Шпаргалки — Цитология
Cytology_1
1. Клеточная теория.
2. Ядерно-цитоплазматический транспорт.
3. Ядерная оболочка. Её ультраструктура и роль.
4. Строение и функции ядерной поры.
5. Локализация хромосом в интерфазном ядре.
6. Полиплоидия и политения, их значение.
7. Ядерный белковый матрикс.
8. Уровни укладки ДНК в составе хромосом
9. Репликация ДНК в хромосомах эукариотов и прокариотов.
10. Структура хромосом.
Cytology_2
11. Доказательство непрерывности хромосом в течение клеточного цикла.
12. Строение ядрышка.
13. Строение рибосом, их состав и роль в синтезе белка.
14. Клеточный цикл, его стадии и способы их изучения.
15. Мейоз, последовательность фаз мейоза и его значение.
16. Кариотип, определение, методы изучения.
17. Цитотомия бактериальных, растительных и животных клеток.
18. Митоз, механизм движения хромосом в этом процессе.
Cytology_3
19. Типы митотического веретена, процесс их образования.
24. Вакуоли растений, строение и функции.
25. Вакуолярная система растений.
20. Кинетохор, его участие в делении клетки.
23. Пластиды высших и низших растений.
29. Строение и функции гладкого эндоплазматического ретикулума.
27. Проблема автономности хлоропластов и митохондрий.
28. Синтез белка.
30. Строение и функции гранулярного эндоплазматического ретикулума.
26. Ультраструктура митохондрий, функции.
32. Лизосомы, их классификация и строение.
31. Строение и функции аппарата Гольджи.
37. Рецепторная роль плазматической мембраны.
36. Структура плазматической мембраны клетки и способы ее изучения.
33. Развитие хлоропластов.
34. Синтез клеточных мембран
39. Проницаемость плазматической мембраны (пассивный и активный транспорт).
38. Проницаемость клеточных мембран (эндоцитоз и экзоцитоз).
22. Судьба органелл при митозе.
Cytology_4
41. Процесс образования клеточной стенки растений.
42. Строение и свойства клеточных растительных клеток и бактерий.
45. Центриоли, их строение и поведение в клеточном цикле.
46. Центриоли, их структура и воспроизведение.
40. Межклеточные контакты.
27. Проблема автономности хлоропластов и митохондрий.
35. Молекулярная организация клеточных мембран.
54. Отличия в строении клеток прокариот от эукариот.
Cytology_5
41. Процесс образования клеточной стенки.
51. Промежуточные филаменты.
55. Строение жгутиков бактерий.
42. Строение и свойства клеточных стенок растительных клеток и бактерий.
56. Метод авторадиографии.
44. Скелетно-двигательный аппарат клетки.
61. Вакуолярная система.
58. Методы электронно-микроскопического изучения клеток.
50. Микротрубочки интерфазной клетки, строение и функции.
47. Микрофиламенты.
Cytology_6
59. Апоптоз и некроз.
49. Моторные белки.
21. Периферический материал митотических хромосом.
52. Образование ресничек.
57. Иммуноцитохимический метод в клеточной биологии.
48. Сократимые структуры клетки.
60. Регуляция клеточного цикла.
1.Клеточная теория(КТ)
КТ- обобщ. предст-я о стр-и кл-к, как единиц живого, об их размножении и роли в форм-и однокл орг-мов.
КТ сформулирована 3мя немцами:
М.Шлейден-ботан, Т.Шванн-зоолог, Р.Вирхов-патан.
1838-39-осн положения:
1) все живые орг-мы сост. из клеток(клл. сходны по стр-ю и осн. св-вам),
2) клетка –единица живого (вне кл. нет жизни);
1858-59-Вирхов
3)Omnis cellula e cellula(клл увелич-ся в ч-ле путемделения исх кл пс удв-я её генетич мат-ла)
/4) Кл.- единая система сопряженных функц. единиц (субструктур~органелл)
5) Клл. многокл орг-мов тотипотентны, т.е.
а) равнозначны по V генетич info, облад всеми возм-стями клл данного орг-ма,
б)отличаются друг от друга разной экспрессией генов(акт-стью) –> дифференцировка.
6) Многокл орг-м -новая сист.- сложн. ансамбль из мн-ва клл, объед. и интегрированный в подсистт. тканей и органов, связ. друг с другом с помощью хим. факторов.
2. Ядерно-цитоплазматический транспорт
-м-лы до 60 кДа(d=9нм)проникают через поры свободно, по gradC(пассивная диффузия), v~m
-более крупные м-лы проходят с помощью акт.транспорта (Еатф, белки-переносчики)
-транспортные белки(шаттлы= «челноки»)
—эксопртины(выводят пре-рибосомы, тРНК, и/рРНП)
—импортины(вводят белки: ламины, для мяРНП, гистоны, кислые белки)
—для импорта необх.:1)NLS(nuclear localisation sequence) последовательность, 2) рецептор(нуклеопорины), 3)АТФ(для транслокации)
—подробнее: [импортин-α+импортин-β+cargo]→ в ядро → +GTP=>[GTP+импортин- β] +импортин-α +cargo; cargo ост.в ядре, ост.возвр.в цитоплазму; в цитоплазме: +GAP=> импортин- β +GDP+P(возвр.в ядро)
—для экспорта необх.:1)NES(nuclear export sequence), 2)GTP, 3)белки
—подробнее: [cargo(NES)+exportin-1+GTP(Ran)]→через поровый комплекс , из ядра→в цитопл. под возд. GAP (GTP Associating Protein) →(GDP(Ran)+P)+exportin-1 +cargo; cargo ост.в цитоплазме, ост.возвр.в ядро; в ядре: GDP(Ran)+RCC-1→GTP(Ran)
-Сущ. белки-транспортеры (РНКнаружу); транспорт, если:
1) правильная(не дефектная) последовательность РНК,
2) завершен сплайсинг(нет интронов),
3) исп-ся белок-транспортер-(гетерогенный)hnRNPA1
—в пор.комплексе меняется оболочка cargo: в ядре CBC(cap binding complex), в цитоплазме PABP (poly A binding protein)
3. Ядерная оболочка, её структура и роль
-ЯО сост.из 2х мембран(внеш и внутр), между ними перинукл. простр-во; внутр. связ. с ламиной, наруж.-с риб. и глЭР; ЯО имеет ядерные поры (отл. от МХ иХЛ)
-ЯО-регулятор ядерно-цитоплазм.транспорта, при этом комплекс яд.поры – транслокатор и сортировщик
—пониж метаболич.акт-сть=>пор меньше(эритроцит)
—наруж.мембрана:интегр., транмп.белки
—внутр.мембрана:транспорт только через поры
—LBR(lamini binding receptor); LAP-1,LAP-2(lamin assoc.protein), эмерин – интегр.белки, связ.внутр.мембр. с ламиной
-ламина имеет решетчатую структ. (замораживание, скалывание, напыление, травление)
-ламина “заякоривает” хроматин на внутр.мембране
РОЛЬ:
1)ЯО характ.для ЭУ, обособляет синтез Р/ДНК от синтеза белка=>регуляция клет.акт-сти; 2)3-мерная структ.интерФ-ного ядра(часть-ЯБМ); 3)регуляция ядерного “импорта” и “экспорта”
-ЯО восстанавливается из поровых комплексов, ламины и везикул
4. Стр-е и ф-ции яд.поры(ЯП)
-компл.ЯП сост.из белков нуклеопоринов
—М у дрожжей –50 МДа=30 белков, у позвоночных 120 МДа=50-100 белков
-во всех моделях ЯП присутствует внутриядерная корзина (h=200нм)
—одна из моделей: 2 ”кольца”(d=120 нм) – цитоплазм. И ядерное(по 8 субъединиц), “спицы”(80нм – d поры), центр.гранула(10-40нм), “корзина”
-Ф-ЦИИ: 1)транслокатор (мех.сито,по gradC),2) сорт-щик (рецепция и сегрегация)
5.Локализация хр-м в интерФ-м ядре
ИнтерФ-рабочая Ф кл-ного ядра или t,когда хр-мы функц-ют. На этой Ф хр-мы б.ч. деконденсируются.
Степень деконденсации ~ активности (min-const метаболически неактивн.уч-ки структ. гетероХ).
Кроме периферич. слоя Х,с яд.оболочкой находятся в контакте специфч. уч-ки хр-м, такие, как половые хр-мы, околоцентромерные уч-ки гетероХ, теломерные хромоцентры, гетероХ,ассоц.с ядрышком и др.(дифф. окраска на гетероХ).=>Ведущая роль этой связи (преимущ. связь гетероХ с яд.оболочкой)
Сущ. модель орг-ции интерФ-ного ядра: развернутая хр-ма в интерФ «заякорена» на ядерной оболочке с помощью гетероХ-вых уч-ков (теломерный гетероХ, прицентормерный гетероХ, околоядрышковый гетероХ, вставочные зоны гетероХ), так,что её расположение становится фикс. в простр-ве ядра, часто повторяя телоФ-ную ориентацию, и занимает в нем соотв. V.
Кроме компактных уч-ков гетероХ сущ. больш. ч-ло конденс. уч-ков эуХ.
6.Полиплоидия, политения, их значение
полиплоидия(П)-увеличение с (кол-ва ДНК).
(эуплоидия-кратно 2n,анэуплоидия-не кратно 2n-чуть меньше/больше-признак рака)
(П)-рез-т нарушения фаз клеточного цикла.
А)полиплоидизирующий М (нет цитокинеза)-развитие человеч. печени
Б)колхицино~М(К-М)-(выпадает телоФ и анаФ)-нет расхожд. в метаФ-кардиомиоциты
В)М выпадает (не пост.)-при облучениях, обр. полиплоидные лимфоциты(эндорепликация-нет М однократно)
Г)политенизация(многонитчатость)-у насекомых (мотыль-личинка хирономуса, человеч. эмбрион-трофобласт -кл. пуповины) — (репл->покой->Терм.)
Д)слияние->дикарион
Е)n ядер -> поликарион (10-20), >20ядер->симпласт (поперечно -полосатая мм.)
Ж)многополюсный М -расхожд. хр-м, обр-е неск. ядер(похоже на Д)
пуф-место транскрипции (синтез РНК),диски-зарепрессированные хр-мные ус-ки
пуф-врем-е обр-е(аналогично «ёлочкам»ядрышек ооцитов рыб)
знач.-увеличение размера->продуктивности,рост органов
7. Ядерный белковый матрикс(ЯБМ)
-белковый компонент, «держащий» структ.ядра
-ДНК имеет участки, взаимод.с ЯБМ специфич.образом
-экстркция ядерных компонентов в процессе выделения ЯБМ:(пс.этих действий ядро не теряет своей целосности)
—0.2 мМ MgCl2-связать белки(чтобы сохранить матрикс),
—2M NaCl-гипотонич р-р(для удаления всех гистонов),
—1% тритон Х100(неионный детергент)-разрушение ядерной оболочки,
—ДНК-аза и РНК-аза-отщепление ДНК и РНК.
—ЛАМИНА(Л)-подстилает внутр.мембрану яд.оболочки
-белки,вход.в состав Л:ламины А,В,С
-Л соединяет яд.оболочку и конденс.Х
-ЯБМ=(Л)+остатки ядрышка (белковый матрикс) +поровые комплексы+ (межхроматиновая белковая сеть матрикса) (Збарский,Ченцов,Георгиев)
—ЯБМ-не артефакт,т.к.связь с ДНК-специфич.и функц.
—Состав ЯБМ:
—белок97,ДНК0.1,РНК1.2,фосфолипиды1.1(крысиная печень)
—белок92.3,ДНК1.2,РНК0.05,фосфолипиды6.9(HeLa)
—ДНК: 10000 нукл.посл.- сателлитные,120-140-гетерогенные
—РНК: гетерогенно-ядерная, рРНК, тРНК, малая ядерная
-транскрипция связана с ЯБМ(располагает участки Д/РНК опр.образом)
-Сущ. артефакт-белковый остов внутри хр-мы(scafull)-он обнаруживается только тотально(т.е.только из белков)
11. Док-ва непр-сти хр-м в течние клеточного цикла
В интерфазном ядре не видно хр-м, подобных митотическим (они там есть).
-
Наблюдения Бовери(1907) за морф. постоянством хр-м при делении бластомеров аскариды→хар-р распол-я хр-м в телоФ опр-т форму интерфазного ядра и порядок распол-я хр-м в след. профазе (это посл.основой для теории↑).
-
(косв)Пост-во ч-ла и морфологии хр-м для данного кариотипа нельзя объяснить при «разборке» хр-м.
-
Закономерно повт-ся расположение хр-м в метафазных пластинках
-
Правило Тейлора(1964)→воспроизв-е хр-м сходно с полуконсервативной редупликацией ДНК (метка Н3Т сохр-ся в одной хр-ме пс. деления).
-
Индивид. хр-мы иногда можно наблюдать в интерФ-х ядрах(тельце Барра).
-
В ряде объектов возм. выявление теломерных уч-ков хр-м в интерФ-х ядрах(хромоцентры итерФ-х ядер клеток меристемы корешков лука-теломерные уч-ки хр-м—радиоавтограф)
-
Центромерные уч-ки хр-м в интерФ-х ядрах выявл-ся диффер. окраской на гетерохроматин(культура фибробластов мыши).
-
Зоны локализации отд. хр-м можно выявить и в интерФ-х ядрах, не имеющих хромоцентров или конденсрованных уч-ков хр-м.(импульсная Н3Т метка в среде пс. делений располагается не диффузно, а над опр уч-ками)
-
Изучение репаративного синтеза ДНК при УФ-облучении клеток→не > ½ хр-м клетки облуч-ся перед делением, облученная клетка поглощает Н3Т до синт. периода , т.к. репартивный синтез, причем ввключ-я неравномерно, в соотв с пораж. хр-мами.
10. Прямые биохим данные: при опр-и мах М ДНК дрозофиллы→1хр-ма–1ДНК, длина-const в митозе и интерФ
14. Клеточный цикл, его стадии и способы изучения
-время сущ-я кл.как таковойот деления до деления = клет.цикл (бактерия-20мин, стентор-2-3сут.)
—смысл кл-ного деления — в равномерном распр-и редуплиц. кл-ного мат-ла по 2м новым клл.
→G1→S→G2постсинтетич/премитотич{→G0 Ф пролиферативного покоя, откр. Lagta-R2}→ M{→G0-R(est)1} →G1пресинтетич/постмитотич→
—продолжительность ФФ сильно варьирует у разл.клл, но ~ одинакова у клл. 1 органа
-разл.содерж-е Д/РНК и интенс-сть их синтеза (по ФФ):
G1 — 2c(ДНК), S↑(РНК),S(1/4→1)max(белок)
S — (2→4)c, S(ДНК), Smax(РНК),Smax(белок)
G2 — 4c, Smax(РНК), S(1/4→1)max(белок)
M — (4→2)c, 1/4Smax(белок)
