
«Биология отрицает законы математики: при делении происходит умножение» Валерий Красовский
Теория симбиогенеза (симбиотическая теория, эндосимбиотическая теория, теория эндосимбиоза) объясняет механизм возникновения некоторых органоидов эукариотической клетки — митохондрий и пластид – путем симбиоза.
Доказательства:
- Общее сходство по форме, размеру, ультраструктурой мембран митохондрий и бактерий, специфическими ферментными комплексами, хлоропластов и синезеленых водорослей.
- Кольцевая молекула ДНК.
- Образование путем бинарного деления независимо от клетки. Если эукариотные клетки лишить митохондрий или хлоропластов, то они не появляются вновь, эти органоиды передаются только от исходной материнской клетки путем деления предшествующих.
- Сходство на ультраструктурном и биохимическом уровне, например присутствие тилакоидов и специфических хлорофиллов, очень напоминают таковые образования в цианобактерий.
- Наличие собственного белкового синтеза.
- Мелкие рибосомы прокариотического типа.
- Движение митохондрий напоминает движение некоторых бактерий.
- Ферменты белкового синтеза у митохондрий, хлоропластов и бактерий, с одной стороны, и в цитоплазме – с другой, чувствительны к разным антибиотикам.
- Двуслойная мембрана: внутренний слой сходен по составу с бактериальными клетками, а внешний — с мембранами пищеварительных вакуолей эукариот (это не случайно, ведь бактерия была съедена эукариотической клеткой, а значит, оказалась в пищеварительной вакуоли).

Просмотров: 18619
Последние обновления
Последние видео:
Подписывайся на обновления, обсуждай вопросы в соцсетях


Какие черты сходства митохондрий с прокариотами позволили выдвинуть симбиотическую теорию происхождения эукариотической клетки?
Спрятать пояснение
Пояснение.
1) Имеют две полностью замкнутые мембраны. При этом внешняя сходна с мембранами вакуолей, внутренняя — бактерий.
2) Размножаются бинарным делением (причем делятся иногда независимо от деления клетки).
3) Генетический материал — кольцевая ДНК, не связанная с гистонами, имеют свой аппарат синтеза белка — рибосомы и др. Рибосомы прокариотического типа.
Спрятать критерии
Критерии проверки:
| Критерии оценивания ответа на задание С4 | Баллы |
|---|---|
| Ответ включает все названные выше элементы, не содержит биологических ошибок | 3 |
| Ответ включает 2 из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает 3 из названных выше элементов, но содержит негрубые биологические ошибки | 2 |
| Ответ включает 1 из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает 2 из названных выше элементов, но содержит негрубые биологические ошибки | 1 |
| Ответ неправильный | 0 |
| Максимальное количество баллов | 3 |
Раздел: Основы эволюционного учения
Симбиогенез
Исследования ДНК показывают, что первыми формами жизни были прокариоты, бактерии и археи, клетки которых довольно мелки и примитивны. Теория симбиогенеза предполагает, что более крупные и сложные клетки, протисты и многоклеточные организмы появились, когда прокариоты стали объединяться в группы.
Симбиоз — феномен, когда два разных вида извлекают выгоду от сосуществования. В 1967 г. американский генетик Линн Маргулис выдвинула удивившую многих теорию о том, что эукариотические клетки с их сложными внутренними структурами развились из неродственных им прокариотов, образовавших группы. Митохондрии и хлоропласты в эукариотических клетках, как заметили еще в 1910 г., похожи на бактерии — отсюда и вывод. Более поздние открытия показали, что эти органеллы содержат собственные ДНК.
