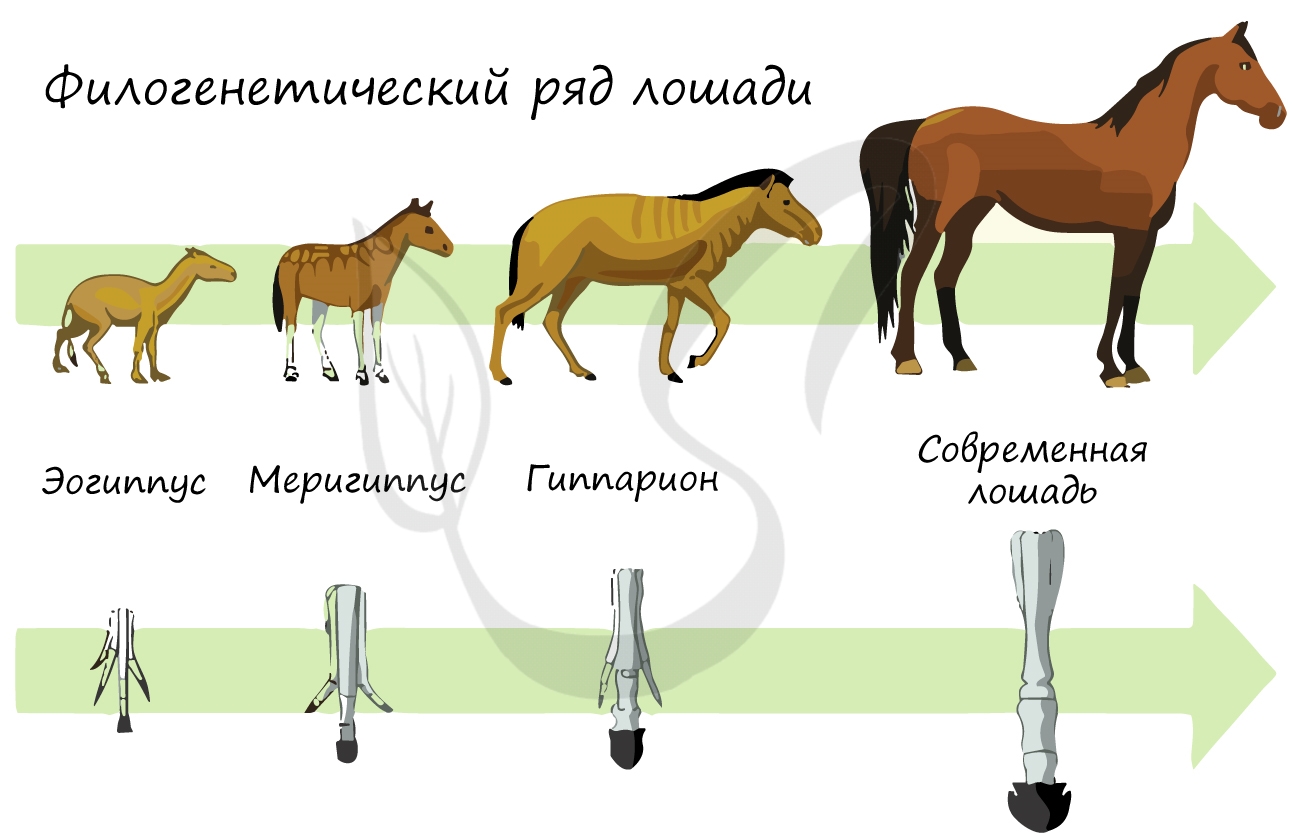
Палеонтологические доказательства эволюции
Палеонтология (греч. palaios – древний) изучает ископаемые останки вымерших животных, их сходства и различия с ныне живущими
видами. Сопоставляя друг с другом ископаемые останки разных геологических эпох, можно увидеть как происходила эволюция различных
видов животных и растений.
В результате таких исследований иногда удается открыть переходные формы, а иногда — целые филогенетические ряды, то есть совокупность
последовательно сменяющих друг друга форм одного вида. Так, к примеру, был открыт филогенетический ряд лошади.
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Популяция — самая мелкая из групп особей, способная к эволюционному развитию, поэтому её называют элементарной единицей эволюции.
Отдельно взятый организм не может являться единицей эволюции — эволюция происходит только в группе особей.
Естественный отбор идет по фенотипам (признакам), по-этому для эволюции нужно разнообразие особей в популяции. Отбирая наиболее «выгодные» фенотипы, естественный отбор оставляет «выгодные» генотипы (комбинации генов). В результате выживают и оставляют потомство особи с наиболее выгодными в данных условиях генотипами.
Совокупность генотипов всех особей популяции — генофонд — основа микроэволюционных процессов в природе.
Вид как целостная система не может быть принят за единицу эволюции, т.к. обычно виды распадаются на составные их части — популяции. Вот почему роль элементарной эволюционной единицы принадлежит популяции.
Общая схема микроэволюции
Микроэволюция — эволюционные процессы, происходящие на уровне популяции (т. е. внутри вида), ведущие к накоплению наследственных особенностей и приводящие к образованию нового вида.

Популяция является элементарной единицей эволюции.
Генофонд популяции — это совокупность генов организмов данной популяции.
Элементарные эволюционные факторы — факторы, способствующие изменению генофонда популяций: мутации, миграции, поток генов, популяционные волны, дрейф генов, изоляция.
МИГРАЦИИ И ПОТОК ГЕНОВ
Поток генов — перенос генов между популяциями.
Большую роль в осуществлении потока генов играют миграции, кочевки, перелеты, перенос пыльцы и семян ветром, насекомыми. В зависимости от вида организмов в каждом поколении, по мнению Э. Майра, имеется от 30 до 50 % пришельцев . Именно благодаря потоку генов на обширных территориях наблюдается фенотипическая однородность особей.
ЗАКОН МАЙРА
Благодаря свободному скрещиванию при миграции происходит обмен генами между особями популяции одного вида (поток генов). При этом гены мигрирующих особей включаются при скрещивании в генофонд популяций. В результате генофонд популяций обновляется.
Например, клоп-черепашка разлетается по направлению ветра. Клопы не обязательно возвращаются в места рождения. Дальность полета на зимовку зависит от упитанности. В результате на зимовках оказываются клопы из разных мест. Часть клопов вообще не улетает далеко, а остается зимовать в ближайших лесопосадках.
Группы крови человека системы АВО: частота гена А меняется с Востока на Запад — от низкой к высокой, частота гена В, наоборот, от высокой к низкой. Такой градиент концентраций этих генов объясняют крупными миграциями людей с азиатского Востока в Европу в период с 500 до 1500 гг. н. э.
ДРЕЙФ ГЕНОВ
Дрейф генов — случайное изменение концентрации аллелей в небольшой, полностью изолированной популяции.
Дрейф генов непредсказуем. Небольшую популяцию он может привести к гибели, а может сделать ее еще более приспособленной к данной среде и усилить ее дивергенцию от родительской популяции.
Он происходит вследствие увеличения количества гомозигот при близкородственном скрещивании.
В 1419 г. на корабле случайно оказалась беременная крольчиха, которая родила во время путешествия. Все детеныши были выпущены на остров Порту-Санту. Популяция кроликов на острове сильно увеличилась. Кролики сильно уменьшились в размерах. По окраске кролик с Порту-Санту значительно отличается от обыкновенного. Они необычайно дики и проворны. По своим привычкам они более ночные животные. С другими породами не скрещиваются (образование нового вида).
МУТАЦИИ
Мутация — случайное скачкообразное изменение генотипа.
Генные мутации, затрагивающие доминантные гены, а также хромосомные и геномные мутации чаще снижают приспособленность особи и не так важны для эволюции. Хотя известно, что в природе полиплоидные формы растений имеют преимущество перед диплоидными.
Возможны следующие исходы проявления мутаций:
- летальные (не совместимые с жизнью) мутации исчезнут из популяции вместе с их носителями;
- мутации, вызывающие стерильность особей, не могут иметь значения, так как их носители бесплодны;
- мутации, не оказывающие отрицательного воздействия на особь, включаются в генофонд популяций.
Следовательно, фенотипически однородная природная популяция является гетерогенной, что обусловливает ее возможность эволюционировать.
Популяция, как губка, накапливает мутации, при этом ее приспособленность не нарушается. Следовательно, рецессивные мутации представляют собой «скрытый резерв наследственной изменчивости», что важно для эволюционного процесса.
Популяции на протяжении многих поколений стабильны и относительно однородны. Это объясняется действием стабилизирующего отбора. А поскольку отбор идет по фенотипу, то возможность сохранения мутанта будет определяться степенью нарушения приспособленности этой особи. Сильно уклонившиеся формы устраняются отбором. Таким образом поддерживается внешняя стабильность популяции.
Материал для эволюционного процесса дает и комбинативная изменчивость. Создавая новые сочетания генов в генотипе, она увеличивает разнообразие особей в популяции и предоставляет естественному отбору поле деятельности.
ПОПУЛЯЦИОННЫЕ ВОЛНЫ (ВОЛНЫ ЖИЗНИ)
Популяционные волны — колебания численности особей в популяции. Их причинами могут быть различные изменения окружающей среды: засуха, наводнения, снежные зимы, болезни, наличие паразитов, врагов, нехватка кормовых ресурсов и др. В урожайные годы численность особей в какой-либо популяции может повыситься, вслед за чем произойдет ее спад.
Например, увеличение количества зайцев через некоторое время приводит к возрастанию числа волков и рысей из-за достаточного количества пищи (зайцев).
Волны жизни приводят к изменению концентраций аллелей в генофонде популяций. При снижении особей в популяции из ее генофонда могут выпасть редкие аллели, и наоборот, при возрастании количества особей такие аллели могут распространяться. Популяционные волны, таким образом, случайны и служат поставщиком эволюционного материала.
В малочисленных популяциях (менее 500 особей), просуществовавших на протяжении многих поколений в изоляции от других популяций своего вида, влияние случайных факторов может выйти на первый план по отношению к действию отбора. Случайное изменение концентраций аллелей в популяции называется дрейфом генов .
ИЗОЛЯЦИЯ
Изоляция — возникновение любых барьеров, ограничивающих свободное скрещивание. Различают пространственную и биологическую изоляцию.
