Эта группа клеток состоит из нескольких видов, также называемых субпопуляциями лимфоцитов. Долгое время выделяли только три их вида: это Т-лимфоциты хелперы, киллеры и супрессоры. Однако в последние годы, начиная с 1990-2000-х, представление об их существующих разновидностях претерпело очередную трансформацию. Помимо известных клеток, специалисты определили существование других типов: Т-клеток памяти и клеток-амплификаторов. Рассмотрим все существующие виды лимфоцитов подробнее.
Т-киллеры:
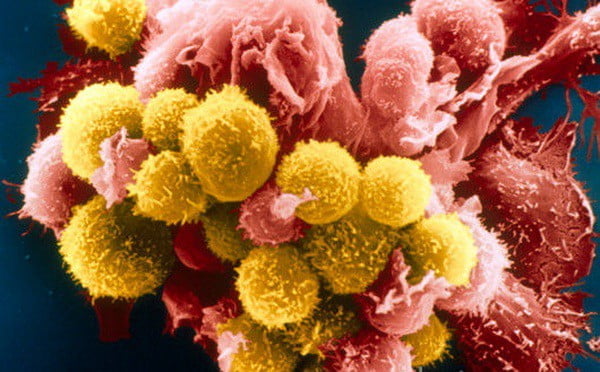 Т-киллеры – самая известная субпопуляция лимфоцитов. Они обладают способностью разрушать неполноценные клетки организма, вступая с ними в непосредственный контакт. Их еще называют цитотоксические лимфоциты: «цито» в переводе означает «клетка», значение слова «токсические» объяснять не надо.
Т-киллеры – самая известная субпопуляция лимфоцитов. Они обладают способностью разрушать неполноценные клетки организма, вступая с ними в непосредственный контакт. Их еще называют цитотоксические лимфоциты: «цито» в переводе означает «клетка», значение слова «токсические» объяснять не надо.
Т-киллеры, строго осуществляющие иммунный надзор, агрессивно реагируют на чужеродный белок. Именно они вызывают реакцию отторжения трансплантата при пересадке органов. По этой причине при пересадке человеку любого органа ему некоторое время дают специальные медикаменты, которые угнетают иммунную систему: уменьшают повышенное содержание лимфоцитов и нарушают их взаимодействие. Иначе любая подобная операция заканчивалась бы отторжением нового органа или ткани, а может, даже гибелью пациента, которому такое вмешательство проводится.
Интересен механизм работы этих клеток. В отличие от фагоцитов, активно атакующих, пожирающих и переваривающих чужеродные частицы, Т-киллеры ведут себя на первый взгляд достаточно сдержанно. Своими отростками они прикасаются к объекту, а затем разрывают контакт и «уходят по своим делам». Клетка же, к которой прикоснулся лимфоцит, спустя какое-то время погибает… Отчего?
Дело в том, что во время своего «смертельного поцелуя» Т-киллеры оставляют на поверхности уничтожаемой ими клетки частицы своей мембраны. В местах контакта частицы «разъедают» поверхность объекта нападения. В результате в обреченной на гибель клетке фактически образуется сквозное отверстие. Она теряет ионы калия, внутрь нее входят ионы натрия и вода – так как клеточный барьер нарушается, ее внутренняя среда начинает напрямую сообщаться с внешней… В исходе клетка раздувается проникшей внутрь ее водой, из нее выходят белки цитоплазмы, органеллы разрушаются… Она погибает, а дальше к ней подходят фагоциты и пожирают ее остатки. Вот такое страшное наказание готовит организм всем клеткам, которые были распознаны иммунитетом как «неправильные» или чужеродные.
Т-хелперы:
Задача хелперов на первый взгляд тоже довольно очевидна.  Это клетки-помощники («help» значит «помогать»). А кому или чему они помогают? Они индуцируют, стимулируют иммунный ответ: под их влиянием усиливают свою работу цитотоксические лимфоциты. Также хелперы передают информацию о присутствии в теле чужеродного белка В-лимфоцитам, которые выделяют против них защитные антитела. Наконец, хелперы оказывают стимулирующее действие на работу фагоцитов, главным образом моноцитов.