-G1-рост кл., подготовкак синтезу ДНК(синтез белка-инициатора S?), синтезы ферментов метаболизма РНК и белка, ферм.,необх для обр-я ДНК
—S –етсь всегда(искл.-2е деление мейоза), длительность ~v репл.ДНК(ч-лу репликонов), v(полипл.)=v(дипл.); синтез гистонов в цитопл., синтез рРНК(необх в G2)
—G2 –обычно короче ост.периодов, м. Выпадать; синтез кл-ных РНК и белков, синтез иРНК (для М), рРНК из S исп-ся для синтеза «белков деления»(напр.тубулинов для М веретена)
—G0 –дифф.клл, либо “в покое”(могут делиться)
—способы изуч-я:
1)метод получения гетерокарионов (с помощью инактивированных вирусов) из 2х разл клл.=> индуцированное влияние со ст-ны цитоплазм факторов
2) включение Н3-предшественников (синтезов Д/РНК)
{М-деление, ост. -интерФ}
15. Мейоз(МЗ), последовательнось фаз мейоза и его значение
—МЗ-2 клет.цикла, клл.делятся, но репл-ся только 1 раз
-процесс МЗ-а универсален для всех ЭУ орг-мов
-одна из специализаций- пол.клл., команда – оч.рано (циклоп-пс.4го деления, дрозофилла – ранняя бластула, ч-к – до заклкдки всех органов — в каудальном отделе)
—Август Вейсман: (для пол клл.) 1е делелние МЗ 2n→{S}→4n→{ProI(длинная профаза)}→{MI}→2*2n→ {нетS}→2*2n→{MII}→4*n (единица репл.–хр-ма)
—первичн.зарод.клл. →гонии→ауксоциты→(синапсис) →мейотич.деление→гаметы(рост ооцита, дифф. сператоцита)
—обр-е зиготы: ♀(1n)+♂(1n) →2n→(S)→4n→(M) →2*2n
-ProI сост.из неск.уч-ков:
1) лептотена(тонк.нить); «хр-мный букет»
2) зиготена(соед.нить); объед.матер. и отцовск.(1-1)
3) пахитена(толст.нить); длинная у ч-ка и мыши, обр-ся синаптинемный комплекс(характ для МЗ, 0.6 Σt)
4) диплотена(двойная нить); центромерные уч-ки отталкиваются , связь — хиазмы
5) диплокинез(расхождение хр-м);
—МЗ(отличия от М):
1) ProI – длительная (сом.0.5-1.5ч, пол.-до 50 лет)
Строение клетки
Строение прокариотических и эукариотических клеток
Основными структурными компонентами клеток являются плазматическая мембрана, цитоплазма и наследственный аппарат. В зависимости от особенностей организации различают два основных типа клеток: прокариотические и эукариотические. Главным отличием прокариотических клеток от эукариотических является организация их наследственного аппарата: у прокариот он находится непосредственно в цитоплазме (эта область цитоплазмы называется нуклеоидом) и не отделен от нее мембранными структурами, тогда как у эукариот бульшая часть ДНК сосредоточена в ядре, окруженном двойной мембраной. Кроме того, генетическая информация прокариотических клеток, находящаяся в нуклеоиде, записана в кольцевой молекуле ДНК, а у эукариот молекулы ДНК незамкнутые.
В отличие от эукариот, цитоплазма прокариотических клеток содержит также небольшое количество органоидов, тогда как для эукариотических характерно значительное разнообразие этих структур.
Строение и функции биологических мембран
Строение биомембраны. Мембраны, ограничивающие клетки и мембранные органоиды эукариотических клеток, имеют общий химический состав и строение. В их состав входят липиды, белки и углеводы. Липиды мембраны представлены в основном фосфолипидами и холестерином. Большинство белков мембран относится к сложным белкам, например гликопротеинам. Углеводы не встречаются в мембране самостоятельно, они связаны с белками и липидами. Толщина мембран составляет 7–10 нм.
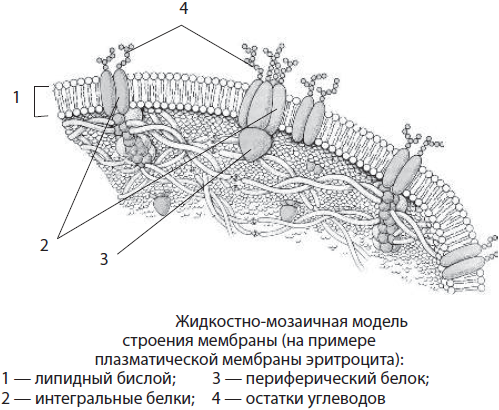
Согласно общепринятой в настоящее время жидкостно-мозаичной модели строения мембран, липиды образуют двойной слой, или липидный бислой, в котором гидрофильные «головки» молекул липидов обращены наружу, а гидрофобные «хвосты» спрятаны вовнутрь мембраны. Эти «хвосты» благодаря своей гидрофобности обеспечивают разделение водных фаз внутренней среды клетки и ее окружения. С липидами с помощью различных типов взаимодействия связаны белки. Часть белков расположена на поверхности мембраны. Такие белки называют периферическими, или поверхностными. Другие белки частично или полностью погружены в мембрану — это интегральные, или погруженные белки. Белки мембран выполняют структурную, транспортную, каталитическую, рецепторную и другие функции.
Мембраны не похожи на кристаллы, их компоненты постоянно находятся в движении, вследствие чего между молекулами липидов возникают разрывы — поры, через которые в клетку могут попадать или покидать ее различные вещества.
Биологические мембраны различаются по расположению в клетке, химическому составу и выполняемым функциям. Основные типы мембран — плазматическая и внутренние. Плазматическая мембрана содержит около 45 % липидов (в т. ч. гликолипидов), 50 % белков и 5 % углеводов. Цепочки углеводов, входящих в состав сложных белков-гликопротеинов и сложных липидов-гликолипидов, выступают над поверхностью мембраны. Гликопротеины плазмалеммы чрезвычайно специфичны. Так, например, по ним происходит взаимное узнавание клеток, в том числе сперматозоида и яйцеклетки.
На поверхности животных клеток углеводные цепочки образуют тонкий поверхностный слой — гликокаликс. Он выявлен почти во всех животных клетках, но степень его выраженности неодинакова (10–50 мкм). Гликокаликс обеспечивает непосредственную связь клетки с внешней средой, в нем происходит внеклеточное пищеварение; в гликокаликсе размещены рецепторы. Клетки бактерий, растений и грибов, помимо плазмалеммы, окружены еще и клеточными оболочками.
Внутренние мембраны эукариотических клеток разграничивают различные части клетки, образуя своеобразные «отсеки» — компартменты, что способствует разделению различных процессов обмена веществ и энергии. Они могут различаться по химическому составу и выполняемым функциям, но общий план строения у них сохраняется.
Функции мембран:
- Ограничивающая. Заключается в том, что они отделяют внутреннее пространство клетки от внешней среды. Мембрана является полупроницаемой, то есть ее свободно преодолевают только те вещества, которые необходимы клетке, при этом существуют механизмы транспорта необходимых веществ.
- Рецепторная. Связана в первую очередь с восприятием сигналов окружающей среды и передачей этой информации внутрь клетки. За эту функцию отвечают специальные белки-рецепторы. Мембранные белки отвечают еще и за клеточное узнавание по принципу «свой-чужой», а также за образование межклеточных соединений, наиболее изученными из которых являются синапсы нервных клеток.
- Каталитическая. На мембранах расположены многочисленные ферментные комплексы, вследствие чего на них происходят интенсивные синтетические процессы.
- Энерготрансформирующая. Связана с образованием энергии, ее запасанием в виде АТФ и расходованием.
- Компартментализация. Мембраны разграничивают также пространство внутри клетки, разделяя тем самым исходные вещества реакции и ферменты, которые могут осуществлять соответствующие реакции.
- Образование межклеточных контактов. Несмотря на то, что толщина мембраны настолько мала, что ее невозможно различить невооруженным глазом, она, с одной стороны, служит достаточно надежным барьером для ионов и молекул, в особенности водорастворимых, а с другой — обеспечивает их перенос в клетку и наружу.
- Транспортная.
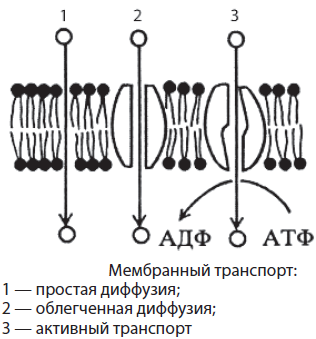
Мембранный транспорт. В связи с тем, что клетки как элементарные биологические системы являются открытыми системами, для обеспечения обмена веществ и энергии, поддержания гомеостаза, роста, раздражимости и других процессов требуется перенос веществ через мембрану — мембранный транспорт. В настоящее время транспорт веществ через мембрану клетки делят на активный, пассивный, эндо- и экзоцитоз.
Пассивный транспорт — это вид транспорта, который происходит без затраты энергии от большей концентрации к меньшей. Растворимые в липидах небольшие неполярные молекулы (О2, СО2) легко проникают в клетку путем простой диффузии. Нерастворимые же в липидах, в том числе заряженные небольшие частицы, подхватываются белкамипереносчиками или проходят через специальные каналы (глюкоза, аминокислоты, К+, PO43-). Такой вид пассивного транспорта называется облегченной диффузией. Вода поступает в клетку через поры в липидной фазе, а также по специальным каналам, выстланным белками. Транспорт воды через мембрану называется осмосом.
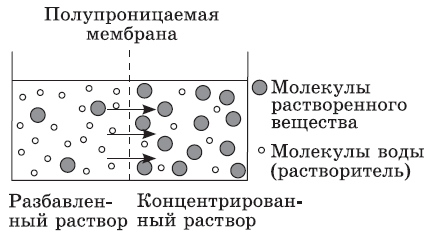
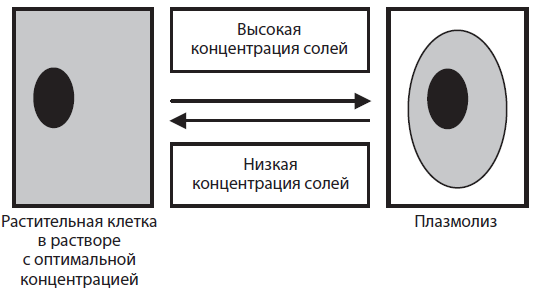
Осмос имеет чрезвычайно важное значение в жизни клетки, так как если ее поместить в раствор с более высокой концентрацией солей, чем в клеточном растворе, то вода начнет выходить из клетки, и объем живого содержимого начнет уменьшаться. У животных клеток происходит съеживание клетки в целом, а у растительных — отставание цитоплазмы от клеточной стенки, которое называется плазмолизом. При помещении клетки в менее концентрированный, чем цитоплазма, раствор, транспорт воды происходит в обратном направлении — в клетку. Однако существуют пределы растяжимости цитоплазматической мембраны, и животная клетка в конце концов разрывается, а у растительной этого не позволяет сделать прочная клеточная стенка. Явление заполнения клеточным содержимым всего внутреннего пространства клетки называется деплазмолизом. Внутриклеточную концентрацию солей следует учитывать при приготовлении лекарственных препаратов, особенно для внутривенного введения, так как это может приводить к повреждению клеток крови (для этого используют физиологический раствор с концентрацией 0,9 % хлорида натрия). Это не менее важно при культивировании клеток и тканей, а также органов животных и растений.
Активный транспорт протекает с затратой энергии АТФ от меньшей концентрации вещества к большей. Он осуществляется с помощью специальных белков-насосов. Белки перекачивают через мембрану ионы К+, Na+, Са2+ и другие, что способствует транспорту важнейших органических веществ, а также возникновению нервных импульсов и т. д.
Эндоцитоз — это активный процесс поглощения веществ клеткой, при котором мембрана образует впячивания, а затем формирует мембранные пузырьки — фагосомы, в которых заключены поглощаемые объекты. Затем с фагосомой сливается первичная лизосома, и образуется вторичная лизосома, или фаголизосома, или пищеварительная вакуоль. Содержимое пузырька расщепляется ферментами лизосом, а продукты расщепления поглощаются и усваиваются клеткой. Непереваренные остатки удаляются из клетки путем экзоцитоза. Различают два основных вида эндоцитоза: фагоцитоз и пиноцитоз.
Фагоцитоз — это процесс захвата клеточной поверхностью и поглощения клеткой твердых частиц, а пиноцитоз — жидкости. Фагоцитоз протекает в основном в животных клетках (одноклеточные животные, лейкоциты человека), он обеспечивает их питание, а часто и защиту организма . Путем пиноцитоза происходит поглощение белков, комплексов антиген-антитела в процессе иммунных реакций и т. д. Однако путем пиноцитоза или фагоцитоза в клетку также попадают многие вирусы. В клетках растений и грибов фагоцитоз практически невозможен, так как они окружены прочными клеточными оболочками.
Экзоцитоз — процесс, обратный эндоцитозу. Таким образом выделяются непереваренные остатки пищи из пищеварительных вакуолей, выводятся необходимые для жизнедеятельности клетки и организма в целом вещества. Например, передача нервных импульсов происходит благодаря выделению посылающим импульс нейроном химических посредников — медиаторов, а в растительных клетках так выделяются вспомогательные углеводы клеточной оболочки.
Клеточные оболочки клеток растений, грибов и бактерий. Снаружи от мембраны клетка может выделять прочный каркас — клеточную оболочку, или клеточную стенку.