Пространственная изоляция может привести к глубоким внутренним различиям, к генетической несовместимости и, следовательно, к возникновению новых видов.
Биологическая изоляция может произойти на одной территории между группами особей с измененными поведением, морфологическими, функциональными и другими признаками, препятствующими скрещиванию.
Изоляция как эволюционный фактор не создает новых генотипов или внутривидовых форм. Значение ее в эволюции состоит в том, что она закрепляет и усиливает начальные стадии генотипической дифференцировки. Действие изоляции, как и других факторов, ненаправленно.
Таким образом, мутации, миграции, популяционные волны, дрейф генов, изоляция —ненаправленные факторы эволюции. В природе они действуют совместно, однако роль каждого может усиливаться в конкретной обстановке. Все эти факторы обеспечивают генетическую неоднородность популяций.
Макроэволюция — процесс формирования надвидовых таксонов (семейств, отделов, типов, классов).
К маакроэволюции можно отнести и возникновение и развитие жизни на Земле.
Процесс эволюции не обязательно связан с усложнением организации. Именно поэтому в современной живой природе одновременно с высокоорганизованными формами существуют и низкоорганизованные. Ж. Б. Ламарк объяснял существование примитивных форм постоянным самозарождением простых организмов из неорганической материи. Ч. Дарвин же считал, что существование высших и низших форм не представляет затруднений для объяснения, «так как естественный отбор, или выживание наиболее приспособленных, не предполагает обязательного прогрессивного развития — он только дает преимущество тем изменениям, которые благоприятны для обладающего ими существа в сложных условиях жизни… А если от этого нет никакой пользы, то естественный отбор или не будет вовсе совершенствовать эти формы, или усовершенствует их в очень слабой степени, так что они сохранятся на бесконечные времена на их современной низкой ступени организации».
ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
К этой проблеме в начале 20-х годов обратился А. Н. Северцов. Учение о прогрессе в эволюции было в дальнейшем развито его учеником И. И. Шмальгаузеном. К основным направлениям эволюции относятся:
- Биологический прогресс
- Биологический регресс
биологический прогресс
Биологический прогресс — возрастание приспособленности организмов к окружающей среде (по А. Н. Северцову).
Критерии биологического прогресса:
- увеличение численности;
- повышение видового разнообразия (прогрессивная дифференциация);
- расширение ареала.
МЕХАНИЗМ БИОЛОГИЧЕСКОГО ПРОГРЕССА
- возникновение новых приспособлений снижает гибель особей
- средний уровень численности вида возрастает
- увеличивается плотность населения
- обостряется внутривидовая конкуренция + возрастает приспособленность
- расширяется ареал
- вид заселяет новые территории и вынужден приспосабливаться к новым условиям
- отдельные популяции приобретают разные признаки (дивергенция признаков)
- образование дочерних таксонов
ПУТИ БИОЛОГИЧЕСКОГО ПРОГРЕССА
| путь биологического прогресса | изменение | пример |
|---|---|---|
| Арогенез — путь развития группы организмов, характеризующийся повышением уровня морфофизиологической организации, освоением новой среды обитания. | ароморфоз —морфофизиологический прогресс |
Возникновение и расцвет класса птиц. Ароморфозы: крыло, четырехкамерное сердце, теплокровность. |
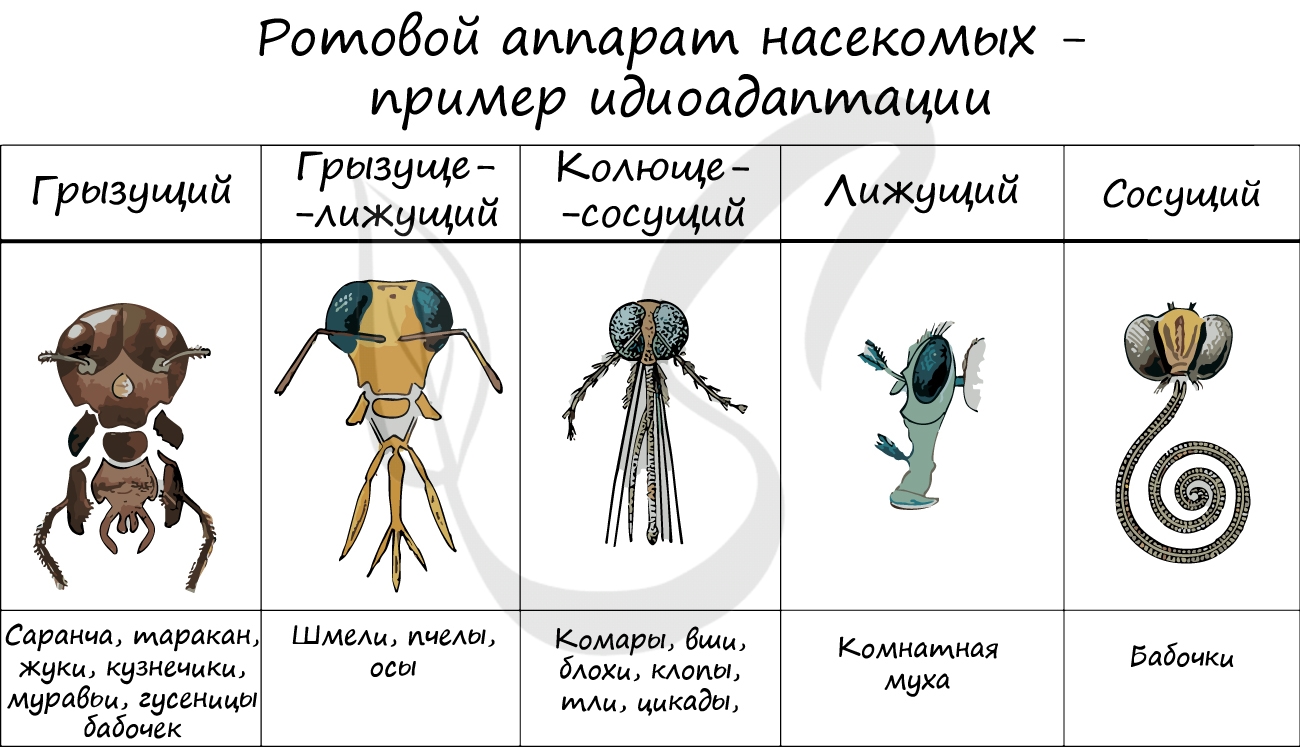
| Аллогенез — путь развития группы организмов, связанный с развитием частных приспособлений к окружающей среде, а уровень организации остается прежним. | алломорфоз, или идиоадаптация — приспособления к окружающей среде | Разная форма ротового аппарата насекомых; покровительственная и защитная окраска; мимикрия. |
| Катагенез — путь развития группы организмов, связанный с резким упрощением строения и образа жизни. | общая дегенерация — общее упрощение строения | редукция органов зрения у обитателей почвы и пещер; редукция пищеварительной и выделительной системы у ленточных червей. |

Биологический регресс
Биологический регресс — отставание темпов эволюции группы от скорости изменения внешней среды.
Биологический регресс может привести к вымиранию группы.
Критерии биологического регресса:
- снижение численности особей
- уменьшение видового разнообразия
- сужение ареала обитания
В состоянии биологического регресса в настоящее время находятся крупные млекопитающие, такие, как уссурийский тигр, гепард, белый медведь, и целые группы животных — китообразные, амфибии, человекообразные обезьяны (кроме людей).
Закон Северцова
В эволюции всех групп организмов за периодом арогенеза всегда следует период возникновения частных приспособлений — аллогенез.
Этот закон может быть выведен из теории естественного отбора. Если сравнить частоту возникновения арогенезов и аллогенезов, то можно заметить, что первые характерны для возникновения крупных групп организмов в эволюции — типов, отделов, отдельных отрядов, иногда семейств. Другими словами, арогенезы появляются значительно реже, чем аллогенезы (определяющие появление отдельных видов, родов).

Таким образом, стегоцефалы путем арогенеза дали рептилий, а путем аллогенезов — современных амфибий. Группа безногих амфибий приобрела облик червеобразных форм, лишенных конечностей и хвоста (червяга). Хвостатые частично сохраняют пожизненные жабры, малоподвижные конечности и хорошо приспособленный к плавательным функциям хвост (тритоны). Бесхвостые амфибии приобрели сильные подвижные (в особенности задние) конечности (лягушки). Эта последняя группа пошла по пути завоевания суши, конечно, в пределах возможного, т. е. не слишком далеко от водоемов и во влажных лесах. Все эти формы экологически разошлись, конкуренция стала слабее, а биологический потенциал повысился.
Аллогенезы могут сменяться также катагенезом, и тогда биологический прогресс достигается благодаря морфофизиологическому регрессу. Например, существует паразит крабов — саккулина — который и сам является ракообразным, однако имеет вид мешка, набитого половыми продуктами, который ветвится и пронизывает тело хозяина. Трудно представить, что их предок относится к усоногим ракам, но в результате паразитического существования утратил почти все органы.

Паразитический рак саккулина и его планктонная личинка.
- Необратимость эволюционного процесса.
Например, при переходе наземных животных в водную среду изменяется строение конечностей. При этом во внутреннем строении плавников дельфина, кита сохранены признаки пятипалой конечности млекопитающих. Так как мутация приводит к обновлению генофонда популяции, она никогда не повторяет генофонд прошлого поколения. Так, если на каком-то этапе от примитивных земноводных возникли пресмыкающиеся, то пресмыкающиеся не могут вновь дать начало земноводным.
Другой пример — иглица. Листья у иглицы исчезли еще в древности, в процессе приспособления к засухе. Затем при переходе опять во влажную среду обитания вместо листьев у них появились ветви, похожие на листья.

- Скорость эволюционного процесса в разных группах организмов и в разные периоды времени различная.