Это клетки-помощники («help» значит «помогать»). А кому или чему они помогают? Они индуцируют, стимулируют иммунный ответ: под их влиянием усиливают свою работу цитотоксические лимфоциты. Также хелперы передают информацию о присутствии в теле чужеродного белка В-лимфоцитам, которые выделяют против них защитные антитела. Наконец, хелперы оказывают стимулирующее действие на работу фагоцитов, главным образом моноцитов.
Т-хелперы
Т-супрессоры:
«Супрессия» означает «подавление». Если Т-лимфоциты хелперы усиливают реакцию иммунитета, то супрессоры, наоборот, подавляют. Притом эти клетки вовсе не устраивают саботаж иммунным процессам и не вредят нашему здоровью. Они просто регулируют силу иммунного ответа, что позволяет иммунной системе сдержанно и с умеренной силой отвечать на раздражители.
Лимфоциты-амплификаторы:
После того как в организм проник агрессор, в крови и тканях отмечается повышенное содержание лимфоцитов. Их число возрастает буквально в течение нескольких часов и может увеличиваться более чем в 2 раза. Отчего же нарастание количества клеток происходит так быстро? Просто в организме имеется их некоторый запас.
В селезенке и тимусе живут зрелые, полноценные лимфоциты. Их отличие от остальных состоит лишь в том, что они «не определились», к какому виду лимфоцитов принадлежат. Это и есть клетки-амплификаторы, при необходимости они участвуют в увеличении численности других Т-лимфоцитов.
Т-клетки памяти:
Справившись с очередной угрозой, лимфоциты ее запоминают. В организме человека образуется особый клон клеток, которые и хранят эти «воспоминания». Каждый клон несет в себе информацию об определенном виде угрозы. Если какой-то агрессор, с которым иммунная система уже встречалась, проникает в тело, то соответствующий клон размножается и быстро формирует вторичный иммунный ответ.
Разговор о разновидностях лимфоцитов и их функциях достаточно долгий. Здесь эта тема была представлена в максимально приемлемой и простой форме, без загрузки специфическими терминами и непонятными названиями. Будем надеяться, что любой читатель, даже не имеющий медицинского образования, примерно разобрался в том, как функционируют в его организме разные виды Т-лимфоцитов.
 Из всего этого можно сделать очевидный вывод: для того чтобы жить полноценной здоровой жизнью, надо быть обладателем крепкой иммунной системы. Необходимо, чтобы процессы, о которых многие не задумываются, а еще большее количество людей даже не знает, происходили как надо.
Из всего этого можно сделать очевидный вывод: для того чтобы жить полноценной здоровой жизнью, надо быть обладателем крепкой иммунной системы. Необходимо, чтобы процессы, о которых многие не задумываются, а еще большее количество людей даже не знает, происходили как надо.
Если природа не наградила вас устойчивым иммунитетом, вам следует задуматься о том, чтобы самостоятельно взяться за его укрепление. Для этого можно начать прием препарата Трансфер Фактор. В нем содержатся информационные молекулы, при помощи которых лимфоциты в норме общаются друг с другом, управляют различными процессами и координируют их. Восполняя недостаток естественных информационных молекул, средство является одним из самых рекомендуемых и эффективных препаратов для нормализации работы иммунитета, улучшения здоровья и профилактики заболеваний.
Клетки иммунитета обладают памятью
и передают друг другу информацию
Иммунитет
Иммунитетом называют способы защиты живого организма от воздействия разнообразных факторов (микроорганизмов, опасных веществ и пр.), которые могут негативно повлиять на него, нарушить гомеостаз, привести к «поломкам». Иммунитет в широком смысле — невосприимчивость организма к болезням. Достойный иммунный ответ осуществляют клетки-иммуноциты, которые все вместе составляют сложную и не до конца еще изученную иммунную систему.
Аллергия — высокий уровень восприимчивости иммунной системы к аллергенам, в качестве которых может выступать огромное количество веществ. Проявляется чиханием, насморком, отеками, кашлем, кожными высыпаниями и т. д.
Лейкоциты (белые кровяные тельца)
1. Образуются в красном костном мозге, тимусе, селезенке, лимфатических узлах. Стволовые клетки лейкоцитов находятся только в красном костном мозге, но могут размножаться в других лимфоидных органах: в тимусе, лимфоузлах.
2. Имеют ядро и цитоплазму.