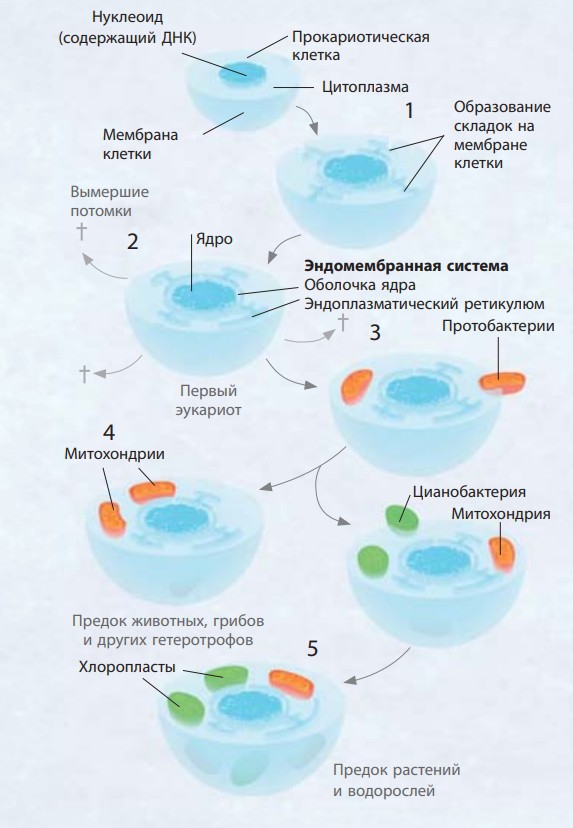
Симбиогенез потенциально может включать следующие этапы. 1) Прокариотическая клетка, вероятно архея, увеличивается, на ее мембране формируются складки, за счет которых растет площадь ее поверхности. 2) Складки отделяются от мембраны клетки и формируют мембрану вокруг ядра, где хранится генетический материал, — так образуется эндоплазматический ретикулум. 3) Терпимые к кислороду несерные пурпурные бактерии проникают внутрь и выживают. Они способны размножаться и сохраняться при делении клетки. 4) Бактерия использует кислород как средство производства энергии для клетки — как примитивная митохондрия. 5) Цианобактерия попадает в клетку и начинает производить сахара с помощью фотосинтеза. Клетка приобретает сходство с растительной
Анализ ДНК митохондрий, которые находят почти во всех живых клетках, доказал их сходство с протобактериями, также известными как несерные пурпурные бактерии. Это многочисленная группа прокариотов: некоторые из них вызывают заболевания, а прочие — это азотфиксирующие бактерии в почвах. Хлоропласты в клетках растений в свою очередь близко связаны с цианобактериями, или сине-зелеными водорослями. Это были первые фотосинтезирующие организмы.
Исключение из правил
Идея Маргулис известна как симбиогенез, или эндосимбиотическая теория, ведь составные элементы клетки зависят друг от друга. Но, возможно, все начиналось совсем не так. Инородные микроорганизмы могла поглотить более крупная клетка в качестве пищи, они также могли быть паразитами. ДНК в ядре эукариотов ближе к ДНК археи, а не бактерии, поэтому предполагается, что клетка, которая сформировала укрытие для эндосимбионта, была археей, мембрана которой разрослась и усложнилась.
Первая эукариотическая клетка появилась по меньшей мере 1,5 млрд лет назад. Считается, что этот процесс увенчался успехом только однажды. Все протисты, грибы, растения и животные произошли от одной этой клетки. Считается, что первый эукариот кормился другими организмами и не использовал фотосинтез. Он дал начало грибам и животным. Фотосинтезирующий хлоропласт возник позже, и с него началась история растений.
Поделиться ссылкой
На чтение 6 мин Просмотров 2.5к. Опубликовано 15.07.2021
Содержание
- Симбиогенез
- История cимбиогенеза
- Симбиотическое происхождение митохондрий и пластид
- Доказательства симбиогенеза
- Проблемы симбиогенеза
- Примеры эндосимбиозов
- Гипотезы эндосимбиотического происхождения других органелл
- Жгутики и реснички
- Пероксисомы
Симбиогенез
Теория симбиогенеза (симбиотическая теория, эндосимбиотическая теория, теория эндосимбиоза) объясняет механизм возникновения некоторых органоидов эукариотической клетки — митохондрий, гидрогеносом и пластид.