В течение нескольких сотен миллионов лет на Земле существуют в неизменном виде «живые ископаемые» — скорпионы, кистеперые рыбы, гаттерия. Однако, на Филиппинах и в Австралии за 800 тыс. лет появилось несколько новых родов грызунов. Приблизительно за 20 млн. лет на Байкале возникло 240 видов раков, принадлежащих к 34 новым родам. Темпы эволюции снижаются и замедляются в стабильных условиях среды (океанические впадины, пещерные озера). На островах, где мало хищников, естественный отбор идет очень медленно. Наоборот, где проходит интенсивный отбор, эволюция также протекает быстрее. Например, в 30-х годах XX в. против вредителей использовали ядовитый препарат (ДДТ). Через несколько лет появились устойчивые к препарату формы, которые быстро распространились на Земле. Широкое применение антибиотиков — пенициллина, стрептомицина, грамицидина в середине XX в. привело к появлению устойчивых форм микроорганизмов. - Новые виды образуются не из высокоразвитых и специализированных, а, напротив, из относительно простых, неспециализированных форм. Наиболее специализированные формы оказываются наименее пластичными в изменяющихся условиях и более подвержены вымиранию
- Эволюция не всегда идет от простого к сложному. Существует много примеров «регрессивной» эволюции, когда сложная форма давала начало более простым. Большинство паразитов произошло от свободноживущих предков, которые имели более сложную организацию, чем современные формы; такие бескрылые птицы, как казуары, произошли от птиц, которые были способны летать; многие бескрылые насекомые произошли от крылатых; безногие змеи — от рептилий, имевших конечности; киты, не имеющие задних конечностей, — от четвероногих млекопитающих. Все это связано со случайным характером мутаций и с тем, что они не обязательно вызывают изменения от простого к сложному и от несовершенного к совершенному. Если какому-либо виду выгодно иметь более простое строение или вовсе лишиться какого-либо органа, то любые мутации, происходящие в этом направлении, будут накапливаться естественным отбором.
- Происходит чередование главных направлений эволюции: арогенеза и аллогенеза
-
Дивергенция — процесс расхождения признаков в процессе адаптации к разным условиям существования.
При дивергенции сходство между организмами объясняется общностью их происхождения, а различия — приспособлением к разным условиям среды. Примером дивергенции форм является возникновение разнообразных по морфофизиологическим особенностям вьюрков от одного или немногих предковых видов на Галапагосских островах. Расхождение внутривидовых форм и видов по разным местообитаниям определяется конкуренцией в борьбе за одинаковые условия, выход из которых и заключается в расселении по разным экологическим нишам.
Дивергенция может быть одним из путей видообразования, когда в результате действия элементарных эволюционных факторов популяции и группы популяций приобретают и сохраняют признаки, все более заметно отличающие их от родительского вида, что может приводить к распаду исходного вида на два и более дочерних. В конечном счете дивергенция приводит к формированию более крупных таксонов — родов, семейств и т.д. — которые продолжают расходиться.
Результат дивергенции: гомологичные органы у родственных форм приобретают разные функции и разное строение
-
Конвергенция — процесс эволюционного развития неродственных групп в сходном направлении и приобретение ими сходных признаков в процессе адаптации к одинаковым условиям среды.
Классическим примером конвергентного развития считается возникновение сходных форм тела у акуловых (первичноводные формы), ихтиозавров и китообразных (вторичноводные формы). При конвергентном развитии сходство между неродственными организмами бывает всегда только внешним (эволюционным изменениям в одном направлении подвергаются внешние признаки как результат приспособления к одинаковым условиям среды). По форме тела акула и дельфин сходны, но по таким существенным чертам, как строение кожных покровов, черепа, мускулатуры, кровеносной системы, дыхательной и других систем, эти группы позвоночных различны.
При конвергентном способе эволюции возникают аналогичные органы.
Сверху вниз: акула, ихтиозавр, дельфин.
Конвергентное сходство животных, питающихся муравьями.конвергенция
- Параллелизм (параллельное развитие) — такое явление, когда сходство признаков возникает в результате сходного (но независимого) изменения гомологичных структур.



Причиной параллелизма, по-видимому, является высокая вероятность похожих мутаций одних и тех же генов у разных видов. То же явление в рамках генетики описывает закон гомологических рядов наследственной изменчивости Н.И. Вавилова. Согласно этому закону, родственные виды характеризуются сходными рядами наследственной изменчивости. Поэтому сходные признаки могут возникать у родственны видов независимо — параллельно. Так, некоторые виды бабочек, принадлежащие к разным родам, имеют глазки на крыльях, а их ближайшие родственники глазков не имеют. Этот признак независимо возникал у разных видов, а не был унаследован от общих предков. От общих предков все бабочки унаследовали гены, мутации которых приводят к образованию глазков.
Другой пример того же рода — независимое возникновение саблезубых форм у млекопитающих. Широко известны вымершие саблезубые кошки, в частности, саблезубый тигр. Саблезубые кошки — это подсемейство семейства кошачьих, но саблезубость встречалась и в других отрядах млекопитающих, ныне вымерших (например, псевдо-саблезубые кошки). А в палеонтологической летописи Австралии есть сумчатый саблезубый волк. Таким образом, один и тот же признак в разных группах млекопитающих возник независимо.


Черепа сумчатого волка тилакосмила (слева) и саблезубой кошки смилодона (справа).
Вид, его критерии
Основатель современной систематики К. Линней рассматривал вид как группу сходных по морфологическим признакам организмов, которые свободно скрещиваются между собой. По мере развития биологии были получены свидетельства того, что различия между видами намного глубже, и затрагивают химический состав и концентрацию веществ в тканях, направление и скорость химических реакций, характер и интенсивность процессов жизнедеятельности, число и форму хромосом, т. е. вид является наименьшей группой организмов, отражающей их близкое родство. К тому же виды не существуют вечно — они возникают, развиваются, дают начало новым видам и исчезают.
Вид — это совокупность особей, сходных по строению и особенностям процессов жизнедеятельности, имеющих общее происхождение, свободно скрещивающихся между собой в природе и дающих плодовитое потомство.
Все особи одного вида имеют одинаковый кариотип и занимают в природе определенную географическую область — ареал.
Признаки сходства особей одного вида называют критериями вида. Так как ни один из критериев не является абсолютным, для правильного определения вида необходимо использовать совокупность критериев.
Основными критериями вида являются морфологический, физиологический, биохимический, экологический, географический, этологический (поведенческий) и генетический.
- Морфологический — совокупность внешних и внутренних признаков организмов одного вида. Несмотря на то, что у некоторых видов существуют уникальные признаки, с помощью одних только морфологических черт часто очень трудно различить близкородственные виды. Так, в последнее время открыт ряд видов-двойников, обитающих на одной территории, например домовая и курганчиковая мыши, поэтому использовать исключительно морфологический критерий для определения вида недопустимо.
- Физиологический — сходство процессов жизнедеятельности у организмов, в первую очередь, размножения. Он также не является универсальным, поскольку некоторые виды в природе скрещиваются и дают плодовитое потомство.
- Биохимический — сходство химического состава и протекания процессов обмена веществ. Несмотря на то, что данные показатели могут значительно варьировать у разных особей одного вида, в настоящее время им уделяется большое внимание, поскольку особенности строения и состава биополимеров помогают идентифицировать виды даже на молекулярном уровне и устанавливать степень их родства.
- Экологический — различие видов по их принадлежности к определенным экосистемам и экологическим нишам, которые они занимают. Однако многие неродственные виды занимают сходные экологические ниши, поэтому данный критерий может быть использован для выделения вида только в совокупности с другими признаками.
- Географический — существование популяции каждого вида в определенной части биосферы — ареале, который отличается от ареалов всех остальных видов. В связи с тем, что у множества видов границы ареалов совпадают, а также имеется ряд видов-космополитов, ареал которых охватывает огромные пространства, географический критерий также не может служить маркерным «видовым» признаком.
- Генетический — постоянство признаков хромосомного набора — кариотипа — и нуклеотидного состава ДНК у особей одного вида. В связи с тем, что негомологичные хромосомы не могут конъюгировать во время мейоза, потомство от скрещивания особей разных видов с неодинаковым набором хромосом либо не появляется вовсе, либо не плодовито. Это создает репродуктивную изоляцию вида, поддерживает его целостность и обеспечивает реальность существования в природе. Данное правило может нарушаться в случае скрещивания близких по происхождению видов с одинаковым кариотипом или возникновения различных мутаций, однако исключение лишь подтверждает общее правило, и виды следует рассматривать как устойчивые генетические системы. Генетический критерий является основным в системе критериев вида, но также не исчерпывающим.
При всей сложности системы критериев вид нельзя представить как группу абсолютно идентичных по всем параметрам организмов, то есть клонов. Наоборот, для многих видов характерно значительное разнообразие даже внешних признаков, как, например, для одних популяций божьих коровок характерно преобладание в окраске красного цвета, а для других — черного.
Популяция — структурная единица вида и элементарная единица эволюции
Трудно представить, чтобы в реальности особи одного вида были равномерно распределены по земной поверхности в пределах ареала, поскольку, например, лягушка озерная обитает в основном в достаточно редких стоячих пресных водоемах, и вряд ли ее можно встретить на полях и в лесах. Виды в природе чаще всего распадаются на отдельные группы, в зависимости от подходящих по совокупности условий местам обитания — популяции.
Популяция — группа особей одного вида, занимающих часть его ареала, свободно скрещивающихся между собой и относительно обособленных от других совокупностей особей того же вида в течение более или менее длительного времени.
Популяции могут быть разделены не только пространственно, они могут обитать даже на одной территории, но иметь различия в пищевых предпочтениях, сроках размножения и т. д..
Таким образом, вид — это совокупность популяций особей, обладающих рядом общих морфологических, физиологических, биохимических признаков и типов взаимоотношений с окружающей средой, населяющих определенный ареал, а также способных скрещиваться между собой с образованием плодовитого потомства, но почти или совсем не скрещивающихся с другими группами особей того же вида.
Внутри видов с большими ареалами, охватывающими территории с разными условиями жизни, иногда различают и подвиды — большие популяции или группы соседних популяций, имеющих стойкие морфологические отличия от других популяций.
Популяции разбросаны по земной поверхности не случайным образом, они привязаны к конкретным ее участкам. Совокупность всех факторов неживой природы, необходимых для проживания особей данного вида, называется местообитанием. Однако только этих факторов может быть недостаточно для занятия этого участка популяцией, поскольку она должна быть еще вовлечена в тесное взаимодействие с популяциями других видов, то есть занять определенное место в сообществе живых организмов — экологическую нишу. Так, австралийский сумчатый медведь коала при всех прочих равных условиях не может существовать без своего основного источника питания — эвкалипта.
Образующие неразрывное единство в одних и тех же местообитаниях популяции различных видов обыкновенно обеспечивают более или менее замкнутый круговорот веществ и являются элементарными экологическими системами (экосистемами) — биогеоценозами.