3. Количество их в крови — от 4 до 9 тысяч на 1 кубический миллиметр.
4. Среди лейкоцитов есть такие, которые живут всего несколько дней, но есть и «аксакалы», возраст которых доходит до нескольких лет.
5. Легко проникают сквозь стенки капилляров и выходят в тканевую жидкость.
6. При клеточном иммунитете определенные лейкоциты (то есть клетки) сами нападают на вирусы и бактерии, уничтожая их.
7. Клеточный иммунитет, основанный на фагоцитозе, является более древним механизмом иммунитета, чем гуморальный.
8. При гуморальном иммунитете определенные лейкоциты выделяют особые белки (антитела), которые уничтожают чужеродных агентов (антигены).
Виды лейкоцитов. Лимфоциты
Иммунный «надзор» — распознавание и уничтожение микроорганизмов, чужеродных веществ, генетически измененных собственных клеток организма — осуществляется лимфоцитами Т и В. У детей до 50 процентов всех лейкоцитов составляют именно лимфоциты, у взрослых их 19–37 процентов.
Т-лимфоциты
1. Образуются в тимусе.
2. Т-киллеры, один из подвидов Т-лимфоцитов, прозванные «клетками-убийцами», уничтожают чужеродные клетки и собственные мутировавшие клетки организма, в том числе раковые, что является примером клеточного иммунитета.
3. При гуморальном иммунитете под воздействием антигенов формируются Т-хелперы (клетки-помощники). Они помогают В- лимфоцитам распознавать антигены.
4. Среди Т-лимфоцитов также есть Т-супрессоры, которые блокируют реакции В- лимфоцитов, осуществляя торможение затянувшихся иммунных реакций.
В-лимфоциты
1. Формируются в костном мозге.
2. Участвуют исключительно в реакциях гуморального иммунитета.
3. Имеют микроворсинки с иммуноглобулиновыми рецепторами, которые распознают чужеродные вещества — антигены.
4. На основе В-лимфоцитов создаются плазматические клетки (плазмоциты), которые начинают сильно делиться после контакта с антигенами. Они синтезируют различные антитела-иммуноглобулины, выбрасывают их в кровь, тканевую жидкость, лимфу, секреты желез.
5. В-лимфоцит тоже имеет на своей поверхности антитела, однако от плазмоцитов антитела уже свободно перемещаются. Иммуноглобулины нейтрализуют антигены.
6. Антитело связываются с чужеродным агентам — антигеном (вирусом, бактерией и др.), образуя комплекс «антиген–антитело», в котором антиген инактивируется.
7. После взаимодействия с антигеном В-лимфоциты могут превращаться не только в плазмоциты, но и в так называемые «клетки памяти», задача которых — помнить всё, обеспечивать приобретенный иммунитет.
8. «Клетки памяти» из крови двигаются в лимфоидные ткани и органы, где остаются десятилетиями, до встречи с определенными антигенами.
Выводы: Т-лимфоциты в основном обеспечивают клеточный иммунитет
(за счет Т-киллеров), В-лимфоциты участвуют в гуморальном иммунитете, выделяя антитела (иммуноглобулины) против антигенов. Вместе с тем, Т-хелперы особым способом также принимают участие в гуморальном иммунитете, помогая В-лимфоцитам распознавать антигены.
Хочешь сдать экзамен на отлично? Жми сюда — курсы по биологии для подготовки к ОГЭ
Софья Касацкая
«Природа» №2, 2016

Об авторе
Софья Алексеевна Касацкая — младший научный сотрудник лаборатории геномики адаптивного иммунитета Института биоорганической химии им. академиков М. М. Шемякина и Ю. А. Овчинникова РАН. Область научных интересов — Т-клеточный иммунитет, нейро- и онкоиммунология. Победитель конкурса «Био/мол/текст» 2015 г. в номинации «Лучшая статья по иммунологии».