Схема эволюции эукариотических клеток. 1 — образование двойной мембраны ядра, 2 — приобретение митохондрий, 3 — приобретение пластид, 4 — внедрение получившейся фотосинтезирующей эукариотической клетки в нефотосинтезирующую (например, в ходе эволюции криптофитовых водорослей), 5 — внедрение получившейся клетки снова в нефотосинтезирующую (например, при симбиозе этих водорослей с инфузориями). Цветом обозначен геном предков эукариот, митохондрий и пластид.
История cимбиогенеза
Теорию эндосимбиотического происхождения хлоропластов впервые предложил в 1883 году Андреас Шимпер, показавший их саморепликацию внутри клетки. Её возникновению предшествовал вывод А. С. Фаминцина и О. В. Баранецкого о двойственной природе лишайников — симбиотического комплекса гриба и водоросли (1867 год). К. С. Мережковский в 1905 году предложил само название «симбиогенез», впервые детально сформулировал теорию и даже создал на её основе новую систему органического мира. Фаминцин в 1907 году, опираясь на работы Шимпера, также пришёл к выводу, что хлоропласты являются симбионтами, как и водоросли в составе лишайников.
В 1920-е годы теория была развита Б. М. Козо-Полянским, было высказано предположение, что симбионтами являются и митохондрии. Затем долгое время о симбиогенезе практически не упоминали в научной литературе. Второе рождение расширенная и конкретизированная теория получила уже в работах Линн Маргулис начиная с 1960-х годов.
Симбиотическое происхождение митохондрий и пластид
В результате изучения последовательности оснований в митохондриальной ДНК были получены весьма убедительные доводы в пользу того, что митохондрии — это потомки аэробных бактерий (прокариот), родственных риккетсиям, поселившихся некогда в предковой эукариотической клетке и «научившимися» жить в ней в качестве симбионтов. Теперь митохондрии есть почти во всех эукариотических клетках, размножаться вне клетки они уже не способны.
Существуют свидетельства того, что первоначально эндосимбиотические предки митохондрий не могли ни импортировать белки, ни экспортировать АТФ. Вероятно, первоначально они получали от клетки-хозяина пируват, а выгода для хозяина состояла в обезвреживании аэробными симбионтами токсичного для нуклеоцитоплазмы кислорода.
Пластиды, подобно митохондриям, имеют свои собственные прокариотические ДНК и рибосомы. По-видимому, хлоропласты произошли от фотосинтезирующих бактерий, поселившихся в своё время в гетеротрофных клетках протистов, превратив их в автотрофные водоросли.
Доказательства симбиогенеза
Митохондрии и пластиды:
- имеют две полностью замкнутые мембраны. При этом внешняя сходна с мембранами вакуолей, внутренняя — бактерий.
- размножаются бинарным делением (причём делятся иногда независимо от деления клетки), никогда не синтезируются de novo.
- генетический материал — кольцевая ДНК, не связанная с гистонами (По доле ГЦ ДНК митохондрий и пластид ближе к ДНК бактерий, чем к ядерной ДНК эукариот)
имеют свой аппарат синтеза белка — рибосомы и др. - рибосомы прокариотического типа — c константой седиментации 70S. По строению 16s рРНК близки к бактериальной.
- некоторые белки этих органелл похожи по своей первичной структуре на аналогичные белки бактерий и не похожи на соответствующие белки цитоплазмы.
Проблемы симбиогенеза
- ДНК митохондрий и пластид, в отличие от ДНК большинства прокариот, содержат интроны.
- В собственной ДНК митохондрий и хлоропластов закодирована только часть их белков, а остальные закодированы в ДНК ядра клетки. В ходе эволюции происходило «перетекание» части генетического материала из генома митохондрий и хлоропластов в ядерный геном. Этим объясняется тот факт, что ни хлоропласты, ни митохондрии не могут более существовать (размножаться) независимо.
- Не решён вопрос о происхождении ядерно-цитоплазматического компонента (ЯЦК), захватившего прото-митохондрии. Ни бактерии, ни археи не способны к фагоцитозу, питаясь исключительно осмотрофно. Молекулярно-биологические и биохимические исследования указывают на химерную архейно-бактериальную сущность ЯЦК. Как произошло слияние организмов из двух доменов, также не ясно.