У растений основу клеточной оболочки составляет целлюлоза, упакованная в пучки по 50–100 молекул. Промежутки между ними заполняют вода и другие углеводы. Оболочка растительной клетки пронизана канальцами — плазмодесмами, через которые проходят мембраны эндоплазматической сети. По плазмодесмам осуществляется транспорт веществ между клетками. Однако транспорт веществ, например воды, может происходить и по самим клеточным стенкам. Со временем в клеточной оболочке растений накапливаются различные вещества, в том числе дубильные или жироподобные, что приводит к одревеснению или опробковению самой клеточной стенки, вытеснению воды и отмиранию клеточного содержимого. Между клеточными стенками соседних клеток растений располагаются желеобразные прокладки — срединные пластинки, которые скрепляют их между собой и цементируют тело растения в целом. Они разрушаются только в процессе созревания плодов и при опадании листьев.
Клеточные стенки клеток грибов образованы хитином — углеводом, содержащим азот. Они достаточно прочны и являются внешним скелетом клетки, но все же, как и у растений, препятствуют фагоцитозу.
У бактерий в состав клеточной стенки входит углевод с фрагментами пептидов — муреин, однако его содержание существенно различается у разных групп бактерий. Поверх от клеточной стенки могут выделяться также иные полисахариды, образующие слизистую капсулу, защищающую бактерии от внешних воздействий.
Оболочка определяет форму клетки, служит механической опорой, выполняет защитную функцию, обеспечивает осмотические свойства клетки, ограничивая растяжение живого содержимого и предотвращая разрыв клетки, увеличивающейся вследствие поступления воды. Кроме того, клеточную стенку преодолевают вода и растворенные в ней вещества, прежде чем попасть в цитоплазму или, наоборот, при выходе из нее, при этом по клеточным стенкам вода транспортируется быстрее, чем по цитоплазме.
Цитоплазма
Цитоплазма — это внутреннее содержимое клетки. В нее погружены все органоиды клетки, ядро и разнообразные продукты жизнедеятельности.
Цитоплазма связывает все части клетки между собой, в ней протекают многочисленные реакции обмена веществ. Цитоплазма отделяется от окружающей среды и делится на отсеки мембранами, то есть клеткам присуще мембранное строение. Она может находиться в двух состояниях — золя и геля. Золь — это полужидкое, киселеобразное состояние цитоплазмы, при котором процессы жизнедеятельности протекают наиболее интенсивно, а гель — более плотное, студнеобразное состояние, затрудняющее протекание химических реакций и транспорт веществ.
Жидкая часть цитоплазмы без органоидов называется гиалоплазмой. Гиалоплазма, или цитозоль, представляет собой коллоидный раствор, в котором находится своеобразная взвесь достаточно крупных частиц, например белков, окруженных диполями молекул воды. Осаждения этой взвеси не происходит вследствие того, что они имеют одинаковый заряд и отталкиваются друг от друга.
Органоиды
Органоиды — это постоянные компоненты клетки, выполняющие определенные функции.
В зависимости от особенностей строения их делят на мембранные и немембранные. Мембранные органоиды, в свою очередь, относят к одномембранным (эндоплазматическая сеть, комплекс Гольджи и лизосомы) или двумембранным (митохондрии, пластиды и ядро). Немембранными органоидами являются рибосомы, микротрубочки, микрофиламенты и клеточный центр. Прокариотам из перечисленных органоидов присущи только рибосомы.
Строение и функции ядра. Ядро — крупный двумембранный органоид, лежащий в центре клетки или на ее периферии. Размеры ядра могут колебаться в пределах 3–35 мкм. Форма ядра чаще сферическая или эллипсоидная, однако имеются также палочковидные, веретеновидные, бобовидные, лопастные и даже сегментированные ядра. Некоторые исследователи считают, что форма ядра соответствует форме самой клетки.
Большинство клеток имеет одно ядро, но, например, в клетках печени и сердца их может быть два, а в ряде нейронов — до 15. Волокна скелетных мышц содержат обычно много ядер, однако они не являются клетками в полном смысле этого слова, поскольку образуются в результате слияния нескольких клеток.
Ядро окружено ядерной оболочкой, а его внутреннее пространство заполнено ядерным соком, или нуклеоплазмой (кариоплазмой), в которую погружены хроматин и ядрышко. Ядро выполняет такие важнейшие функции, как хранение и передача наследственной информации, а также контроль жизнедеятельности клетки.
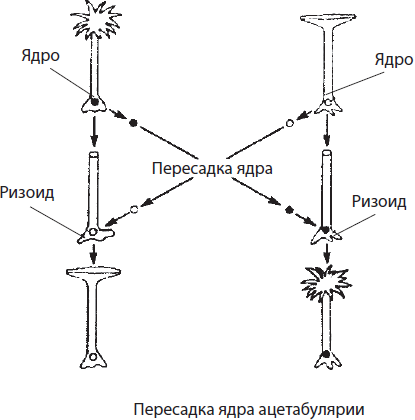
Роль ядра в передаче наследственной информации была убедительно доказана в экспериментах с зеленой водорослью ацетабулярией. В единственной гигантской клетке, достигающей в длину 5 см, различают шляпку, ножку и ризоид. При этом она содержит только одно ядро, расположенное в ризоиде. В 1930-е годы И. Хеммерлинг пересадил ядро одного вида ацетабулярии с зеленой окраской в ризоид другого вида, с коричневой окраской, у которого ядро было удалено. Через некоторое время у растения с пересаженным ядром выросла новая шляпка, как у водоросли- донора ядра. В то же время отделенные от ризоида шляпка или ножка, не содержащие ядра, через некоторое время погибали.
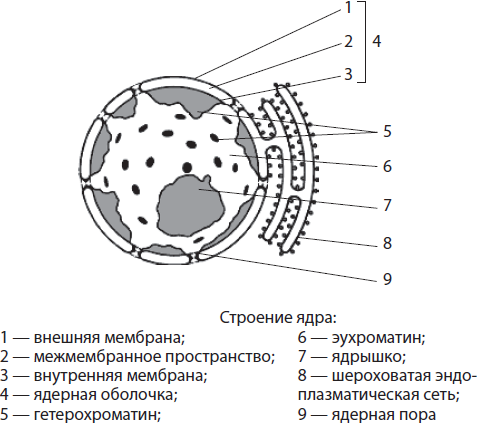
Ядерная оболочка образована двумя мембранами — наружной и внутренней, между которыми есть пространство. Межмембранное пространство сообщается с полостью шероховатой эндоплазматической сети, а наружная мембрана ядра может нести рибосомы. Ядерная оболочка пронизана многочисленными порами, окантованными специальными белками. Через поры происходит транспорт веществ: в ядро попадают необходимые белки (в т. ч. ферменты), ионы, нуклеотиды и другие вещества, и покидают его молекулы РНК, отработанные белки, субъ единицы рибосом. Таким образом, функциями ядерной оболочки являются отделение содержимого ядра от цитоплазмы, а также регуляция обмена веществ между ядром и цитоплазмой.
Нуклеоплазмой называют содержимое ядра, в которое погружены хроматин и ядрышко. Она представляет собой коллоидный раствор, по химическому составу напоминающий цитоплазму. Ферменты нуклеоплазмы катализируют обмен аминокислот, нуклеотидов, белков и др. Нуклеоплазма связана с гиалоплазмой через ядерные поры. Функции нуклеоплазмы, как и гиалоплазмы, состоят в обеспечении взаимосвязи всех структурных компонентов ядра и осуществлении ряда ферментных реакций.
Хроматином называют совокупность тонких нитей и гранул, погруженных в нуклеоплазму. Выявить его можно только при окрашивании, так как коэффициенты преломления хроматина и нуклеоплазмы приблизительно одинаковы. Нитчатый компонент хроматина называют эухроматином, а гранулярный — гетерохроматином. Эухроматин слабо уплотнен, поскольку с него считывается наследственная информация, тогда как более спирализованный гетерохроматин является генетически неактивным.
Хроматин представляет собой структурное видоизменение хромосом в неделящемся ядре. Таким образом, хромосомы постоянно присутствуют в ядре, изменяется лишь их состояние в зависимости от функции, которую ядро выполняет в данный момент.
В состав хроматина в основном входят белки-нуклеопротеины (дезоксирибонуклеопротеины и рибонуклеопротеины), а также ферменты, важнейшие из которых связаны с синтезом нуклеиновых кислот, и некоторые другие вещества.
Функции хроматина состоят, во-первых, в синтезе специфических для данного организма нуклеиновых кислот, которые направляют синтез специфических белков, во-вторых, в передаче наследственных свойств от материнской клетки дочерним, для чего хроматиновые нити в процессе деления упаковываются в хромосомы.
Ядрышко — сферическое, хорошо заметное под микроскопом тельце диаметром 1–3 мкм. Оно формируется на участках хроматина, в которых закодирована информация о структуре рРНК и белках рибосом. Ядрышко в ядре часто одно, однако в тех клетках, где происходят интенсивные процессы жизнедеятельности, ядрышек может быть два и более. Функции ядрышек — синтез рРНК и сборка субъединиц рибосом путем объединения рРНК с белками, поступающими из цитоплазмы.
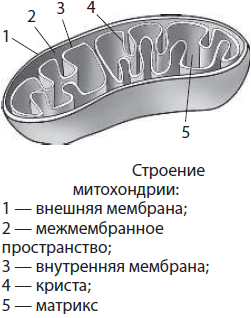
Митохондрии — двумембранные органоиды округлой, овальной или палочковидной формы, хотя встречаются и спиралевидные (в сперматозоидах). Диаметр митохондрий составляет до 1 мкм, а длина — до 7 мкм. Пространство внутри митохондрий заполнено матриксом. Матрикс — это основное вещество митохондрий. В него погружены кольцевая молекула ДНК и рибосомы. Наружная мембрана митохондрий гладкая, она непроницаема для многих веществ. Внутренняя мембрана имеет выросты — кристы, увеличивающие площадь поверхности мембран для протекания химических реакций. На поверхности мембраны расположены многочисленные белковые комплексы, составляющие так называемую дыхательную цепь, а также грибовидные ферменты АТФ-синтетазы. В митохондриях протекает аэробный этап дыхания, в ходе которого происходит синтез АТФ.
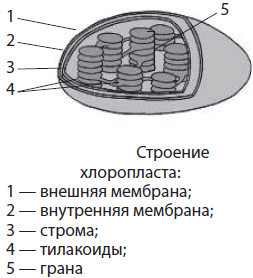
Пластиды — крупные двумембранные органоиды, характерные только для растительных клеток. Внутреннее пространство пластид заполнено стромой, или матриксом. В строме находится более или менее развитая система мембранных пузырьков — тилакоидов, которые собраны в стопки — граны, а также собственная кольцевая молекула ДНК и рибосомы. Различают четыре основных типа пластид: хлоропласты, хромопласты, лейкопласты и пропластиды.
Хлоропласты — это зеленые пластиды диаметром 3–10 мкм, хорошо различимые под микроскопом. Они содержатся только в зеленых частях растений — листьях, молодых стеблях, цветках и плодах. Хлоропласты в основном имеют овальную или эллипсоидную формы, но могут быть также чашевидными, спиралевидными и даже лопастными. Количество хлоропластов в клетке в среднем составляет от 10 до 100 штук. Однако, например, у некоторых водорослей он может быть один, иметь значительные размеры и сложную форму — тогда его называют хроматофором. В других случаях количество хлоропластов может достигать нескольких сотен, при этом их размеры невелики. Окраска хлоропластов обусловлена основным пигментом фотосинтеза — хлорофиллом, хотя в них содержатся и дополнительные пигменты — каротиноиды. Каротиноиды становятся заметными только осенью, когда хлорофилл в стареющих листьях разрушается. Основной функцией хлоропластов является фотосинтез. Световые реакции фотосинтеза протекают на мембранах тилакоидов, на которых закреплены молекулы хлорофилла, а темновые реакции — в строме, где содержатся многочисленные ферменты.
Хромопласты — это желтые, оранжевые и красные пластиды, содержащие пигменты каротиноиды. Форма хромопластов может также существенно варьировать: они бывают трубчатыми, сферическими, кристаллическими и др. Хромопласты придают окраску цветкам и плодам растений, привлекая опылителей и распространителей семян и плодов.
Лейкопласты — это белые или бесцветные пластиды в основном округлой или овальной формы. Они распространены в нефотосинтезирующих частях растений, например в кожице листа, клубнях картофеля и т. д. В них откладываются в запас питательные вещества, чаще всего крахмал, но у некоторых растений это могут быть белки или масло.
Пластиды образуются в растительных клетках из пропластид, которые имеются уже в клетках образовательной ткани и представляют собой небольшие двумембранные тельца. На ранних этапах развития разные виды пластид способны превращаться друг в друга: при попадании на свет лейкопласты клубня картофеля и хромопласты корнеплода моркови зеленеют.
Пластиды и митохондрии называют полуавтономными органоидами клетки, так как они имеют собственные молекулы ДНК и рибосомы, осуществляют синтез белка и делятся независимо от деления клеток. Эти особенности объясняются происхождением от одноклеточных прокариотических организмов. Однако «самостоятельность » митохондрий и пластид является ограниченной, так как их ДНК содержит слишком мало генов для свободного существования, остальная же информация закодирована в хромосомах ядра, что позволяет ему контролировать данные органоиды.