Адекватная защитная реакция организма на вторжение вирусов, бактерий и других патогенов — уничтожить пораженные клетки, не допустив распространения инфекции и гибели большого числа собственных клеток. Если инфицированная вирусом клетка его заметила, запускаются процессы врожденного иммунитета: аутофагия (утилизация внутренних компонентов клетки с помощью ферментов лизосом) и апоптоз (запрограммированная гибель клеток). Однако патогенных вирусов и бактерий очень много, к тому же они постоянно меняются до неузнаваемости. Чтобы справиться с ними, подключается система адаптивного иммунитета и его главные участники — лимфоциты. Вершиной эволюции адаптивного иммунитета стал цитотоксический Т-лимфоцит, или Т-киллер. Для распознавания фрагмента вируса (антигена) на зараженной клетке он использует Т-клеточный рецептор (T cell receptor, TCR), случайно и независимо собирающийся на каждой Т-клетке в вилочковой железе (тимусе). Механизм сборки TCR уникален и присущ только иммунной системе позвоночных животных. Считается, что эти преимущества впервые получили примитивные рыбы около 500 млн лет назад, когда в результате ретровирусной инфекции в их гаметы внедрились гены, кодирующие особые белки (рекомбиназы), ответственные за рекомбинацию генов TCR.
Классическая иммунология человека построена на изучении иммунных клеток крови просто потому, что образец крови можно взять у любого пациента и исследовать в норме и в патологии. Именно на клетках крови была выстроена классификация Т-лимфоцитов — деление на Т-киллеры и Т-хелперы, которые проверяют антигенную специфичность Т-киллеров, выдают им «лицензию на убийство» и способны управлять всем ходом иммунного ответа через сигнальные растворимые молекулы, цитокины. Позднее из ветви Т-хелперов была выделена группа регуляторных Т-клеток, подавляющих избыточный адаптивный иммунитет.
Но, как нам напоминает реклама йогурта, значительная часть клеток иммунной системы сосредоточена вокруг слизистой оболочки пищеварительного тракта и в других тканях. В то время как в 5–6 л крови взрослого человека находится около 6–15 млрд T-лимфоцитов, в эпидермисе и коже — 20 млрд Т-клеток [1], в печени — еще 4 млрд [2]. Достаточно ли изучения образцов крови для полного описания функций Т-клеток, если в периферических органах Т-клеток больше, чем в кровотоке? И достаточно ли классических субпопуляций, чтобы описать все типы Т-клеток, находящихся в организме человека?
Жизненный цикл Т-лимфоцита
Каждая Т-клетка после сборки TCR проходит тестирование на функциональность случайно собранного рецептора (положительная селекция) и на отсутствие специфичности к собственным антигенам организма, т.е. на отсутствие очевидной аутоиммунной угрозы (отрицательная селекция). Этапы селекции происходят в вилочковой железе; при этом более 90% клеток-предшественников погибает, не сумев правильно собрать рецептор либо пройти селективный отбор. Выжившие Т-клетки пролиферируют и выходят из тимуса в кровоток — это наивные Т-лимфоциты, еще не встречавшиеся с антигеном. Наивная Т-клетка циркулирует в крови и периодически заходит в лимфоузлы, где в Т-клеточной зоне контактирует со специализированными клетками, которые представляют ей чужеродный антиген.

После встречи с антигеном в лимфоузле Т-клетка приобретает способность снова делиться — становится предшественницей Т-клеток памяти (Stem Cell Memory T cells, TSCM). Cреди ее потомков появляются клетки центральной памяти (Central Memory T cells, TCM) и эффекторные клетки-предшественники (Effector Memory T cells, TEM), которые при делении дают короткоживущие эффекторные клетки, осуществляющие иммунную реакцию (TEMRA-клетки) [3]. Все эти клетки выходят из лимфоузла и перемещаются по крови. Эффекторные клетки затем могут покинуть кровоток для осуществления иммунной реакции в периферической ткани органа, где находится патоген. Что потом — снова путешествие по крови и лимфоузлам?

Клетки стромы, т.е. основы лимфоузла, выделяют сигнальные вещества (хемокины) для того, чтобы позвать Т-клетку в лимфоузел. Распознают хемокины лимфоузлов рецепторы хоминга CCR7 и CD62L. Но на эффекторных клетках оба рецептора отсутствуют. Из-за этого долгое время было загадкой, как эффекторные клетки могут попасть из периферической ткани обратно во вторичные лимфоидные органы — селезенку и лимфоузлы.