Примеры эндосимбиозов
В наши дни существует ряд организмов, содержащих внутри своих клеток другие клетки в качестве эндосимбионтов. Они, однако, не являются сохранившимися до наших дней первичными эукариотами, у которых симбионты еще не интегрировались в единое целое и не потеряли своей индивидуальности. Тем не менее, они наглядно и убедительно показывают возможность симбиогенеза.
- Mixotricha paradoxa — наиболее интересный с этой точки зрения организм. Для движения она использует более 250 000 бактерий Treponema spirochetes, прикреплённых к поверхности её клетки. Митохондрии у этого организма вторично потеряны, но внутри его клетки есть сферические аэробные бактерии, заменяющие эти органеллы.
- Амёбы рода Pelomyxa также не содержат митохондрий и образуют симбиоз с бактериями.
- Инфузории рода Paramecium постоянно содержат внутри клеток водоросли, в частности, Paramecium bursaria образует эндосимбиоз с зелёными водорослями рода хлорелла (Chlorella).
- Одноклеточная жгутиковая водоросль Cyanophora paradoxa содержит цианеллы — органоиды, напоминающие типичные хлоропласты красных водорослей, но отличающиеся от них наличием тонкой клеточной стенки, содержащей пептидогликан (размер генома цианелл такой же, как у типичных хлоропластов, и во много раз меньше, чем у цианобактерий).
Гипотезы эндосимбиотического происхождения других органелл
Эндосимбиоз — наиболее широко признанная версия происхождения митохондрий и пластид. Но попытки объяснить подобным образом происхождение других органелл и структур клетки не находят достаточных доказательств и наталкиваются на обоснованную критику.
Клеточное ядро, нуклеоцитоплазма
Смешение у эукариот многих свойств, характерных для архей и бактерий, позволило предположить симбиотическое происхождение ядра от метаногенной архебактерии, внедрившейся в клетку миксобактерии. Гистоны, к примеру, обнаружены у эукариот и некоторых архей, кодирующие их гены весьма схожи. Другая гипотеза, объясняющая сочетание у эукариот молекулярных признаков архей и эубактерий, состоит в том, что на некотором этапе эволюции похожие на архей предки нуклеоцитоплазматического компонента эукариот приобрели способность к усиленному обмену генами с эубактериями путём горизонтального переноса генов.
В последнее десятилетие сформировалась также гипотеза вирусного эукариогенеза. В её основании лежит ряд сходств устройства генетического аппарата эукариот и вирусов: линейное строение ДНК, её тесное взаимодействие с белками и др. Было показано сходство ДНК-полимеразы эукариот и поксивирусов, что сделало именно их предков основными кандидатами на роль ядра.
Жгутики и реснички
Линн Маргулис в книге Symbiosis in Cell Evolution (1981) предположила в том числе происхождение жгутиков и ресничек от симбиотических спирохет. Несмотря на сходство размеров и строения указанных органелл и бактерий и существование Mixotricha paradoxa, использующей спирохет для движения, в жгутиках не было найдено никаких специфически спирохетных белков. Однако известен общий для всех бактерий и архей белок FtsZ, гомологичный тубулину и, возможно, являющийся его предшественником. Жгутики и реснички не обладают такими признаками бактериальных клеток, как замкнутая наружная мембрана, собственный белоксинтезирующий аппарат и способность к делению. Данные о наличии ДНК в базальных тельцах, появившиеся в 1990-е годы, были впоследствии опровергнуты. Увеличение числа базальных телец и гомологичных им центриолей происходит не путём деления, а путём достраивания нового органоида рядом со старым.
Пероксисомы
Кристиан де Дюв обнаружил пероксисомы в 1965 году. Ему же принадлежит предположение, что пероксисомы были первыми эндосимбионтами эукариотической клетки, позволившими ей выживать при нарастающем количестве свободного молекулярного кислорода в земной атмосфере. Пероксисомы, однако, в отличие от митохондрий и пластид, не имеют ни генетического материала, ни аппарата для синтеза белка. Было показано, что эти органеллы формируются в клетке de novo в ЭПР и нет никаких оснований считать их эндосимбионтами.