Эндоплазматическая сеть (ЭПС), или эндоплазматический ретикулум (ЭР), — это одномембранный органоид, представляющий собой сеть мембранных полостей и канальцев, занимающих до 30 % содержимого цитоплазмы. Диаметр канальцев ЭПС составляет около 25–30 нм. Различают два вида ЭПС — шероховатую и гладкую. Шероховатая ЭПС несет рибосомы, на ней происходит синтез белков. Гладкая ЭПС лишена рибосом. Ее функция — синтез липидов и углеводов, а также транспорт, запасание и обезвреживание токсических веществ. Она особенно развита в тех клетках, где происходят интенсивные процессы обмена веществ, например в клетках печени — гепатоцитах — и волокнах скелетных мышц. Вещества, синтезированные в ЭПС, транспортируются в аппарат Гольджи. В ЭПС происходит также сборка мембран клетки, однако их формирование завершается в аппарате Гольджи.
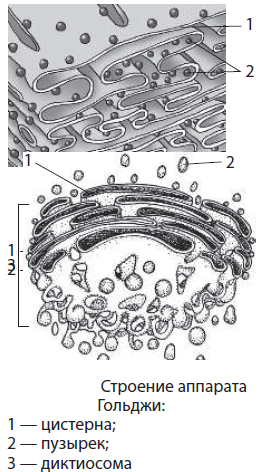
Аппарат Гольджи, или комплекс Гольджи, — одномембранный органоид, образованный системой плоских цистерн, канальцев и отшнуровывающихся от них пузырьков. Структурной единицей аппарата Гольджи является диктиосома — стопка цистерн, на один полюс которой приходят вещества из ЭПС, а с противоположного полюса, подвергшись определенным превращениям, они упаковываются в пузырьки и направляются в другие части клетки. Диаметр цистерн — порядка 2 мкм, а мелких пузырьков — около 20–30 мкм. Основные функции комплекса Гольджи — синтез некоторых веществ и модификация (изменение) белков, липидов и углеводов, поступающих из ЭПС, окончательное формирование мембран, а также транспорт веществ по клетке, обновление ее структур и образование лизосом. Свое название аппарат Гольджи получил в честь итальянского ученого Камилло Гольджи, впервые обнаружившего данный органоид (1898).
Лизосомы — небольшие одномембранные органоиды до 1 мкм в диаметре, в которых содержатся гидролитические ферменты, участвующие во внутриклеточном пищеварении. Мембраны лизосом слабопроницаемы для этих ферментов, поэтому выполнение лизосомами своих функций происходит очень точно и адресно. Так, они принимают активное участие в процессе фагоцитоза, образуя пищеварительные вакуоли, а в случае голодания или повреждения определенных частей клетки переваривают их, не затрагивая иных. Недавно была открыта роль лизосом в процессах клеточной гибели.
Вакуоль — это полость в цитоплазме растительных и животных клеток, ограниченная мембраной и заполненная жидкостью. В клетках простейших обнаруживаются пищеварительные и сократительные вакуоли. Первые принимают участие в процессе фагоцитоза, так как в них происходит расщепление питательных веществ. Вторые обеспечивают поддержание водно-солевого баланса за счет осморегуляции. У многоклеточных животных в основном встречаются пищеварительные вакуоли.
В растительных клетках вакуоли присутствуют всегда, они окружены специальной мембраной и заполнены клеточным соком. Мембрана, окружающая вакуоль, по химическому составу, строению и выполняемым функциям близка к плазматической мембране. Клеточный сок представляет собой водный раствор различных неорганических и органических веществ, в том числе минеральных солей, органических кислот, углеводов, белков, гликозидов, алкалоидов и др. Вакуоль может занимать до 90 % объема клетки и оттеснять ядро на периферию. Эта часть клетки выполняет запасающую, выделительную, осмотическую, защитную, лизосомную и другие функции, поскольку в ней накапливаются питательные вещества и отходы жизнедеятельности, она обеспечивает поступление воды и поддержание формы и объема клетки, а также содержит ферменты расщепления многих компонентов клетки. К тому же биологически активные вещества вакуолей способны препятствовать поеданию этих растений многими животными. У ряда растений за счет разбухания вакуолей происходит рост клетки растяжением.
Вакуоли имеются также и в клетках некоторых грибов и бактерий, однако у грибов они выполняют только функцию осморегуляции, а у цианобактерий поддерживают плавучесть и участвуют в процессах усвоения азота из воздуха.
Рибосомы — небольшие немембранные органоиды диаметром 15–20 мкм, состоящие из двух субъединиц — большой и малой. Субъединицы рибосом эукариот собираются в ядрышке, а затем транспортируются в цитоплазму. Рибосомы прокариот, митохондрий и пластид меньше по величине, чем рибосомы эукариот. В состав субъединиц рибосом входят рРНК и белки.
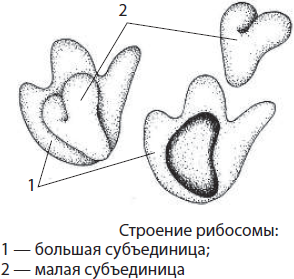
Количество рибосом в клетке может достигать нескольких десятков миллионов: в цитоплазме, митохондриях и пластидах они находятся в свободном состоянии, а на шероховатой ЭПС — в связанном. Они принимают участие в синтезе белка, в частности, осуществляют процесс трансляции — биосинтеза полипептидной цепи на молекуле иРНК. На свободных рибосомах синтезируются белки гиалоплазмы, митохондрий, пластид и собственные белки рибосом, тогда как на прикрепленных к шероховатой ЭПС рибосомах осуществляется трансляция белков для выведения из клеток, сборки мембран, образования лизосом и вакуолей.
Рибосомы могут находиться в гиалоплазме поодиночке или собираться в группы при одновременном синтезе на одной иРНК сразу нескольких полипептидных цепей. Такие группы рибосом называются полирибосомами, или полисомами.
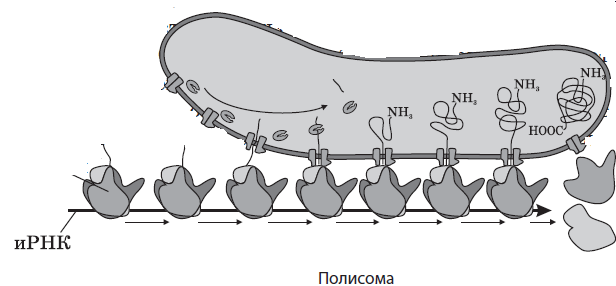
Микротрубочки — это цилиндрические полые немембранные органоиды, которые пронизывают всю цитоплазму клетки. Их диаметр составляет около 25 нм, толщина стенки — 6–8 нм. Они образованы многочисленными молекулами белка тубулина, которые сначала формируют 13 нитей, напоминающих бусы, а затем собираются в микротрубочку. Микротрубочки образуют цитоплазматическую сеть, которая придает клетке форму и объем, связывают плазматическую мембрану с другими частями клетки, обеспечивают транспорт веществ по клетке, принимают участие в движении клетки и внутриклеточных компонентов, а также в делении генетического материала. Они входят в состав клеточного центра и органоидов движения — жгутиков и ресничек.
Микрофиламенты, или микронити, также являются немембранными органоидами, однако они имеют нитевидную форму и образованы не тубулином, а актином. Они принимают участие в процессах мембранного транспорта, межклеточном узнавании, делении цитоплазмы клетки и в ее движении. В мышечных клетках взаимодействие актиновых микрофиламентов с миозиновыми нитями обеспечивает сокращение.
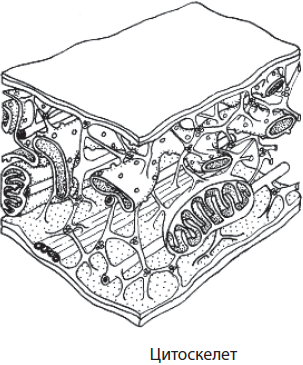
Микротрубочки и микрофиламенты образуют внутренний скелет клетки — цитоскелет. Он представляет собой сложную сеть волокон, обеспечивающих механическую опору для плазматической мембраны, определяет форму клетки, расположение клеточных органоидов и их перемещение в процессе деления клетки.
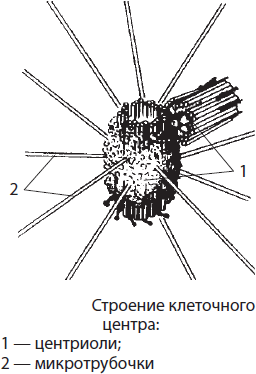
Клеточный центр — немембранный органоид, располагающийся в животных клетках вблизи ядра; в растительных клетках он отсутствует. Его длина составляет около 0.2–0.3 мкм, а диаметр — 0.1–0.15 мкм. Клеточный центр образован двумя центриолями, лежащими во взаимно перпендикулярных плоскостях, и лучистой сферой из микротрубочек. Каждая центриоль образована девятью группами микротрубочек, собранных по три, т. е. триплетами. Клеточный центр принимает участие в процессах сборки микротрубочек, делении наследственного материала клетки, а также в образовании жгутиков и ресничек.
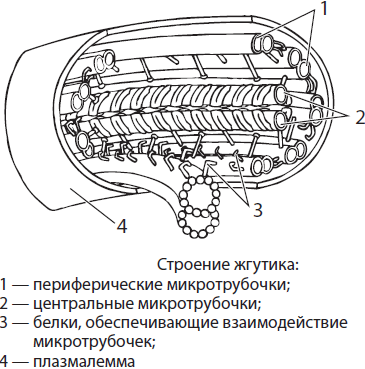
Органоиды движения. Жгутики и реснички представляют собой выросты клетки, покрытые плазмалеммой. Основу этих органоидов составляют девять пар микротрубочек, расположенных по периферии, и две свободные микротрубочки в центре. Микротрубочки связаны между собой различными белками, обеспечивающими их согласованное отклонение от оси — колебание. Колебания энергозависимы, то есть на этот процесс тратится энергия макроэргических связей АТФ. Восстановление утраченных жгутиков и ресничек является функцией базальных телец, или кинетосом, расположенных в их основании.
Длина ресничек составляет около 10–15 нм, а жгутиков — 20–50 мкм. За счет строго направленных движений жгутиков и ресничек осуществляется не только движение одноклеточных животных, сперматозоидов и др., но и происходит очистка дыхательных путей, продвижение яйцеклетки по маточным трубам, поскольку все эти части организма человека выстланы реснитчатым эпителием.
Включения
Включения — это непостоянные компоненты клетки, которые образуются и исчезают в процессе ее жизнедеятельности. К ним относят как запасные вещества, например, зерна крахмала или белка в растительных клетках, гранулы гликогена в клетках животных и грибов, волютина у бактерий, капли жира во всех типах клеток, так и отходы жизнедеятельности, в частности, непереваренные в результате фагоцитоза остатки пищи, образующие так называемые остаточные тельца.
Взаимосвязь строения и функций частей и органоидов клетки — основа ее целостности
Каждая из частей клетки, с одной стороны, является обособленной структурой со специфическим строением и функциями, а с другой — компонентом более сложной системы, называемой клеткой. Бульшая часть наследственной информации эукариотической клетки сосредоточена в ядре, однако само ядро не в состоянии обеспечить ее реализацию, поскольку для этого необходимы как минимум цитоплазма, выступающая как основное вещество, и рибосомы, на которых и происходит этот синтез. Большинство рибосом расположено на гранулярной эндоплазматической сети, откуда белки чаще всего транспортируются в комплекс Гольджи, а затем после модификации — в те части клетки, для которых они предназначены, или выводятся наружу. Мембранные упаковки белков и углеводов могут встраиваться в мембраны органоидов и цитоплазматическую мембрану, обеспечивая их постоянное обновление. От комплекса Гольджи отшнуровываются также выполняющие важнейшие функции лизосомы и вакуоли. Например, без лизосом клетки быстро превратились бы в свое образную свалку отработанных молекул и структур.
Протекание всех этих процессов требует энергии, вырабатываемой митохондриями, а у растений — и хлоропластами. И хотя эти органоиды являются относительно автономными, т. к. имеют собственные молекулы ДНК, часть их белков все равно кодируется ядерным геномом и синтезируется в цитоплазме.
Таким образом, клетка представляет собой неразрывное единство составляющих ее компонентов, каждый из которых выполняет свою уникальную функцию.
Задача С5 ЕГЭ по Биологии. Подборка заданий по цитологии
-
Примеры задач первого типа
-
Примеры задач второго типа
-
Примеры задач третьего типа
-
Примеры задач четвертого типа
-
Примеры задач пятого типа
-
Примеры задач шестого типа
-
Примеры задач седьмого типа
-
Приложение I Генетический код (и-РНК)
-
Ответы
Д. А. Соловков, кандидат биологических наук
Эта подборка задач содержит все основные типы заданий по цитологии, встречающиеся в ЕГЭ, и предназначена, прежде всего, для самостоятельной подготовки абитуриента к решению задания С5 на экзамене. Для удобства задачи сгруппированы по основным разделам и темам, включенным в программу по биологии (раздел «Цитология»). В конце приведены ответы для самопроверки.
Примеры задач первого типа
- В молекуле ДНК содержится
 тимина. Определите, сколько (в ) в этой молекуле содержится других нуклеотидов.
тимина. Определите, сколько (в ) в этой молекуле содержится других нуклеотидов. - В молекуле ДНК содержится тимина. Определите, сколько (в ) в этой молекуле содержится других нуклеотидов.
- В молекуле ДНК содержится гуанина. Определите, сколько (в ) в этой молекуле содержится других нуклеотидов.
- В молекуле ДНК содержится гуанина. Определите, сколько (в ) в этой молекуле содержится других нуклеотидов.
- В молекуле ДНК содержится цитозина. Определите, сколько (в ) в этой молекуле содержится других нуклеотидов.
- В молекуле ДНК содержится цитозина. Определите, сколько (в ) в этой молекуле содержится других нуклеотидов.