![]()
В то же время стали накапливаться данные (о различиях в репертуарах TCR и профилях транскрипции между TEM в крови и в других тканях), которые никак не укладывались в концепцию постоянной миграции Т-клеток между тканями и кровью. Решено было выделить новую субпопуляцию — резидентные клетки памяти (Resident Memory T cells, TRM), которые населяют определенный орган и не рециркулируют [5].
Откуда впервые появляются резидентные клетки ткани? Это потомки эффекторных клеток, которые потеряли способность рециркулировать. Некоторые периферические для иммунной системы ткани, например слизистая тонкого кишечника и брюшная полость, позволяют эффекторным Т-лимфоцитам проникать внутрь свободно, другие — очень ограниченно. Большой поток эффекторных Т-клеток в эти ткани наблюдается только при реакции воспаления. К тканям второго типа относятся головной и спинной мозг, отделенные барьером от иммунной системы, а также многие другие ткани: периферические ганглии, слизистые половых органов и кишечника, легкие, эпидермис, глаза. Разница между двумя типами тканей — в экспрессии дополнительных молекул хоминга для эффекторных Т-клеток, например молекул адгезии MadCAM-1 для проникновения в эпителий [3].
Резидентные Т-клетки в старении тканей человека
Карта соотношений присутствия отдельных субпопуляций Т-клеток в разных органах человека, как ни странно, была составлена только в 2014 г. Команда Донны Фарбер из медицинского центра Колумбийского университета Нью-Йорка провела сравнение фенотипов Т-клеток, выделенных из крови и тканей доноров органов всех возрастных групп от 3 до 73 лет (всего 56 доноров) [6]. Анализ субпопуляций Т-клеток при помощи проточной цитофлуориметрии подтвердил многие данные, полученные методами с меньшим разрешением и меньшей статистикой, и некоторые черты описания иммунной системы, перенесенные с иммунологии мыши на человека, к примеру снижение содержания наивных Т-лимфоцитов во всех органах при старении организма.
Уменьшение числа наивных Т-клеток с возрастом связано с быстрым старением вилочковой железы, в которой будущие Т-клетки проходят этапы сборки TCR, проверку его работоспособности и селекцию на отсутствие аутоиммунного потенциала. Важно не только снижение абсолютной численности наивных Т-клеток, но и уменьшение разнообразия репертуара Т-клеточных рецепторов, а значит, и возможности сформировать адаптивный иммунный ответ на ранее незнакомую инфекцию [7]. Для наивных Т-киллеров подтвердилось прогрессирующее падение численности в крови и лимфоузлах, хотя для наивных Т-хелперов отрицательная корреляция численности с возрастом в данном исследовании оказалась значительной только для вторичных лимфоидных органов, но не для крови.

Выделение Т-лимфоцитов памяти, эффекторных клеток памяти и короткоживущих эффекторных клеток из слизистых легких, тонкого и толстого кишечника, паховых и мезентериальных лимфоузлов доноров органов позволило впервые оценить динамику данных популяций в тканях человека при старении. Доля центральных клеток памяти ожидаемо растет с течением жизни, в соответствии с ростом числа инфекций, которые успели встретиться организму и попасть в библиотеку памяти иммунной системы. Эффекторные клетки памяти TEM стремительно заполняют нишу для Т-клеток в тканях ребенка, быстро, примерно к 12 годам, вытесняя наивные Т-клетки. Растет и количество терминально дифференцированных эффекторных Т-киллеров (TEMRA), но только в лимфоузлах и в селезенке; в нелимфоидных тканях численность TEMRA падает. Короткоживущие терминально дифференцированные Т-киллеры чаще всего встречаются в крови, селезенке и слизистых легких в любом возрасте, а вот среди Т-хелперов эта субпопуляция представлена исчезающе малым числом клеток. Аналогично мало центральных клеток памяти среди Т-киллеров, преимущественно они находятся в слизистых двух барьерных тканей: легких и кишечника.