При всей своей требовательности к условиям окружающей среды популяции одного вида неоднородны по занимаемой площади, численности, плотности и пространственному размещению особей, часто образующих более мелкие группы (семьи, стаи, стада и др.), полу, возрасту, генофонду и т. д., поэтому различают их размерную, возрастную, половую, пространственную, генетическую, этологическую и другие структуры, а также динамику.
Важными характирестиками популяции являются генофонд — совокупность генов, характерных для особей данной популяции или вида, а также частоты определенных аллелей и генотипов. Разные популяции одного и того же вида изначально имеют неодинаковый генофонд, так как новые территории осваивают особи со случайными, а не специально подобранными генами. Под действием внутренних и внешних факторов генофонд подвергается еще более существенным изменениям: обогащается за счет возникновения мутаций и нового сочетания признаков и обедняется в результате выпадения отдельных аллелей при гибели или миграции некоторого числа особей.
Новые признаки и их сочетания могут быть полезными, нейтральными или вредными, поэтому в популяции выживают и успешно размножаются только приспособленные к данным условиям окружающей среды особи. Однако в двух различных точках земной поверхности условия окружающей среды никогда не бывают полностью идентичными, поэтому и направление изменений даже в двух соседних популяциях может быть совершенно противоположным или они будут протекать с различной скоростью. Результатом изменений генофонда является расхождение популяций по морфологическим, физиологическим, биохимическим и другим признакам. Если популяции при этом еще и изолированы друг от друга, то они могут давать начало новым видам.
Так, возникновение каких-либо препятствий в скрещивании особей различных популяций одного вида, например, вследствие образования горных массивов, изменения русел рек, различий в сроках размножения и т. д., приводит к тому, что популяции постепенно приобретают все больше различий и, в конце концов, становятся различными видами. Некоторое время на границах этих популяций происходит скрещивание особей и возникают гибриды, однако со временем и эти контакты исчезают, т. е. популяции из открытых генетических систем становятся закрытыми.
Несмотря на то, что действию факторов окружающей среды подвергаются в первую очередь отдельные особи, изменение генетического состава у единичного организма является незначительным и проявится в лучшем случае только у его потомков. Подвиды, виды и более крупные таксоны также не подходят на роль элементарных единиц эволюции, поскольку они не отличаются морфологическим, физиологическим, биохимическим, экологическим, географическим и генетическим единством, тогда как популяции как наименьшие структурные единицы вида, накапливающие разнообразие случайных изменений, худшие из которых будут отсеяны, соответствуют этому условию и являются элементарными единицами эволюции.
Микроэволюция
Изменение генетической структуры популяций не всегда приводит к образованию нового вида, а может лишь улучшать приспособление популяции к конкретным условиям среды, однако виды не являются вечными и неизменными — они способны развиваться. Этот процесс необратимого исторического изменения живого называется эволюцией. Первично эволюционные преобразования происходят внутри вида на уровне популяций. В их основе лежат, прежде всего, мутационный процесс и естественный отбор, приводящие к изменению генофонда популяций и вида в целом, или даже к образованию новых видов. Совокупность этих элементарных эволюционных событий называется микроэволюцией.
Популяции характеризуются огромным генетическим разнообразием, которое часто не проявляется фенотипически. Генетическое разнообразие возникает вследствие спонтанного мутагенеза, который происходит непрерывно. Большинство мутаций неблагоприятны для организма и снижают жизнеспособность популяции в целом, но, если они рецессивны, то могут сохраняться в гетерозиготе в течение продолжительного времени. Некоторые мутации, не имеющие приспособительной ценности в данных условиях существования, способны приобрести такую ценность в будущем или при освоении новых экологических ниш, создавая, таким образом, резерв наследственной изменчивости.
Значительное влияние на микроэволюционные процессы оказывают колебания численности особей в популяциях, миграции и катастрофы, а также изоляция популяций и видов.
Новый вид является промежуточным результатом эволюции, но никак не ее итогом, поскольку на этом микроэволюция не прерывается — она продолжается далее. Возникающие новые виды в случае удачного сочетания признаков заселяют новые местообитания, и, в свою очередь, дают начало новым видам. Такие группы близкородственных видов объединяются в роды, семейства и т. д. Эволюционные процессы, происходящие в надвидовых группах, называются уже макроэволюцией. В отличие от макроэволюции, микроэволюция протекает в гораздо более сжатые сроки, тогда как первой требуются десятки и сотни тысяч и миллионов лет, как, например, эволюция человека.
В результате микроэволюции формируется все многообразие видов живых организмов, когдалибо существовавших и ныне живущих на Земле.
Вместе с тем эволюция необратима, и уже исчезнувшие виды никогда не возникают вновь. Появляющиеся виды закрепляют все достигнутое в процессе эволюции, однако это не гарантирует того, что в будущем не появятся новые виды, которые будут иметь более совершенные приспособления к условиям окружающей среды.
Образование новых видов
В широком смысле под образованием новых видов понимается не только отщепление от основного ствола вида нового или распад материнского вида на несколько дочерних, а и общее развитие вида как целостной системы, приводящее к существенным изменениям его морфоструктурной организации. Однако чаще все же видообразование рассматривают как процесс формирования новых видов посредством разветвления «родословного древа» вида.
Принципиальное решение проблемы видообразования было предложено Ч. Дарвином. Согласно его теории расселение особей одного вида приводит к образованию популяций, которые вследствие различий условий окружающей среды вынуждены адаптироваться к ним. Это, в свою очередь, влечет за собой обострение внутривидовой борьбы за существование, направляемой естественным отбором. В настоящее время считается, что борьба за существование вовсе не является обязательным фактором видообразования, наоборот, давление отбора в ряде популяций может снижаться. Различие условий существования способствует возникновению неодинаковых приспособительных изменений в популяциях вида, следствием которого является расхождение признаков и свойств популяций — дивергенция.
Однако накопления различий, даже на генетическом уровне, отнюдь не достаточно для появления нового вида. До тех пор, пока различающиеся по каким-либо признакам популяции не только контактируют, но и способны к скрещиванию с образованием плодовитого потомства, они относятся к одному виду. Лишь невозможность перетока генов из одной группы особей в другую, даже в случае разрушения разделяющих их преград, т. е. скрещивания, означает завершение сложнейшего эволюционного процесса формирования нового вида.
Видообразование является продолжением микроэволюционных процессов. Существует точка зрения, что видообразование не может быть сведено к микроэволюции, оно представляет качественный этап эволюции и осуществляется благодаря другим механизмам.
Способы видообразования
Выделяют два основных способа видообразования: аллопатрическое и симпатрическое.
Аллопатрическое, или географическое видообразование является следствием пространственного разделения популяций физическими преградами (горные хребты, моря и реки) вследствие их возникновения или расселения в новые места обитания (географическая изоляция). Поскольку в данном случае генофонд отделившейся популяции существенно отличается от материнской, да и условия в месте ее обитания не будут совпадать с исходными, со временем это приведет к дивергенции и формированию нового вида. Ярким примером географического видообразования является обнаруженное Ч. Дарвином во время путешествия на корабле «Бигль» разнообразие видов вьюрков на Галапагосских островах вблизи побережья Эквадора. По-видимому, отдельные особи единственного обитающего на южноамериканском континенте вьюрка каким-то образом попали на острова, и, вследствие различия условий (в первую очередь доступности пищи) и географической изоляции, они постепенно эволюционировали, образовав группу родственных видов.
В основе симпатрического, или биологического видообразования лежит какая-либо из форм репродуктивной изоляции, при этом новые виды возникают внутри ареала исходного вида. Обязательным условием симпатрического видообразования является быстрая изоляция образовавшихся форм. Это более быстрый процесс, чем аллопатрическое видообразование, а новые формы похожи на исходные предковые.
Симпатрическое видообразование может быть вызвано быстрыми изменениями хромосомного набора (полиплоидизация) или хромосомными перестройками. Иногда новые виды возникают вследствие гибридизации двух исходных видов, как, например, у домашней сливы, являющейся гибридом терна и алычи. В некоторых случаях симпатрическое видообразование связано с разделением экологических ниш у популяций одного вида в пределах единого ареала или сезонной изоляции — расхождения сроков репродукции у растений (разные виды сосны в Калифорнии пылят в феврале и апреле) и сроков размножения у животных.
Из всего многообразия вновь возникающих видов только немногие, наиболее приспособленные, могут существовать длительное время и дать начало новым видам. Причины гибели большинства видов до сих пор неизвестны, скорее всего это обусловлено резкими изменениями климата, геологическими процессами и вытеснением их более приспособленными организмами. В настоящее время одной из причин гибели значительного числа видов является человек, который истребляет наиболее крупных животных и самые красивые растения, и если в XVII веке этот процесс только начался истреблением последнего тура, то в XXI веке ежечасно исчезает более 10 видов.
Сохранение многообразия видов как основа устойчивости биосферы
Несмотря на то, что на планете, по разным оценкам, обитает 5–10 млн еще не описанных видов организмов, о существовании большинства из них мы никогда не узнаем, поскольку ежечасно с лица Земли исчезает около 50 видов. Исчезновение живых организмов в настоящее время совсем не обязательно связано с их физическим истреблением, чаще это обусловлено разрушением в результате человеческой деятельности их природных мест обитания. Гибель отдельного вида вряд ли способна привести к фатальным последствиям для биосферы, однако уже давно установлено, что вымирание одного вида растений влечет за собой гибель 10–12 видов животных, а это уже представляет угрозу как для существования отдельных биогеоценозов, так и для глобальной экосистемы в целом.
Накопленные за предыдущие десятилетия печальные факты вынудили Международный союз охраны природы и природных ресурсов (МСОП) начать в 1949 году сбор информации о редких и находящихся под угрозой исчезновения видах растений и животных. В 1966 году МСОП издал первую «Красную книгу фактов».
Красная книга — это официальный документ, содержащий регулярно обновляемые данные о состоянии и распространении редких и находящихся под угрозой исчезновения видов растений, животных и грибов.
В этом документе принята пятиступенчатая шкала статуса охраняемого вида, причем к первой ступени охраны относят виды, спасение которых невозможно без осуществления специальных мероприятий, а к пятой — восстановленные виды, состояние которых благодаря принятым мерам не вызывает опасений, но они еще не подлежат промышленному использованию. Разработка такой шкалы позволяет направить первоочередные усилия в области охраны именно на самые редкие виды, такие как амурские тигры.