к оглавлению ▴
Примеры задач второго типа
- В трансляции участвовало молекул т-РНК. Определите количество аминокислот, входящих в состав образующегося белка, а также число триплетов и нуклеотидов в гене, который кодирует этот белок.
- В трансляции участвовало молекул т-РНК. Определите количество аминокислот, входящих в состав образующегося белка, а также число триплетов и нуклеотидов в гене, который кодирует этот белок.
- В трансляции участвовало молекул т-РНК. Определите количество аминокислот, входящих в состав образующегося белка, а также число триплетов и нуклеотидов в гене, который кодирует этот белок.
- Фрагмент ДНК состоит из нуклеотидов. Определите число триплетов и нуклеотидов в иРНК, а также количество аминокислот, входящих в состав образующегося белка.
- Фрагмент ДНК состоит из нуклеотида. Определите число триплетов и нуклеотидов в иРНК, а также количество аминокислот, входящих в состав образующегося белка.
- Фрагмент ДНК состоит из нуклеотидов. Определите число триплетов и нуклеотидов в иРНК, а также количество аминокислот, входящих в состав образующегося белка.
- Фрагмент ДНК состоит из нуклеотидов. Определите число триплетов и нуклеотидов в иРНК, а также количество аминокислот, входящих в состав образующегося белка.
- Фрагмент ДНК состоит из нуклеотидов. Определите число триплетов и нуклеотидов в иРНК, а также количество аминокислот, входящих в состав образующегося белка.