Мириады живых существ в экосистеме находятся в неисчислимом количестве связей с другими существами. Это
сложнейшая сеть взаимосвязей между организмами обеспечивает устойчивость экосистемы, служит предметом
интереснейшей науки — экологии.
Мы коснемся с вами этого уникального, хрупкого мира взаимосвязей живых существ. Среди форм взаимоотношений
между организмами выделяют:
Форма существования двух организмов, принадлежащих к разным видам. Некоторые организмы-симбионты никак не могут существовать друг без друга — облигатный симбиоз (лат. obligatus — обязанный). Примером облигатного
симбиоза могут служить лишайники, организмы, образованные симбиозом гриба и водоросли.

Иногда симбиоз между особями возможен, но не является обязательным условием. Если особи могут быть в симбиозе, а
могут и поодиночке, то такой симбиоз будет считаться факультативным (франц. facultatif — необязательный).
Известный пример факультативного симбиоза (протокооперации) — отношения между раком-отшельником и актинией. Актиния крепится к панцирю рака-отшельника, своими
щупальцами обездвиживает мелких животных, таким образом, достает пищу для себя и рака. Рак-отшельник постоянно перемещает актинию, за
счет чего вероятность ее встречи с потенциальной жертвой увеличивается.

В рамках симбиоза можно выделить мутуализм, комменсализм и паразитизм.
- Мутуализм (лат. mutual — взаимный)
- Комменсализм (лат. com — вместе + mensa — стол, трапеза)
- Паразитизм (греч. parasitos — нахлебник)
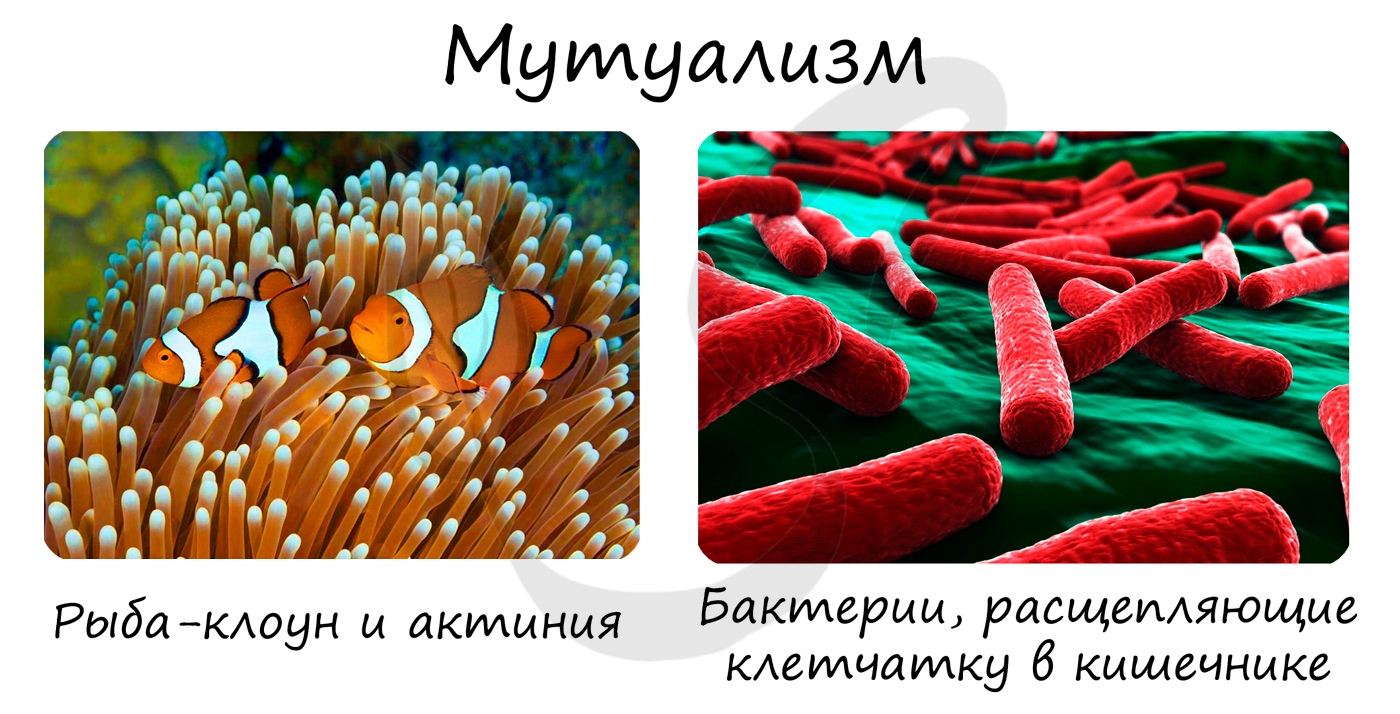
Форма взаимовыгодного облигатного симбиоза. Примером мутуализма могут послужить взаимоотношения между рыбой-клоуном и актинией. Рыба-клоун
спасается от врагов среди щупалец актинии, проводит там санитарную обработку: она удаляет из актинии непереваренные
остатки пищи, вентилирует воду.