Помимо международного варианта Красной книги, существуют также национальные и региональные ее варианты. В СССР Красная книга была учреждена в 1974 году, а в Российской Федерации порядок ее ведения регламентируется Федеральными законами «Об охране окружающей среды», «О животном мире» и Постановлением Правительства РФ «О Красной книге Российской Федерации». Сегодня в Красную книгу РФ занесено 610 видов растений, 247 видов животных, 42 вида лишайников и 24 вида грибов. Популяции некоторых из них, в свое время оказавшиеся под угрозой исчезновения (бобр европейский, зубр), уже довольно успешно восстановлены.
Под охрану в России взяты следующие виды животных: русская выхухоль, тарбаган (монгольский сурок), белый медведь, кавказская европейская норка, калан, манул, амурский тигр, леопард, снежный барс, сивуч, морж, тюлени, дельфины, киты, лошадь Пржевальского, кулан, розовый пеликан, фламинго обыкновенный, аист черный, лебедь малый, орел степной, беркут, журавль черный, стерх, дрофа, филин, чайка белая, черепаха средиземноморская, полоз японский, гюрза, жаба камышовая, минога каспийская, все виды осетровых рыб, лосось озерный, жук-олень, шмель необыкновенный, аполлон обыкновенный, рак-богомол, жемчужница обыкновенная и др.
К растениям Красной книги РФ относятся 7 видов подснежников, некоторые виды полыни, женьшень настоящий, 7 видов колокольчиков, дуб зубчатый, пролеска пролесковидная, 11 видов касатиков, рябчик русский, тюльпан Шренка, лотос орехоносный, башмачок венерин настоящий, пион тонколистный, ковыль перистый, первоцвет Юлии, прострел (сон-трава) луговой, красавка белладонна, сосна пицундская, тис ягодный, щитовник китайский, полушник озерный, сфагнум мягкий, филлофора курчавая, хара нитевидная и др.
Редкие грибы представлены трюфелем летним, или русским черным трюфелем, трутовиком лакированным и др.
Охрана редких видов в большинстве случаев связана с запретом их уничтожения, сохранением их самих в искусственно созданной среде обитания (зоопарках), охране их местообитаний и созданием низкотемпературных генетических банков.
Наиболее эффективной мерой охраны редких видов является сохранение их местообитаний, которое достигается организацией сети особо охраняемых заповедных территорий, имеющих, в соответствии с Федеральным законом «Об особо охраняемых природных территориях» (1995), международное, федеральное, региональное или местное значение. К ним относятся государственные природные заповедники, национальные парки, природные парки, государственные природные заказники, памятники природы, дендрологические парки, ботанические сады и др.
Государственный природный заповедник — это полностью изъятый из хозяйственного использования особо охраняемый природный комплекс (земля, водные объекты, недра, растительный и животный мир), имеющий природоохранное, научное, эколого-просветительское значение как образец естественной природной среды, типичные или редкие ландшафты, места сохранения генетического фонда растительного и животного мира.
Заповедники, которые входят в международную систему биосферных резерватов, осуществляющих глобальный экологический мониторинг, имеют статус государственных природных биосферных заповедников. Заповедник является природоохранным, научно-исследовательским и эколого- просветительским учреждением, имеющим целью сохранение и изучение естественного хода природных процессов и явлений, генетического фонда растительного и животного мира, отдельных видов и сообществ растений и животных, типичных и уникальных экологических систем.
В настоящее время в России насчитывается около 100 государственных природных заповедников, 19 из которых носят статус биосферного, в т. ч. Байкальский, Баргузинский, Кавказский, «Кедровая падь», Кроноцкий, Приокско-Террасный и др.
В отличие от заповедников, территории (акватории) национальных парков включают в себя природные комплексы и объекты, имеющие особую экологическую, историческую и эстетическую ценности, и предназначены для использования в природоохранных, просветительских, научных и культурных целях и для регулируемого туризма. Такой статус имеют 39 особо охраняемых природных территорий, в т. ч. Забайкальский и Сочинский национальные парки, а также национальные парки «Куршская коса», «Русский Север», «Шушенский бор» и др.
Природные парки являются природоохранными рекреационными учреждениями, находящимися в ведении субъектов Российской Федерации, территории (акватории) которых включают в себя природные комплексы и объекты, имеющие значительную экологическую и эстетическую ценности, и предназначены для использования в природоохранных, просветительских и рекреационных целях.
Государственными природными заказниками являются территории (акватории), имеющие особое значение для сохранения или восстановления природных комплексов или их компонентов и поддержания экологического баланса.
Развитие эволюционных идей. Значение эволюционной теории Ч. Дарвина.
Взаимосвязь движущих сил эволюции. Формы естественного отбора, виды борьбы
за существование. Синтетическая теория эволюции. Элементарные факторы эволюции.
Исследования С. С. Четверикова. Роль эволюционной теории в формировании современной
естественнонаучной картины мира
Развитие эволюционных идей
Все теории происхождения и развития органического мира могут быть сведены к трем основным направлениям: креационизму, трансформизму и эволюционизму. Креационизм — это концепция постоянства видов, рассматривающая многообразие органического мира как результат его творения Богом. Данное направление сформировалось в результате установления господства христианской церкви в Европе, опиравшейся на библейские тексты. Видными представителями креационизма были К. Линней и Ж. Кювье.
«Князь ботаников» К. Линней, открывший и описавший сотни новых видов растений, и создавший первую стройную их систему, тем не менее, доказывал, что общее количество видов организмов неизменно со времени сотворения Земли, то есть они не только не появляются вновь, но и не исчезают. Только к концу жизни он пришел к заключению, что делом рук Божьих являются роды, тогда как виды могут развиваться вследствие приспособления к местным условиям.
Вкладом выдающегося французского зоолога Ж. Кювье (1769–1832) в биологию было опиравшееся на многочисленные данные палеонтологии, сравнительной анатомии и физиологии учение о корреляциях — взаимосвязях частей организма. Благодаря этому стало возможным реконструировать по отдельным частям внешний облик животного. Однако в процессе палеонтологических исследований Ж. Кювье не мог не обратить внимания как на явное изобилие ископаемых форм, так и на резкие изменения групп животных в течение геологической истории. Эти данные послужили отправной точкой для формулировки теории катастроф, согласно которой все или почти все организмы на Земле неоднократно погибали в результате периодических стихийных бедствий, а потом планета вновь заселялась видами, пережившими катастрофу. Последователи Ж. Кювье насчитывали до 27 таких катастроф в истории Земли. Соображения же об эволюции представлялись Ж. Кювье оторванными от реальности.
Противоречия в исходных положениях креационизма, которые становились все более очевидными по мере накопления научных фактов, послужили отправной точкой для формирования другой системы взглядов — трансформизма, признающего реальное существование видов и их историческое развитие. Представители данного течения — Ж. Бюффон, И. Гете, Э. Дарвин и Э. Жоффруа Сент-Илер, будучи не в силах вскрыть истинные причины эволюции, сводили их к приспособлению к условиям окружающей среды и наследованию приобретенных признаков. Корни трансформизма можно найти в работах еще древнегреческих и средневековых философов, которые признавали исторические изменения органического мира. Так, Аристотель высказал идею о единстве природы и постепенности перехода от тел неживой природы к растениям, а от них — к животным — «лестнице природы». Основной причиной изменений живых организмов он считал их внутреннее стремление к совершенству.
Французский естествоиспытатель Ж. Бюффон (1707–1788), главным трудом жизни которого была 36-томная «Естественная история», вопреки представлениям креационистов, раздвинул рамки истории Земли до 80–90 тыс. лет. При этом он констатировал единство растительного и животного мира, а также возможность изменения родственных организмов под воздействием факторов окружающей среды в результате одомашнивания и гибридизации.
Английский врач, философ и поэт Э. Дарвин (1731–1802), дед Ч. Дарвина, полагал, что история органического мира насчитывает миллионы лет, а разнообразие животного мира является результатом смешения нескольких «естественных» групп, влияния внешней среды, упражнения и неупражнения органов, и других факторов.
Единство плана строения групп животных считал одним из главных доказательств развития живого мира Э. Жоффруа Сент-Илер (1772–1844). Однако, в отличие от своих предшественников, он склонялся к тому, что изменение видов обусловлено воздействием факторов среды не на взрослые особи, а на зародыши.
Несмотря на то, что в разгоревшейся в 1831 году дискуссии между Ж. Кювье и Э. Жоффруа Сент-Илером в виде серии докладов в Академии наук, явный перевес остался на стороне первого, именно трансформизм стал предтечей эволюционизма. Эволюционизм (теория эволюции, эволюционное учение) — это система взглядов, признающая развитие природы по определенным законам. Он является теоретической вершиной биологии, которая позволяет объяснять наблюдаемые нами многообразие и сложность живых систем. Однако в связи с тем, что эволюционное учение описывает труднодоступные для наблюдения явления, оно сталкивается со значительными затруднениями. Иногда теорию эволюции называют «дарвинизмом» и отождествляют с учением Ч. Дарвина, что является в корне неверным, поскольку, хотя теория Ч. Дарвина и внесла неоценимый вклад в развитие не только эволюционного учения, но и биологии в целом (а также многих других наук), основы эволюционной теории были заложены другими учеными, она продолжает развиваться и по сей день, а «дарвинизм» во многих аспектах имеет лишь историческое значение.
Создателем первой эволюционной теории — ламаркизма — был французский естествоиспытатель Ж. Б. Ламарк (1744–1829). Движущей силой эволюции он считал внутреннее стремление организмов к совершенству (закон градации), однако приспособление к условиям внешней среды вынуждает их к отклонению от этой магистральной линии. При этом органы, которые интенсивно используются животным в процессе жизнедеятельности, развиваются, а ненужные ему, наоборот, ослабляются и даже могут исчезать (закон упражнения и неупражнения органов). Приобретенные в процессе жизни признаки закрепляются и передаются потомкам. Так, наличие перепонок между пальцами ног у водоплавающих птиц он объяснял попытками их предков передвигаться в водной среде, а длинная шея жирафов, по Ламарку, является следствием того, что их предки старались доставать листья с верхушек деревьев.