к оглавлению ▴
Примеры задач третьего типа
- Фрагмент одной из цепей ДНК имеет следующее строение: ААГЦГТГЦТЦАГ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ЦЦАТАТЦЦГГАТ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: АГТТТЦТГГЦАА. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ГАТТАЦЦТАГТТ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ЦТАТЦЦГЦТГТЦ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ААГЦТАЦАГАЦЦ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ГГТГЦЦГГАААГ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ЦЦЦГТАААТТЦГ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
к оглавлению ▴
Примеры задач четвертого типа
- Фрагмент и-РНК имеет следующее строение: ГАУГАГУАЦУУЦААА. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: ЦГАГГУАУУЦЦЦУГГ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: УГУУЦААУАГГААГГ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: ЦЦГЦААЦАЦГЦГАГЦ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: АЦАГУГГЦЦААЦЦЦУ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: ГАЦАГАЦУЦААГУЦУ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: УГЦАЦУГААЦГЦГУА. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: ГЦАГГЦЦАГУУАУАУ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: ГЦУААУГУУЦУУУАЦ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
к оглавлению ▴
Примеры задач пятого типа
- Фрагмент ДНК имеет следующую последовательность нуклеотидов ТАТГГГЦТАТТГ. Установите нуклеотидную последовательность т-РНК, которая синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта т-РНК, если третий триплет соответствует антикодону т-РНК. Для решения задания используйте таблицу генетического кода.
- Фрагмент ДНК имеет следующую последовательность нуклеотидов ЦААГАТТТТГТТ. Установите нуклеотидную последовательность т-РНК, которая синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта т-РНК, если третий триплет соответствует антикодону т-РНК. Для решения задания используйте таблицу генетического кода.
- Фрагмент ДНК имеет следующую последовательность нуклеотидов ГЦЦАААТЦЦТГА. Установите нуклеотидную последовательность т-РНК, которая синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта т-РНК, если третий триплет соответствует антикодону т-РНК. Для решения задания используйте таблицу генетического кода.
- Фрагмент ДНК имеет следующую последовательность нуклеотидов ТГТЦЦАТЦАААЦ. Установите нуклеотидную последовательность т-РНК, которая синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта т-РНК, если третий триплет соответствует антикодону т-РНК. Для решения задания используйте таблицу генетического кода.
- Фрагмент ДНК имеет следующую последовательность нуклеотидов ЦАТГААААТГАТ. Установите нуклеотидную последовательность т-РНК, которая синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта т-РНК, если третий триплет соответствует антикодону т-РНК. Для решения задания используйте таблицу генетического кода.
к оглавлению ▴
Примеры задач шестого типа
- В клетке животного диплоидный набор хромосом равен . Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен . Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен . Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен . Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен . Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен . Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен . Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен . Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.