Широкими мазками карту распределения Т-лимфоцитов человека можно обрисовать так: наивные Т-клетки путешествуют по крови и периодически заходят во вторичные лимфоидные органы, киллеры TEMRA находятся в крови, селезенке и легких. Для центральных клеток памяти, судя по всему, характерно более индивидуальное распределение по тканям, чем для других субпопуляций: во всяком случае, закономерностей динамики при старении разных тканей выявить не удалось. Эффекторные клетки памяти, включающие и TRM-субпопуляцию, доминируют среди Т-клеток слизистых барьерных тканей. В целом, при старении Т-клеточного иммунитета нелимфоидные ткани проявляют большую стабильность субпопуляций, лимфоидные ткани — большую возрастную динамику типов Т-клеток [6]. Стабильность тканевых клеток проще объяснить, если разобраться, какие из эффекторных клеток TEM остаются в ткани, становятся резидентными TRM и из каких событий состоит их жизнь после отказа от путешествий по организму.
Как отличить резидентные клетки тканей от примесей клеток крови?
Резидентные Т-клетки корректно, но неудобно каждый раз определять по способности индивидуальной клетки мигрировать в лимфоузлы, поэтому необходимо составить список характерных признаков, по которым можно выявить принадлежность к этой субпопуляции. Резидентные Т-лимфоциты в тканях — естественных барьерах организма (например в легких и слизистой тонкого кишечника) немного похожи на классические эффекторные клетки крови: экспрессируют маркер активированных клеток CD69, причем экспрессия стабильна в течение жизни при взрослении и старении и характерна для всех нелимфоидных тканей. Но вдобавок CD69 колокализуется с маркером CD103, который обозначает группу молекул адгезии — интегринов, способствующих прикреплению резидентной Т-клетки к эпителию и к фибробластам в подслизистой выбранного органа. Для эффекторных Т-клеток во вторичных лимфоидных органах экспрессия интегринов CD103 совершенно нехарактерна: TEM-клетки постоянно сохраняют подвижный фенотип.
У карты, составленной коллективом Донны Фарбер, есть крупный недочет: неясно, насколько чисто удается выделить Т-лимфоциты из органа, какую долю анализируемых клеток на самом деле составляют Т-лимфоциты крови из капилляров внутри органа.
Особенно остро вопрос загрязнения клетками крови стоит для легких — неслучайно субпопуляционный состав Т-клеток легких неожиданно похож на Т-клетки крови и лимфоузлов. Вопрос загрязнения клетками крови был изящно решен для Т-лимфоцитов мыши: подопытных животных заражали вирусом лимфоцитарного хориоменингита после пересадки трансгенного клона Т-клеток P14, специфичного к данному вирусу. В результате при инфекции большая часть циркулирующих клеток была представлена вирусоспецифичным клоном P14, а его присутствие в тканях можно было выявить с помощью флуоресцирующих антител к TCR P14. Мышам в кровь вводили антитело анти-CD8 к маркеру Т-киллерных клеток, оно быстро распространялось по кровотоку и связывалось со всеми Т-киллерами в крови (но не в тканях). При микроскопии срезов органов легко было отличить резидентные киллерные TRM от только недавно вышедших из крови в орган клеток, помеченных анти-CD8 антителом [9]. Численность резидентных клеток, подсчитанная этим методом, в 70 раз превышала количество, определенное методом проточной цитометрии; разница меньше чем в два раза наблюдалась только для резидентных клеток лимфоузлов и селезенки. Получается, стандартные методики выделения лимфоцитов из органов плохо подходят для анализа киллерных резидентных клеток и существенно занижают размеры популяции.
Работа резидентных Т-клеток: не стоит путать туризм с эмиграцией
Мышиные резидентные клетки тканей в нормальной ситуации почти не перемещаются внутри нелимфоидной ткани и достаточно прочно прикреплены молекулами адгезии к строме органа. Когда резидентные макрофаги той же ткани секрецией цитокинов инициируют реакцию воспаления, ТRM приобретают большую подвижность и патрулируют близлежащий эпителий в поисках зараженных клеток.
Если воспалительная реакция усиливается, то клетки понимают это как сигнал о подкреплении: к работе патрульных TRM подключаются вновь прибывающие из крови TCM— и TEM-клетки. Эти клетки крови куда более подвижны и лучше перемещаются в эпителии. Значит ли это, что именно в крови находятся готовые действовать Т-киллеры среди TEM, а CD8+ TRM выполняют в ткани хелперные и регуляторные функции?