Внутри пищеварительного тракта коровы происходит мутуализм с бактериями. Особая микрофлора заселяет отдел желудка — рубец. Именно здесь целлюлоза, которая не может быть разрушена пищеварительными ферментами коровы, переваривается бактериями-симбионтами. Без бактерий нормальное расщепление целлюлозы невозможно.
Комменсализм — способ симбиоза, при котором один из партнеров (комменсал) возлагает на другого (хозяина) регуляцию своих
взаимоотношений с внешней средой. При этом комменсал получает пользу от таких взаимоотношений, а хозяин не получает ни вреда,
ни пользы.
Примером таких взаимоотношений может послужить «квартиранство», при котором один из организмов использует другой как жилище: в мантийную полость
двустворчатых моллюсков откладывают икринки рыбы-горчаки, благодаря чему развивающиеся икринки надежно защищены раковиной моллюска, но не приносят
ни вреда, ни пользы самому моллюску.
Также примером является и «нахлебничество». Под этот термин подпадают отношения между акулой и рыбой-прилипалой. Рыба-прилипала (комменсал) прикрепляется
к акуле, преодолевает большие расстояния и питается остатками пищи, расплывающимися в стороны после трапезы акулы.

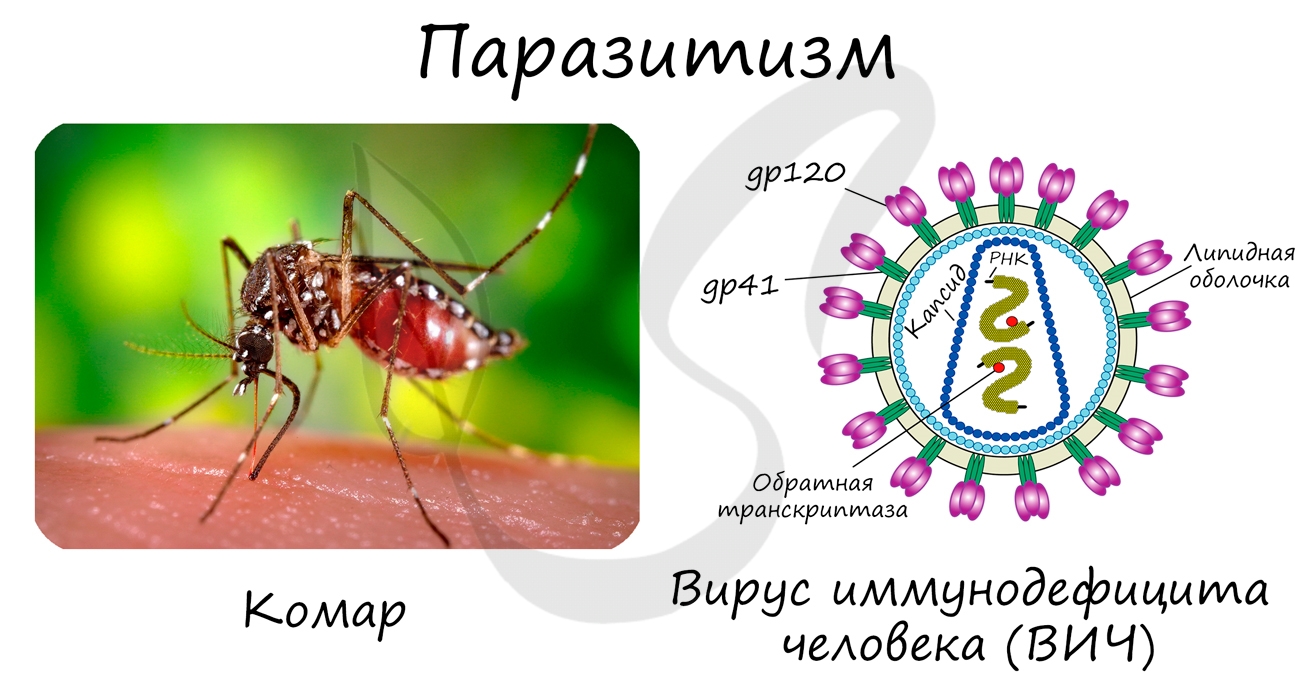
Паразитизм также является способом симбиоза. При этой форме отношений один организм (паразит) использует другой (хозяина) в качестве источника питания
(и среды обитания), при этом частично/полностью возлагая на него регуляцию своих отношений с внешней средой.
Паразитизм бывает облигатный, в случае если паразит не может жить без хозяина, к примеру, у вирусов. Может быть факультативный, если паразит способен
существовать без хозяина: комары, блохи, вши, паразитические черви.
В современной экологии в понятие хищничества вкладывается форма взаимоотношения, при которой один организм питается органами и тканями
другого, при этом между двумя организмами отсутствуют симбиотические связи. То есть они никак не зависят друг от друга.