к оглавлению ▴
Примеры задач седьмого типа
- В диссимиляцию вступило молекул глюкозы. Определите количество АТФ после гликолиза, после энергетического этапа и суммарный эффект диссимиляции.
- В диссимиляцию вступило молекулы глюкозы. Определите количество АТФ после гликолиза, после энергетического этапа и суммарный эффект диссимиляции.
- В диссимиляцию вступило молекул глюкозы. Определите количество АТФ после гликолиза, после энергетического этапа и суммарный эффект диссимиляции.
- В диссимиляцию вступило молекулы глюкозы. Определите количество АТФ после гликолиза, после энергетического этапа и суммарный эффект диссимиляции.
- В цикл Кребса вступило молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.
- В цикл Кребса вступило молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.
- В цикл Кребса вступило молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.
- В цикл Кребса вступило молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.
- В цикл Кребса вступило молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.
- В цикл Кребса вступило молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.









к оглавлению ▴
Приложение I Генетический код (и-РНК)
| Первое основание | Второе основание | Третье основание | |||
| У | Ц | А | Г | ||
| У | Фен | Сер | Тир | Цис | У |
| Фен | Сер | Тир | Цис | Ц | |
| Лей | Сер | — | — | А | |
| Лей | Сер | — | Три | Г | |
| Ц | Лей | Про | Гис | Арг | У |
| Лей | Про | Гис | Арг | Ц | |
| Лей | Про | Глн | Арг | А | |
| Лей | Про | Глн | Арг | Г | |
| А | Иле | Тре | Асн | Сер | У |
| Иле | Тре | Асн | Сер | Ц | |
| Иле | Тре | Лиз | Арг | А | |
| Мет | Тре | Лиз | Арг | Г | |
| Г | Вал | Ала | Асп | Гли | У |
| Вал | Ала | Асп | Гли | Ц | |
| Вал | Ала | Глу | Гли | А | |
| Вал | Ала | Глу | Гли | Г |
к оглавлению ▴
Ответы
- А=. Г=Ц=.
- А=. Г=Ц=.
- Ц=. А=Т=.
- Ц=. А=Т=.
- Г=. А=Т=.
- Г=. А=Т=.
- аминокислот, триплетов, нуклеотидов.
- аминокислот, триплетов, нуклеотидов.
- аминокислот, триплетов, нуклеотидов.
- триплета, аминокислоты, молекулы т-РНК.
- триплетов, аминокислот, молекул т-РНК.
- триплет, аминокислота, молекула т-РНК.
- триплета, аминокислоты, молекулы т-РНК.
- триплетов, аминокислот, молекул т-РНК.
- и-РНК: УУЦ-ГЦА-ЦГА-ГУЦ. Аминокислотная последовательность: фен-ала-арг-вал.
- и-РНК: ГГУ-АУА-ГГЦ-ЦУА. Аминокислотная последовательность: гли-иле-гли-лей.
- и-РНК: УЦА-ААГ-ЦЦГ-ГУУ. Аминокислотная последовательность: сер-лиз-про-вал.
- и-РНК: ЦУА-АУГ-ГАУ-ЦАА. Аминокислотная последовательность: лей-мет-асп-глн.
- и-РНК: ГАУ-АГГ-ЦГА-ЦАГ. Аминокислотная последовательность: асп-арг-арг-глн.
- и-РНК: УУЦ-ГАУ-ГУЦ-УГГ. Аминокислотная последовательность: фен-асп-вал-три.
- и-РНК: ЦЦА-ЦГГ-ЦЦУ-УУЦ. Аминокислотная последовательность: про-арг-про-фен.
- и-РНК: ГГГ-ЦАУ-УУА-АГЦ. Аминокислотная последовательность: гли-гис-лей-сер.
- Фрагмент ДНК: ЦТАЦТЦАТГААГТТТ. Антикодоны т-РНК: ЦУА, ЦУЦ, АУГ, ААГ, УУУ. Аминокислотная последовательность: асп-глу-тир-фен-лиз.
- Фрагмент ДНК: ГЦТЦЦАТААГГГАЦЦ. Антикодоны т-РНК: ГЦУ, ЦЦА, УАА, ГГГ, АЦЦ. Аминокислотная последовательность: арг-гли-иле-про-три.
- Фрагмент ДНК: АЦААГТТАТЦЦТТЦЦ. Антикодоны т-РНК: АЦА, АГУ, УАУ, ЦЦУ, УЦЦ. Аминокислотная последовательность: цис-сер-иле-гли-арг.
- Фрагмент ДНК: ГГЦГТТГТГЦГЦТЦГ. Антикодоны т-РНК: ГГЦ, ГУУ, ГУГ, ЦГЦ, УЦГ. Аминокислотная последовательность: про-глн-гис-ала-сер.
- Фрагмент ДНК: ТГТЦАЦЦГГТТГГГА. Антикодоны т-РНК: УГУ, ЦАЦ, ЦГГ, УУГ, ГГА. Аминокислотная последовательность: тре-вал-ала-асн-про.
- Фрагмент ДНК: ЦТГТЦТГАГТТЦАГА. Антикодоны т-РНК: ЦУГ, УЦУ, ГАГ, УУЦ, АГА. Аминокислотная последовательность: асп-арг-лей-лиз-сер.
- Фрагмент ДНК: АЦГТГАЦТТГЦГЦАТ. Антикодоны т-РНК: АЦГ, УГА, ЦУУ, ГЦГ, ЦАУ. Аминокислотная последовательность: цис-тре-глу-арг-вал.
- Фрагмент ДНК: ЦГТЦЦГГТЦААТАТА. Антикодоны т-РНК: ЦГУ, ЦЦГ, ГУЦ, ААУ, АУА. Аминокислотная последовательность: ала-гли-глн-лей-тир.
- Фрагмент ДНК: ЦГАТТАЦААГАААТГ. Антикодоны т-РНК: ЦГА, УУА, ЦАА, ГАА, АУГ. Аминокислотная последовательность: ала-асн-вал-лей-тир.
- т-РНК: АУА-ЦЦЦ-ГАУ-ААЦ. Антикодон ГАУ, кодон и-РНК — ЦУА, переносимая аминокислота — лей.
- т-РНК: ГУУ-ЦУА-ААА-ЦАА. Антикодон ААА, кодон и-РНК — УУУ, переносимая аминокислота — фен.
- т-РНК: ЦГГ-УУУ-АГГ-АЦУ. Антикодон АГГ, кодон и-РНК — УЦЦ, переносимая аминокислота — сер.
- т-РНК: АЦА-ГГУ-АГУ-УУГ. Антикодон АГУ, кодон и-РНК — УЦА, переносимая аминокислота — сер.
- т-РНК: ГУА-ЦУУ-УУА-ЦУА. Антикодон УУА, кодон и-РНК — ААУ, переносимая аминокислота — асн.
- . Генетический набор:
- перед митозом молекул ДНК;
- после митоза молекулы ДНК;
- после первого деления мейоза молекул ДНК;
- после второго деления мейоза молекул ДНК.
- перед митозом
- . Генетический набор:
- перед митозом молекул ДНК;
- после митоза молекулы ДНК;
- после первого деления мейоза молекул ДНК;
- после второго деления мейоза молекул ДНК.
- перед митозом
- . Генетический набор:
- перед митозом молекул ДНК;
- после митоза молекулы ДНК;
- после первого деления мейоза молекул ДНК;
- после второго деления мейоза молекул ДНК.
- перед митозом
- . Генетический набор:
- перед митозом молекул ДНК;
- после митоза молекулы ДНК;
- после первого деления мейоза молекул ДНК;
- после второго деления мейоза молекул ДНК.
- перед митозом
- . Генетический набор:
- перед митозом молекул ДНК;
- после митоза молекулы ДНК;
- после первого деления мейоза молекул ДНК;
- после второго деления мейоза молекул ДНК.
- перед митозом
- . Генетический набор:
- перед митозом молекул ДНК;
- после митоза молекулы ДНК;
- после первого деления мейоза молекул ДНК;
- после второго деления мейоза молекул ДНК.
- перед митозом
- . Генетический набор:
- перед митозом молекул ДНК;
- после митоза молекулы ДНК;
- после первого деления мейоза молекул ДНК;
- после второго деления мейоза молекул ДНК.
- перед митозом
- . Генетический набор:
- перед митозом молекул ДНК;
- после митоза молекулы ДНК;
- после первого деления мейоза молекул ДНК;
- после второго деления мейоза молекул ДНК.
- перед митозом
- Поскольку из одной молекулы глюкозы образуется молекулы ПВК и АТФ, следовательно, синтезируется АТФ. После энергетического этапа диссимиляции образуется молекул АТФ (при распаде молекулы глюкозы), следовательно, синтезируется АТФ. Суммарный эффект диссимиляции равен АТФ.
- Поскольку из одной молекулы глюкозы образуется молекулы ПВК и АТФ, следовательно, синтезируется АТФ. После энергетического этапа диссимиляции образуется молекул АТФ (при распаде молекулы глюкозы), следовательно, синтезируется АТФ. Суммарный эффект диссимиляции равен АТФ.
- Поскольку из одной молекулы глюкозы образуется молекулы ПВК и АТФ, следовательно, синтезируется АТФ. После энергетического этапа диссимиляции образуется молекул АТФ (при распаде молекулы глюкозы), следовательно, синтезируется АТФ. Суммарный эффект диссимиляции равен АТФ.
- Поскольку из одной молекулы глюкозы образуется молекулы ПВК и АТФ, следовательно, синтезируется АТФ. После энергетического этапа диссимиляции образуется молекул АТФ (при распаде молекулы глюкозы), следовательно, синтезируется АТФ. Суммарный эффект диссимиляции равен АТФ.
- В цикл Кребса вступило молекул ПВК, следовательно, распалось молекулы глюкозы. Количество АТФ после гликолиза — молекул, после энергетического этапа — молекул, суммарный эффект диссимиляции молекул АТФ.
- В цикл Кребса вступило молекул ПВК, следовательно, распалось молекулы глюкозы. Количество АТФ после гликолиза — молекул, после энергетического этапа — молекул, суммарный эффект диссимиляции молекул АТФ.
- В цикл Кребса вступило молекул ПВК, следовательно, распалось молекул глюкозы. Количество АТФ после гликолиза — молекул, после энергетического этапа — молекул, суммарный эффект диссимиляции молекул АТФ.
- В цикл Кребса вступило молекул ПВК, следовательно, распалось молекул глюкозы. Количество АТФ после гликолиза — молекул, после энергетического этапа — молекул, суммарный эффект диссимиляции молекул АТФ.
- В цикл Кребса вступило молекул ПВК, следовательно, распалось молекул глюкозы. Количество АТФ после гликолиза — молекул, после энергетического этапа — молекул, суммарный эффект диссимиляции молекул АТФ.
- В цикл Кребса вступило молекул ПВК, следовательно, распалось молекул глюкозы. Количество АТФ после гликолиза — молекул, после энергетического этапа — молекул, суммарный эффект диссимиляции молекул АТФ.
























