Недостатками ламаркизма были теоретичность многих построений, а также допущение вмешательства Творца в эволюцию. В процессе развития биологии выяснилось, что индивидуальные изменения, приобретаемые организмами в процессе жизни, большей частью укладываются в пределы фенотипической изменчивости, а их передача практически невозможна. Например, немецкий зоолог и теоретик эволюционного учения А. Вейсман (1834–1914) на протяжении многих поколений отрезал мышам хвосты и всегда получал в потомстве только хвостатых грызунов. Теория Ж. Б. Ламарка не была принята современниками, но на рубеже веков легла в основу так называемого неоламаркизма.
Значение эволюционной теории Ч. Дарвина
Предпосылками для создания наиболее известной эволюционной теории Ч. Дарвина, или дарвинизма, стали публикация в 1778 году труда английского экономиста Т. Мальтуса «Трактат о народонаселении», работы геолога Ч. Лайеля, формулировка клеточной теории, успехи селекции в Англии и собственные наблюдения Ч. Дарвина (1809–1882), сделанные в годы учебы в Кембридже, в экспедиции в качестве натуралиста на корабле «Бигль» и по ее завершении.
Так, Т. Мальтус утверждал, что население Земли увеличивается в геометрической прогрессии, что существенно превышает возможности планеты по обеспечению его продовольствием и ведет к гибели части потомства. Параллели, проведенные Ч. Дарвиным и его соавтором А. Уоллесом (1823–1913), указывали на то, что и в природе особи размножаются с очень большой скоростью, однако численность популяций остается относительно постоянной. Исследования английского геолога Ч. Лайеля позволили установить, что поверхность Земли далеко не всегда была такой, как в настоящее время, а ее изменения были вызваны воздействием воды, ветра, вулканическими извержениями и деятельностью живых организмов. Самого же Ч. Дарвина еще в студенческие годы поразила крайняя степень изменчивости жуков, а во время путешествия — сходство флоры и фауны континентальной Южной Америки и лежащих вблизи нее Галапагосских островов, и в то же время значительное разнообразие видов, например вьюрков и черепах. Кроме того, в экспедиции он мог наблюдать скелеты гигантских вымерших млекопитающих, сходных с современными броненосцами и ленивцами, что существенно поколебало его веру в сотворение видов.
Основные положения теории эволюции были высказаны Ч. Дарвином в 1859 году на заседании Лондонского Королевского общества, а впоследствии развиты в книгах «Происхождение видов путем естественного отбора, или сохранение благоприятствуемых пород в борьбе за жизнь» (1859), «Изменение домашних животных и культурных растений» (1868), «Происхождение человека и половой отбор» (1871), «Выражение эмоций у человека и животных» (1872) и др.
Сущность разработанной Ч. Дарвиным концепции эволюции можно свести к ряду вытекающих друг из друга положений, имеющих соответствующие доказательства:
Сущность разработанной Ч. Дарвиным концепции эволюции можно свести к ряду вытекающих друг из друга положений, имеющих соответствующие доказательства:
Основные ароморфозы в эволюции растений и животных
Растительные и животные организмы, представляющие различные ветви эволюции органического мира, в процессе исторического развития независимо приобретали определенные черты строения, которые будут охарактеризованы далее.
У растений важнейшими из них являются переход от гаплоидности к диплоидности, независимость от воды в процессе оплодотворения, переход от наружного оплодотворения к внутреннему и возникновение двойного оплодотворения, расчленение тела на органы, развитие проводящей системы, усложнение и усовершенствование тканей, а также специализация опыления с помощью насекомых и распространения семян и плодов.
Переход от гаплоидности к диплоидности сделал растения более устойчивыми к действию факторов окружающей среды вследствие снижения риска проявления рецессивных мутаций. Повидимому, это преобразование коснулось предков сосудистых растений, в число которых не входят моховидные, характеризующиеся преобладанием в жизненном цикле гаметофита.
Главные ароморфозы в эволюции животных связаны с возникновением многоклеточности и все большим расчленением всех систем органов, возникновением прочного скелета, развитием центральной нервной системы, а также общественного поведения в различных группах высокоорганизованных животных, что дало толчок и к прогрессу человека.
Макроэволюция. Направления и пути эволюции (А. Н. Северцов, И. И. Шмальгаузен).
Биологический прогресс и регресс, ароморфоз, идиоадаптация, дегенерация. Причины
биологического прогресса и регресса. Гипотезы возникновения жизни на Земле. Основные
ароморфозы в эволюции растений и животных. Усложнение живых организмов в процессе
эволюции
Макроэволюция
Образование вида ознаменовывает собой новый виток эволюционного процесса, поскольку особи этого вида, будучи более приспособленными к условиям среды, нежели особи материнского вида, постепенно расселяются на новые территории, и уже в его популяциях играют свою созидательную роль мутагенез, популяционные волны, изоляция и естественный отбор. Со временем эти популяции дают начало новым видам, которые вследствие генетической изоляции имеют гораздо больше признаков сходства между собой, чем с видами того рода, от которого отпочковался видродоначальник, и, таким образом, возникает новый род, затем — новое семейство, отряд (порядок), класс и т. д. Совокупность эволюционных процессов, которые приводят к возникновению надвидовых таксонов (родов, семейств, отрядов, классов и т. п.), называется макроэволюцией. Макроэволюционные процессы как бы обобщают микроэволюционные изменения, происходящие в течение длительного времени, выявляя при этом основные тенденции, направления и закономерности эволюции органического мира, которые не поддаются наблюдению на более низком уровне. До сих пор никаких специфических механизмов макроэволюции не выявлено, поэтому считается, что она осуществляется только посредством микроэволюционных процессов, однако эта позиция постоянно подвергается вполне обоснованной критике.
Возникновение сложной иерархической системы органического мира в значительной степени является результатом неодинаковой скорости эволюционирования различных групп организмов. Так, уже упоминавшийся гинкго двулопастный как бы «законсервировался» на тысячи лет, тогда как достаточно близкие к нему сосны существенно изменились за это время.
Направления и пути эволюции (А. Н. Северцов, И. И. Шмальгаузен). Биологический прогресс и регресс,
ароморфоз, идиоадаптация, дегенерация
Анализируя историю органического мира, можно заметить, что в определенные промежутки времени господствовали отдельные группы организмов, которые затем клонились к упадку либо исчезали вовсе. Таким образом, можно различить три магистральных направления эволюции: биологический прогресс, биологический регресс и биологическая стабилизация. Значительный вклад в разработку учения о направлениях и путях эволюции внесли русские эволюционисты А. Н. Северцов и И. И. Шмальгаузен.
Биологический прогресс связан с биологическим процветанием группы в целом и характеризует ее эволюционный успех. Он отражает закономерное развитие живой природы от простого к сложному, от более низкой степени организации к более высокой. По А. Н. Северцову, критериями биологического прогресса являются увеличение численности особей данной группы, расширение ее ареала, а также появление и развитие в ее составе групп низшего ранга (преобразование вида в род, рода в семейство и т. д.). В настоящее время биологический прогресс наблюдается у покрытосеменных растений, насекомых, костистых рыб и млекопитающих.
По А. Н. Северцову, биологический прогресс может достигаться вследствие определенных морфофизиологических преобразований организмов, при этом он выделил три основные пути достижения: арогенез, аллогенез и катагенез.
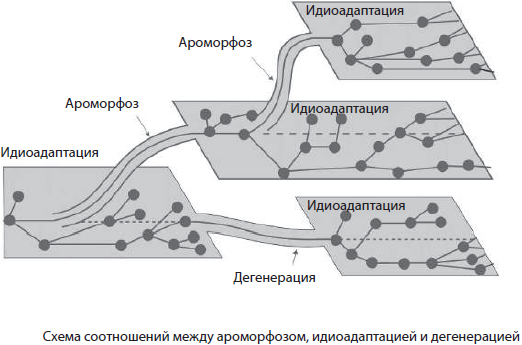
Арогенез, или морфофизиологический прогресс, связан со значительным расширением ареала данной группы организмов вследствие приобретения больших изменений строения — ароморфозов.
Ароморфозом называют эволюционное преобразование строения и функций организма, которое повышает его уровень организации и открывает новые возможности для приспособления к разнообразным условиям существования.
Примерами ароморфозов являются возникновение эукариотической клетки, многоклеточности, появление сердца у рыб и разделение его полной перегородкой у птиц и млекопитающих, формирование цветка у покрытосеменных и т. п.
Аллогенез, в отличие от арогенеза, не сопровождается расширением ареала, однако внутри старого возникает значительное разнообразие форм, имеющих частные приспособления к среде обитания — идиоадаптации.
Идиоадаптация — это мелкое морфофизиологическое приспособление к специальным условиям среды, полезное в борьбе за существование, но не изменяющее уровня организации. Эти изменения иллюстрируют покровительственная окраска у животных, разнообразие ротовых аппаратов у насекомых, колючки растений и др. Не менее удачным примером являются дарвиновы вьюрки, специализирующиеся на различных видах пищи, у которых преобразования вначале затронули клюв, а затем и другие части тела — оперение, хвост и т. п.
Как это ни парадоксально, но и упрощение организации может вести к биологическому прогрессу. Этот путь называется катагенезом.
Дегенерация — это упрощение организмов в процессе эволюции, которое сопровождается потерей определенных функций или органов.
Большей частью она характерна для паразитических организмов и животных, перешедших к прикрепленному способу жизни. Так, растение-паразит повилика европейская лишена хлорофилла, а ее листья редуцированы до чешуек, питается же она за счет поглощения питательных веществ из тела растения-хозяина с помощью специальных присосок. Паразитирующие в тонком кишечнике человека и других животных цепни и лентецы в процессе эволюции принесли свою пищеварительную систему и почти все органы чувств в жертву гипертрофированной половой системе, обеспечивающей необычайную плодовитость (до 11 млрд яиц в течение жизни). Сидячий образ жизни привел у взрослых представителей одной из групп хордовых — асцидий — к исчезновению не только органов чувств и пищеварительной системы, но и самой хорды, обнаруживающейся только у личинок.
Фаза биологического прогресса сменяется фазой биологической стабилизации, сущность которой заключается в сохранении признаков данного вида как наиболее благоприятных в данном микроокружении. По И. И. Шмальгаузену, она вовсе «не означает прекращения эволюции, наоборот, означает максимальную согласованность организма с изменениями среды». В фазе биологической стабилизации находятся «живые ископаемые» латимерия, гингко и др.