С одной стороны, Т-хелперы по спектру Т-клеточных рецепторов более тканеспецифичны, т.е. пересечений между репертуарами TCR клеток, взятых из разных тканей, совсем мало, тогда как клетки одного клона Т-киллера встречаются в разных тканях среди TEM [6]. Спектр функций и репертуар антигенной специфичности TRM еще предстоит исследовать, но способности к уничтожению зараженных клеток тканей у TRM-киллеров точно есть. Более того, в модели мышиной инфекции полиомавирусом, протекающей в ткани головного мозга, аффинность вирусоспецифичных Т-клеточных рецепторов резидентных киллерных клеток выше, чем у вирусоспецифичных клеток центральной памяти [10].
Однако размер популяции Т-клеток зависит не только от специфичности TCR к инфекциям, которые раньше протекали в данном органе, но и от гомеостатической пролиферации Т-клеток — размножения более удачливых клеток для заполнения емкости органа по числу Т-лимфоцитов. По маркерам CD28 и CD127 на поверхности клеток можно отличить недавно и давно активированные через TCR клетки от тех, которые получили только гомеостатический сигнал к пролиферации от фактора роста IL-7. При старении ткани гомеостатическое размножение клеток начинает преобладать над пролиферацией активированных через TCR клеток.
Независимо от Т-клеточных рецепторов часто функционируют NKT-клетки, тип резидентных клеток печени, встречающихся и в других тканях. Они могут быть активированы NK-клеточными рецепторами через распознавание не индивидуальных антигенов, а общих молекулярных паттернов опасности и тканевого стресса. При активации CD8+ NKT-клетки выделяют цитотоксические гранулы и лизируют подозрительные клетки ткани, к примеру единичные опухолевые клетки и зараженные вирусами клетки, экспрессирующие и выставляющие на внешней мембране стрессорные молекулы. При старении тенденция TRM к активации без Т-клеточного рецептора, через NK-клеточные рецепторы или цитокиновые сигналы, может приводить к ошибочному лизису клеток ткани, недостаточному контролю над хронически зараженными или перерождающимися участками эпителия.
Патологические проявления, связанные с работой резидентных Т-клеток, включают органоспецифичные аутоиммунные синдромы и синдромы хронического воспаления в ткани. Примеры хронического воспаления, поддерживаемого резидентными Т-лимфоцитами, — контактный дерматит и псориаз, а механизмом служит выделение воспалительных факторов IL-17 резидентными Т-киллерами и IL-22 резидентными Т-хелперами дермы. CD8+ эффекторные Т-киллеры, находящиеся в головном мозге, похожи по совокупности мембранных молекул-маркеров на TRM кожи, кишечника и легких и способны подталкивать развитие перемежающегося рассеянного склероза при периодических выбросах воспалительных цитокинов. Неясно, однако, есть ли в норме в головном мозге популяция TRM или же это Т-лимфоциты, оставшиеся в ткани после нейротропной вирусной инфекции [8].
Функции резидентных клеток памяти в норме — при отсутствии инфекции или хронического воспаления — могут включать cross-talk (взаимную регуляцию преимущественно через секрецию цитокинов и костимуляторные молекулы) с неклассическими малоизученными лимфоидными клетками. Ими могут быть ассоциированные со слизистыми γ/δ Т-клетки, несущие альтернативный вариант сборки Т-клеточного рецептора, или лимфоидные клетки врожденного иммунитета (Innate Lymphoid Cells, ILC), которые делят с Т- и В-лимфоцитами общие черты эпигенетического ландшафта, но не имеют Т-/В- или NK-клеточных рецепторов [11, 12].

TRM клетки контактируют с антигенпрезентирующими клетками тканей — дендритными клетками кожи и резидентными макрофагами тканей. Резидентные миелоидные клетки в разных тканях дифференцированы и слабо похожи друг на друга. К примеру, макрофаги маргинальной зоны селезенки, макрофаги печени и микроглия (макрофаги мозга) будут сильно отличаться и по морфологии, и по спектру функций. Кроме обнаружения антигенов в ткани, резидентные макрофаги заняты регуляцией процессов старения и самообновления тканей, в частности, выделяют факторы роста и цитокины, стимулирующие деление стволовых клеток тканей. В жировой ткани, к примеру, макрофаги стимулируют дифференцировку новых жировых клеток, но при переходе в активированное M1-состояние запускают воспаление и вместо дифференцировки заставляют увеличиваться и набухать имеющиеся жировые клетки. Сопутствующие изменения метаболизма жировой ткани приводят к накоплению жировой массы и в последние годы связываются с механизмами развития ожирения и диабета II типа. В коже цитокины, выделяемые макрофагами и резидентными γ/δ Т-клетками, стимулируют деление стволовых клеток при регенерации эпидермиса и стволовых клеток волосяных фолликулов [13, 14]. Можно предположить, что хелперные TRM-клетки при патрулировании эпителия и образовании контактов с тканевыми макрофагами могут модулировать спектр и объем выделяемых последними факторов роста для стволовых клеток, воспалительных цитокинов и факторов ремоделирования эпителия — и тем самым участвовать в обновлении тканей.