Спасибо за то, что пользуйтесь нашими материалами.
Информация на странице «Задача С5 ЕГЭ по Биологии. Подборка заданий по цитологии» подготовлена нашими авторами специально, чтобы помочь вам в освоении предмета и подготовке к экзаменам.
Чтобы успешно сдать нужные и поступить в высшее учебное заведение или колледж нужно использовать все инструменты: учеба, контрольные, олимпиады, онлайн-лекции, видеоуроки, сборники заданий.
Также вы можете воспользоваться другими статьями из данного раздела.
Публикация обновлена:
09.03.2023
Цитология (греч. cytos — клетка + logos — наука) — наука о строении и жизнедеятельности клетки. На данный момент нам
кажется очевидным, что растения, грибы и животные состоят из клеток, однако раньше об этом и не догадывались.
Цитология начала свой путь развития относительно недавно, в этой статье мы обсудим клеточную теорию и методы,
которые используются в цитологии для изучения клеток (методологию).

Клеточная теория
Создание и развитие клеточной теории стало возможным после изобретения микроскопа в 1590 году голландским мастером по
изготовлению очков — Захарием Янсеном. Первый микроскоп мог увеличивать изучаемый объект до 3-9 раз.

В 1665 году Роберт Гук, используя микроскоп собственного изобретения, смог различить ячеистые структуры пробки ветки
бузины. Эти ячеистые структуры напомнили Роберту Гуку монашеские кельи, он ввел термин клетка (от лат. сеllа — комната, келья).
На самом деле Роберт Гук увидел не живые клетки, как он предполагал, а оставшиеся от них плотные клеточные стенки, которые и представляли собой ячеистую структуру.

В 70-х годах XVII века нидерландский натуралист Антони ван Левенгук открыл целый мир, невидимый невооруженным глазом. Он
увидел в микроскопе простейшие организмы: инфузорий, сперматозоидов, а также дрожжи, бактерии, эпидермис кожи.
В течение 50 лет он отсылал результаты своих наблюдений в Лондонское королевское общество. Поначалу они были встречены со скептицизмом,
но когда комиссия ученых лично во всем убедилась и подтвердила подлинность его исследований, Антони ван Левенгук был избран
действительным членом Лондонского королевского общества.

В последующее время было много описаний самых разных клеток, однако обобщить накопленный материал оказалось не легкой
задачей. С ней в 1839-1840 годах справились немецкий ботаник Маттиас Шлейден и немецкий зоолог Теодор Шванн.
Изучая строение растений и животных, Шлейден и Шванн независимо друг от друга пришли к одному и тому же выводу: все
организмы, как растительные, так и животные, состоят из клеток, сходных по строению. Они постулировали, что все живое
состоит из клеток. В 1839-1840 годах возникла клеточная теория Шлейдена и Шванна, основные положения которой:
- Все организмы состоят из клеток
- Клетка — мельчайшая структурная единица жизни
- Образование новых клеток — основополагающий способ роста и развития растений и животных
- Организм представляет собой сумму образующих его клеток
Допустили ли Шлейден и Шванн ошибки? Да, они были. Ошибочно предположение о том, что клетка может образоваться из
неклеточного вещества.
Важное дополнение в 1855 в клеточную теорию внес Рудольф Вирхов, который утверждал, что любая клетка
может образоваться только путем деления материнской клетки.

Какие же положения включает в себя современная клеточная теория? Приступим к их изучению:
- Клетка является структурной, функциональной и генетической единицей живого
- Клетки растений и животных сходны между собой по строению и химическому составу
- Клетка образуется только путем деления материнской клетки
- Клетки у всех организмов окружены мембраной (имеют мембранное строение)
- Ядро клетки — ее главный регуляторный органоид
- Клеточное строение растений, животных и грибов свидетельствует о едином происхождении всего живого
- В многоклеточном организме клетки подразделяются (дифференцируются) по строению и функции. Они объединяются в
ткани, органы и системы органов. - Клетка — элементарная, открытая и живая система, способная к самообновлению, воспроизведению и саморегуляции
XX век несомненно стал веком биологических наук: цитологии, генетики. Это произошло во многом благодаря клеточной
теории.
Я хочу поделиться с вами моим искренним восхищением новой жизни. Вдумайтесь — мы ведь когда-то с вами были всего
одной единственной клеткой, зиготой! Как в одной клетке природе удалось уместить столько всего: кожу, мышцы, нервную
систему, пищеварительный тракт? Мы приоткроем завесу этой тайну в статьях по генетике и эмбриологии, и, тем не менее, мое
восхищение этим безгранично.
Наши клетки рождаются и умирают: эпителий кишечника обновляется каждые 5 дней полностью,
при удалении 70% печени оставшиеся клетки способны восстановить всю структуру этого органа, каждые 30 дней мы получаем новую кожу.
При этом наше сознание и память остаются с нами. Мы — чудо, настоящее чудо природы, созданное из одной единственной клетки.

Микроскопия
Микроскопия — важнейший метод цитологии, в ходе которого объекты рассматриваются при помощи микроскопа. Его оптическая система состоит
из двух основных элементов: объектива и окуляра, закрепленных в тубусе. Микропрепарат (срез тканей) располагается
на предметном столике, расстояние от которого до объектива регулируется с помощью винта (винтов).
Чтобы посчитать увеличительную способность микроскопа следует умножить увеличение окуляра на увеличение объектива. К примеру,
если окуляр увеличивает объект в 20 раз, а объектив — в 10, то суммарное увеличение будет в 200 раз.

Некоторое внимание уделим направлениям в биологии, которые необходимо знать на современном этапе технического прогресса.
Биоинженерия
Биоинженерия — направление науки и техники, развивающее применение инженерных принципов в биологии и медицине. В рамках
биоинженерии происходят попытки (и довольно успешные) выращивания тканей и создание искусственных органов, протезов.
То есть биоинженерия занимается преимущественно технической частью. Медицинское направление в биоинженерии ищет замену
органам и тканям человека, которые утратили свою функциональную активность и требуют «замены».
Биотехнология
Биотехнология — направление биологии, изучающее возможность применения живых организмов или продуктов их жизнедеятельности
для решения технологических задач. В биотехнологии путем генной инженерии создают организмы с заданным набором свойств.
В рамках биотехнологии происходит получение антибиотиков — продуктов жизнедеятельности бактерий, очищение водоемов с помощью моллюсков, увеличение плодородия почвы с помощью дождевых червей, клонирование организмов.
Это разительно отличается от задач биоинженерии, хотя безусловно, эти дисциплины смежные. Все-таки в биотехнологии происходит большее вторжение в живой мир, по сути человек выступает эксплуататором, достигая с помощью животных, растений и грибов своих целей.
Человек проводит искусственный отбор, отделяя особей, которые продолжат род, от других, «менее перспективных».

В рамках биотехнологии выделяются следующие направления:
- Генная инженерия
- Сорт кукурузы, устойчивый к действию насекомых-вредителей
- Бактерии, продуктом жизнедеятельности которых является человеческий инсулин, используемый в дальнейшем как лекарство
- Культура клеток, вырабатывающих гормон человека — эритропоэтин, также используемый в лечебных целях
- Клеточная инженерия
Представляет собой совокупность методов и технологий, которые приводят к получению рекомбинантных РНК и ДНК,
выделению генов из клеток и внедрения их в другие организмы.
Изменив молекулу ДНК или РНК, человек добивается своей цели: клетка начинает синтезировать с нее белок. Он то и нужен человеку,
такие продукты жизнедеятельности активно используются в медицине, к примеру, при изготовлении антибиотиков.
В ходе генной инженерии был получены:

Представляет собой совокупность методов и технологий, используемых для конструирования новых клеток. В основе лежит
идея культивирования клеток тканей вне организма.
С помощью клеточной инженерии возможно бесполое размножение ценных форм растений. Часто получаются, так называемые,
гибридные клетки, которые сочетают свойства, к примеру, раковых клеток и лимфоцитов, в результате становится возможно
быстрое получение антител.

© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.