Антиподом биологического прогресса является биологический регресс — эволюционный упадок данной группы вследствие невозможности приспособиться к изменениям окружающей среды. Он проявляется в снижении численности популяций, сужении ареалов, уменьшении количества групп низшего ранга в составе высшего таксона. Группе организмов, которая пребывает в состоянии биологического регресса, угрожает вымирание. В истории органического мира можно увидеть много примеров такого явления, и в настоящее время регресс характерен для некоторых папоротников, амфибий и рептилий. С появлением человека биологический регресс зачастую обусловлен его хозяйственной деятельностью.
Направления и пути эволюции органического мира не являются взаимоисключающими, то есть появление ароморфоза не означает, что идиоадаптация или дегенерация произойти уже не может. Напротив, согласно разработанному А. Н. Северцовым и И. И. Шмальгаузеном правилу смены фаз, различные направления эволюционного процесса и пути достижения биологического прогресса закономерно сменяют друг друга. В ходе эволюции эти пути сочетаются: достаточно редкие ароморфозы переводят группу организмов на качественно новый уровень организации, а в дальнейшем историческое развитие идет по пути идиоадаптации либо дегенерации, обеспечивающих приспособление к конкретным условиям среды обитания.
Причины биологического прогресса и регресса
В процессе эволюции планку естественного отбора преодолевают и, соответственно, прогрессируют только те группы организмов, у которых наследственная изменчивость создает достаточное количество комбинаций, которые могут обеспечить выживание группы в целом.
Те же группы, у которых по каким-то причинам не имеется такого резерва, в большинстве случаев обречены на вымирание. Зачастую это связано с низким давлением отбора на предыдущих этапах эволюционного процесса, приведшим к узкой специализации группы или даже дегенеративным явлениям. Следствием этого является невозможность приспособиться к новым условиям среды при ее резких изменениях. Ярким примером тому является внезапная гибель динозавров вследствие падения гигантского небесного тела на Землю 65 млн лет назад, которое повлекло за собой землетрясение, поднятие в воздух миллионов тонн пыли, резкое похолодание, гибель большей части растений и растительноядных животных. В то же время предки современных млекопитающих, не имея узких предпочтений источников питания и будучи теплокровными, смогли пережить эти условия и занять на планете господствующее положение.
Пути макроэволюции. Биологический прогресс и регресс
Макроэволюция ― процесс образования из видов новых родов, из родов ― семейств, из семейств ― отрядов и т. д.
В отличие от микроэволюции, протекающей внутри популяции, макроэволюция ― эволюция надвидовая. Тем не менее, в макроэволюции действуют всё те же процессы: борьба за существование, естественный отбор.
Макроэволюция позвоночных
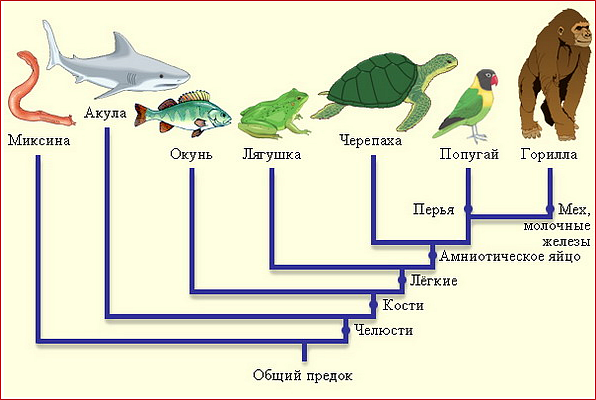
В общем смысле макроэволюцией можно назвать развитие жизни на Земле в целом, включая и ее происхождение.
Между микро- и макроэволюцией нельзя провести резкую грань, потому что процесс микроэволюции, первично вызывающий дивергенцию популяций (вплоть до видообразования), продолжается без какого-либо перерыва и на макроэволюционном уровне внутри вновь возникших форм.
Формы эволюции
Дивергенция – расхождение признаков в процессе эволюции, приводящее к образованию новых форм или таксонов организмов от общего предка.
Например, разные подвиды вьюрков на Галапагосских островах, конечности наземных и водных млекопитающих, ротовые аппараты разных типов у насекомых

Рисунок 1. Дивергенция вьюрков
Конвергенция – независимое возникновение подобных признаков у организмов, неродственных друг другу, или у органов, имеющих в эмбриональном развитии различное происхождение, но выполняющих сходные функции.
Например, роющие конечности крота и медведки; колючки кактусов (видоизменённые листья), белой акации (прилистники) и боярышника (видоизменённые побеги); жабры рыб и жабры личинок стрекоз.
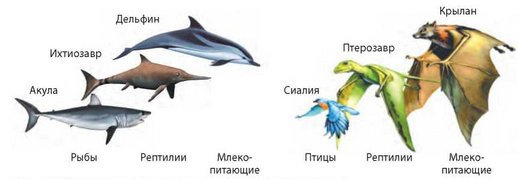
Рисунок 2. Конвергенция у водоплавающих (обтекаемая форма тела) и летающих (крылья) организмов
Параллелизм – процесс эволюционного развития в сходном направлении нескольких ранее дивергировавших групп.
Например, развитие саблезубости у кошачьих, появление ласт у китообразных и ластоногих, сходство млекопитающих тропических лесов, обитающих на разных континентах.

Рисунок 3. Параллелизм в форме тела котика, тюленя и моржа
Коэволюция – совместная эволюция биологических видов, взаимодействующих в экосистеме. Изменения, затрагивающие особей одного вида, приводят к изменениям в других видах.
Например, динамика развития системы «растение – гусеницы, которые его поедают». Растение производит ядовитые для гусениц вещества, но определенные виды гусениц в ходе эволюции приобрели нечувствительность к растительным ядам; более того, они накапливают их в своем теле и сами становятся несъедобными для птиц.
Ещё один яркий пример коэволюции – совместная эволюция цветковых растений и их опылителей (коэволюция при мутуализме). Цветок некоторых растений устроен таким образом, что достать нектар может только один определённый вид.

Рисунок 4. Пример коэволюции – колибри и цветковые
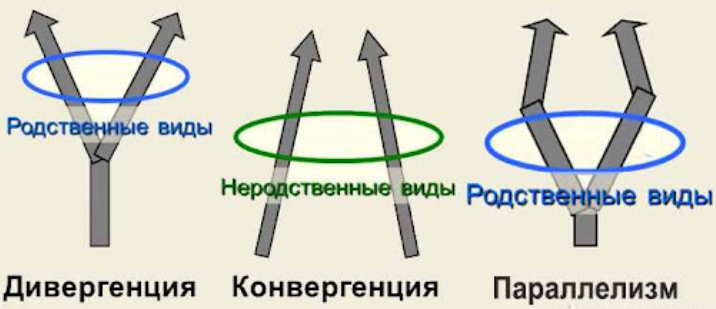
Рисунок 5. Формы эволюционного процесса
Основные направления эволюции
Изучением направлений эволюции занимался русский биолог А. Н. Северцов. Учение о прогрессе в эволюции было в дальнейшем развито его учеником И. И. Шмальгаузеном.
К основным направлениям эволюции относятся:
1. Биологический прогресс.
2. Биологический регресс.
Биологический прогресс
Биологический прогресс – возрастание приспособленности организмов к окружающей среде (по А. Н. Северцову).
Критерии биологического прогресса:
-
увеличение численности;
-
повышение видового разнообразия (прогрессивная дифференциация);
-
расширение ареала.
Механизм биологического прогресса:
возникновение новых приспособлений снижает гибель особей → средний уровень численности вида возрастает → увеличивается плотность населения → обостряется внутривидовая конкуренция + возрастает приспособленность → расширяется ареал → вид заселяет новые территории и вынужден приспосабливаться к новым условиям → отдельные популяции приобретают разные признаки (дивергенция признаков) → происходит образование дочерних таксонов.
Общая причина биологического прогресса – результат успеха биологической группы в борьбе за существование.
По мнению А.Н. Северцова биологический прогресс может быть достигнут тремя путями:
1. Ароморфозом
2. Идиоадаптацией
3. Дегенерацией
Внимание! В первой части экзамена можно встретить большое количество заданий на сравнение понятий ароморфоз/идиоадаптация/дегенерация. Встречаются задания на соответствие примера и одного из путей достижения биологического прогресса.
Ароморфозы
Ароморфоз – прогрессивное эволюционное изменение строения, приводящее к общему повышению уровня организации организмов.
Именно путем ароморфозов возникают крупные таксономические группы – отряды, классы, отделы и т.д.
С ароморфозами прежде всего связаны эволюционные преобразования кровеносной, дыхательной, нервной и других систем органов, оказывающих непосредственное влияние на увеличение интенсивности обмена веществ и энергии.
Ароморфозы формируются на основе наследственной изменчивости и естественного отбора и являются приспособлениями широкого значения. Они дают преимущества в борьбе за существование и открывают возможности освоения новой, прежде недоступной среды обитания.
Идиоадаптации
Идиоадаптации – частные приспособления, не носящие принципиального характера, но позволяющие преуспеть в определенной среде обитания.
Повышения уровня организации при этом не происходит. Благодаря идиоадаптации образуются мелкие таксономические группы: виды, роды, семейства.