Иногда понятие хищничества обобщается, и в него включают плотоядных, растительноядных, всеядных животных и паразитов.

При этой форме взаимоотношений виды не оказывают друг на друга практически никакого влияния. Они редко встречаются
из-за разности типов питания, экологических ниш.

Антибиозом называют такие взаимоотношения между видами, при которых один организм ограничивает возможности другого, иногда вплоть
до невозможности существования. Выделяют аменсализм, аллелопатию и конкуренцию.
- Аменсализм (греч. а — отрицательная частица + лат. mensa -стол, трапеза)
- Аллелопатия (греч. allelon — взаимно + pathos — страдание)
- Конкуренция (лат. concurrentia — столкновение)
При аменсализме один вид подавляет другой без извлечения выгоды для себя и без обратного отрицательного влияния с
подавляемой стороны. Примерами аменсализма являются высокие широкие кроны взрослых деревьев, которые практически
не пропускают свет в подлесок и тем самым угнеют рост молодых растений, мхов.

Аллелопатией называют подавление одного вида организмов другим (и обратное воздействие) вследствие выделения токсичных веществ.
Часто встречается у микроорганизмов, грибов.
Примером может считаться выделение антибиотиков двумя близкорасположенными бактериями. В этом случае антибиотик каждой бактерии
будет замедлять рост и развитие другой, может приводить к гибели.
Если у особей, принадлежащих к двум разным видам (или к одному), сходный образ жизни, кормовая база, занимаемая ими экологическая ниша,
ограниченные возможности для полового размножения: между ними возникает конкуренция.
Особенно часто возникает конкуренция между особями одного вида, ведь их потребности совершенно одинаковы. Недаром
самым ожесточенным вариантом борьбы за существование считается внутривидовая борьба.

Симбиотическая гипотеза в последнее время находит большое количество сторонников и часто называется уже не гипотезой, требующей доказательств, а теорией.