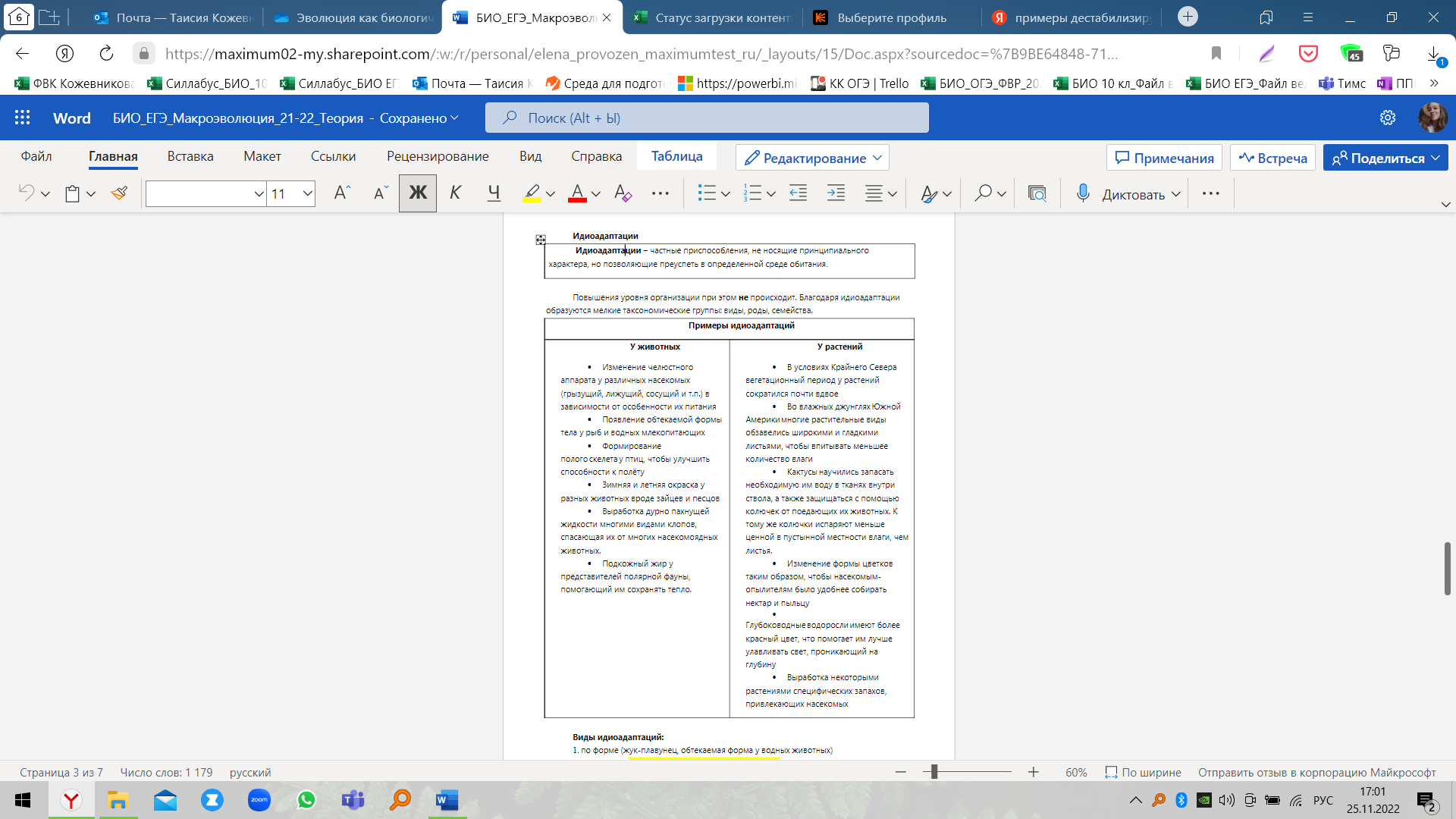
Виды идиоадаптаций:
1. по форме (жук-плавунец, обтекаемая форма у водных животных)
2. по окраске (см. подробнее “Результаты эволюции”)


3. по размножению (подбрасываемые кукушкой яиц в чужие гнёзда, забота о потомстве)
4. по передвижению (перепонки водоплавающих птиц, моржи, тюлени; трубчатые кости и киль у птиц).
5. приспособления к условиям окружающей среды (подкожный жир, линька и т. д.)
Дегенерация
Дегенерация – одно из направлений эволюционного процесса, связанное с упрощением организации, в том числе утратой органов и их систем.
В филогенезе А. Н. Северцов ввел понятия общей и частной дегенерации.
Общая дегенерация, по Северцову, связана с морфофизиологическим регрессом – упрощением организации. Происходит при переходе от активного образа жизни к более пассивному; к паразитическому и подземному образу жизни.
Так, малая подвижность и пассивный тип питания двустворчатых моллюсков привели к исчезновению головы, ленточные черви утратили пищеварительную систему.
Внимание! Не путайте морфофизиологический регресс и биологический регресс. Общая дегенерация – это путь достижения биологического прогресса.
При частной дегенерации у организмов в процессе их исторического развития редуцируются органы, имевшиеся у предков, например конечности у безногих ящериц, раковина у головоногих моллюсков. Причина редукции органов – отсутствие условий, необходимых для их развития и функционирования.
Внимание! Частная дегенерация – это пример идиоадаптации, не ведёт к снижению уровня организации.
Направления биологического прогресса
Биологический регресс
Биологический регресс – отставание темпов эволюции группы от скорости изменения внешней среды.
Регресс, как правило, ведёт к вымиранию видов. В последнее время виновником биологического регресса часто становится человек.
Критерии биологического регресса:
-
снижение численности особей
-
уменьшение видового разнообразия
-
сужение ареала
В состоянии биологического регресса в настоящее время находятся крупные млекопитающие, такие, как уссурийский тигр, гепард, белый медведь, и целые группы животных: китообразные, амфибии, человекообразные обезьяны (кроме людей).
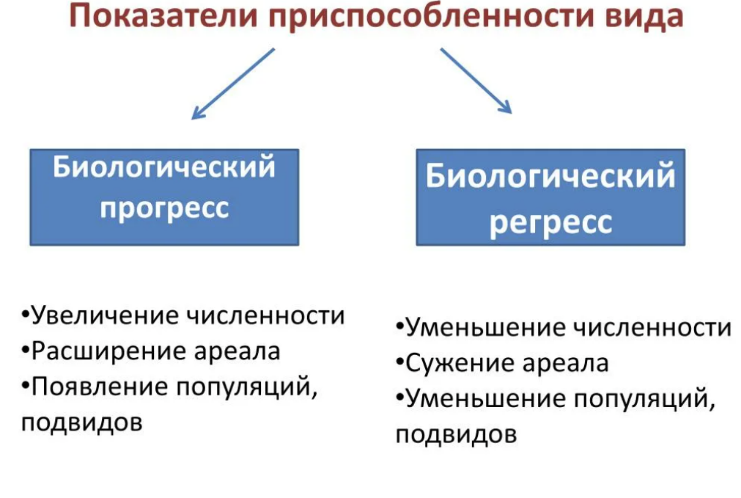
Учение о направлениях эволюционного процесса было разработано русским учёным А. Н. Северцовым.
Биологический прогресс — направление эволюции, характеризующееся повышением приспособленности систематической группы живых организмов к среде обитания.
Критерии биологического прогресса:
- увеличение численности особей;
- расширение ареала;
- увеличение числа таксонов (популяций, подвидов, видов и т. д.).
Биологический прогресс — это результат успеха систематической группы в борьбе за существование. Он обеспечивается появлением новых приспособлений, полезных в данной среде обитания. Организмы выживают и размножаются, что приводит к увеличению численности и освоению новых мест обитания. Возникают новые популяции. Они подвергаются действию разнонаправленного естественного отбора и постепенно превращаются в новые виды, виды — в роды, роды — в семейства и так далее. Происходит увеличение числа таксономических групп и их совершенствование.
В состоянии прогресса в настоящее время находятся многие сорняки (одуванчик, пырей, марь белая), вредители сельскохозяйственных культур (колорадский жук, фитофтора). Их прогресс связан с деятельностью человека.
Биологический прогресс достигается тремя путями: повышением организации и освоением новой среды обитания; приспособлением к новой среде и её заселением; понижением организации и освоением более простой среды.

Рис. (1). Пути достижения биологического прогресса
Эволюционное изменение, которое приводит к существенному усложнению строения организмов, повышению их уровня организации и позволяет освоить новую среду обитания, — ароморфоз.
Ароморфозы привели к возникновению крупных систематических групп: типов, отделов, классов, некоторых отрядов.
Примеры ароморфозов у животных:
- двусторонняя симметрия тела;
- сквозной кишечник;
- трахейное дыхание у членистоногих;
- лёгочное дыхание у позвоночных;
- альвеолярные лёгкие;
- четырёхкамерное сердце;
- два круга кровообращения;
- теплокровность.
Примеры ароморфозов у растений:
- возникновение фотосинтеза;
- формирование тканей;
- возникновение листа, стебля, корня;
- появление семени;
- образование цветка и плода.
Эволюционное изменение, которое приспосабливает организмы к условиям конкретной среды обитания, но не повышает их уровень организации, — идиоадаптация (алломорфоз).
Идиоадаптации возникают на основе ароморфозов и позволяют систематической группе более полно заселить среду обитания. Приводят к возникновению видов, родов, семейств.
Примеры идиоадаптаций:
- разные типы листьев и стеблей у растений;
- разное строение цветков у растений;
- видоизменения побега растений;
- разные клювы у птиц;
- разная форма тела и окраска рыб;
- разные типы ротовых аппаратов насекомых.

Рис. (2). Идиоадаптации ротовых органов насекомых
Упрощение строения организмов при переходе в более простую среду обитания — дегенерация.
Примеры общей дегенерации:
- упрощение нервной системы и органов чувств у паразитических червей;
- редукция пищеварительной системы у ленточных червей;
- редукция головы у двустворчатых моллюсков;
- исчезновение крыльев у некоторых паразитических насекомых;
- редукция листьев у паразитических растений.
|
|
|
|
Рис. (3). Растение-паразит повилика |
Рис. (4). Двустворчатые моллюски |


Биологический регресс — направление эволюции, характеризующееся понижением приспособленности систематической группы живых организмов к среде обитания и её постепенным вымиранием.
Критерии биологического регресса:
- снижение численности особей в систематических группах;
- сужение ареала;
- уменьшение числа таксонов (популяций, подвидов, видов и т. д.).
Виды, находящиеся в состоянии регресса, нуждаются в охране и занесены в Красную книгу.
Это уссурийский тигр, бурый и гималайский медведи, чёрный аист, венерин башмачок, женьшень и многие другие.
Источники:
Рис. 1. Пути достижения биологического прогресса. © ЯКласс.
Рис. 2. Идиоадаптации ротовых органов насекомых. © ЯКласс.
Рис. 3. Растение-паразит повилика. https://cdn.pixabay.com/photo/2017/09/23/01/50/entangled-2777693_960_720. 28.09.2021.
Рис. 4. Двустворчатые моллюски. https://cdn.pixabay.com/photo/2020/01/06/19/11/clam-4746021_960_720. 28.09.2021.