Следует отметить, что впервые идея о симбиотическом происхождении сложно устроенной клетки была высказана в России отечественным учёным-ботаником Андреем Сергеевичем Фаминцыным (1835-1918) в конце 60-х гг. XIX в. на основе изучения лишайников. В дальнейшем, в 1905-1909 гг., биолог Константин Сергеевич Мережковский (1855-1921) предложил термин «симбиоз» и выдвинул концепцию о симбиотическом происхождении носителей пигментов в растительных клетках. Эта идея была развита в 1921-1924 гг. в симбиотическую концепцию происхождения клеток растений и животных ботаником Борисом Михайловичем Козо-Полянским (1890-1957). На Западе эту концепцию развивали Лили Саган (1967), утверждавшая, что эукариотическая клетка возникла путём симбиоза двух или более прокариотических клеток, Линна Маргулис (1983) и другие учёные.
Симбиотическая, или эндосимбиотическая, гипотеза исходит из того, что эукариоты представляют собой результат симбиоза между различными прокариотами. Допускается, что ядро развилось вследствие обособления ДНК из цитоплазмы путём впячивания плазматической мембраны и обволакивания ядерного вещества. Сходным образом могли возникнуть и эндоплазматическая сеть, и аппарат Гольджи.
Однако считается, что появление митохондрий и хлоропластов произошло иным путём. Полагают, что некоторые клетки, будучи гетеротрофами, захватывали других, более мелких бактерий, которых они по неизвестным причинам не переваривали. Захваченные мелкие клетки прокариот были способны поглощать кислород. Это свойство было выгодным фактом для клетки-хозяина, поглотившей такую бактерию, так как давало 34 молекулы АТФ на одну молекулу глюкозы (против только двух молекул АТФ при брожении). Поглотителем могла быть и фототрофная клетка прокариот, питающаяся органическими веществами, у которой в процессе реакций обмена веществ мог образовываться молекулярный кислород, ядовитый для неё, но потребляемый клеткой-симбионтом для своего дыхания. Из этих сохранившихся бактерий-симбионтов в ходе дальнейшей эволюции и могли произойти митохондрии. В пользу этой гипотезы свидетельствуют данные о современных симбиотических отношениях между водорослями и инфузориями. На рис. 58 представлена схема возникновения в процессе эволюции сложной клетки эукариот с помощью разных путей симбиоза. Материал с сайта http://doklad-referat.ru
 |
| Рис. 58. 1 — схема возникновения эукариот согласно симбиотической гипотезе: а — прокариота I (будущее ядро); б — прокариота II (будущая митохондрия); в — прокариота III (будущий хлоропласт); г — первая эукариота с ядром и митохондрией; д-и — возможные пути дальнейшего усложнения структуры клетки эукариот; 2 — бактерия-симбионт с митохондриями (первая эукариотическая клетка) |
Одновременно с этими процессами или несколько позднее таким же путём мог произойти и эндоцитозный захват автотрофных прокариот, например сине-зелёных водорослей (цианобактерий), обладающих полным набором пигментов фотосинтеза. Из этих симбионтов развились хлоропласты. Допускается также, что некоторые сложные по строению клетки, например простейших, произошли в результате симбиотических связей не только с прокариотами, но и с другими эукариотами.
Симбиотическая гипотеза в настоящее время находит подтверждение рядом эволюционных реликтов в существующих и сейчас симбиотических связях между прокариотами и эукариотами. Такие связи наблюдаются среди растительных и животных организмов. Например, свободноживущая амёба Pelomyxa palustris (пеломикса болотная) не имеет митохондрий, но содержит в себе симбиотические бактерии, которые обеспечивают ей дыхание и АТФ.

На этой странице материал по темам:
-
Симбиотическая теория происхождения что это
-
Симбиотическая теория происхождения эукариотических клеток презентация
-
Согласно теории симбиотического присхождения первыми эукариотами были
-
10 класс биология симбиотическое происхождение
-
Гіпотези походження еукаріотів презентація