Что изучение TRM может дать медицине?
Понимание принципов работы резидентных Т-клеток абсолютно необходимо для борьбы с инфекциями, которые не поступают сразу в кровь, а проникают в организм через барьерные ткани, т.е. для подавляющего большинства инфекций. Рациональный дизайн вакцин для защиты от этой группы инфекций может быть направлен именно на усиление первого этапа защиты с помощью резидентных клеток. Ситуация, при которой оптимально активированные специфичные к антигену клетки элиминируют патоген в барьерной ткани, куда выгоднее, чем запуск острого воспаления для вызова Т-лимфоцитов из крови, поскольку меньше повреждается ткань.
Репертуар TCR, ассоциированных со слизистыми барьерных тканей, считается частично вырожденным и наиболее распространенным, т.е. идентичным для многих людей в популяции. Тем не менее искажения при выделении Т-клеток из органов, перекос данных в результате отбора в когорты только определенных европеоидных доноров и общее небольшое количество накопленных данных секвенирования не дают уверенности в публичности репертуаров Т-клеточных рецепторов TRM-клеток. Впрочем, это было бы удобно: дизайн вакцин мог бы сводиться к поиску и модификации наиболее аффинных и иммуногенных пептидов в патогене, взаимодействующих с одним из распространенных вариантов ТCR в барьерной для этого патогена ткани.
Конечно, представления о том, какие TCR несут на своей поверхности TRM-клетки, недостаточно для того, чтобы эффективно манипулировать иммунными реакциями в ткани. Предстоит детально изучить факторы, влияющие на заселение тканей определенными клонами Т-клеток, и разобраться в механизмах активации местного тканевого иммунитета и индукции толерантности TRM. Как заселяются ниши Т-лимфоцитов в слизистых у ребенка до встречи с большим числом патогенов и, соответственно, до формирования значительного пула эффекторных Т-клеток памяти — предшественников резидентных клеток и клеток центральной памяти? Почему и как вместо классической активации лимфоцитов формируется реакция толерантности к микробам непатогенной флоры слизистых? Эти вопросы стоят на повестке дня в изучении резидентных клеток иммунной системы.
Определение закономерностей хоминга Т-лимфоцитов в определенные ткани может дать преимущество в клеточной иммунотерапии опухолевых заболеваний. Теоретически киллерные Т-клетки нужной специфичности к опухолевому антигену, активированные in vitro, должны убивать опухолевые клетки пациента. На практике подобная иммунотерапия осложняется тем, что опухолевые клетки способны подавлять иммунные реакции и приводить в неактивное состояние приближающиеся к опухоли Т-киллеры. Зачастую в массе растущей опухоли и вокруг нее накапливаются анергичные Т-лимфоциты — в первую очередь TRM данной ткани. Из множества инъецированных пациенту активных опухолеспецифичных Т-клеток до цели дойдут немногие, и даже они могут оказаться практически бесполезными в иммуносупрессивном микроокружении опухоли.
Расшифровка механизмов, которые обеспечивают попадание конкретных клонов Т-клеток в определенные ткани, может позволить более эффективно направлять к опухоли сконструированные в лаборатории Т-лимфоциты и приблизить эру доступной персонализированной иммунотерапии.
Литература
1. Clark R. A. Skin resident T cells: the ups and downs of on site immunity // J. Invest. Dermatol. 2010. V. 130. P. 362–370. doi: 10.1038/jid.2009.247
2. Doherty D. G., Norris S., Madrigal-Estebas L. et al. The human liver contains multiple populations of NK cells, T cells, and CD3+CD56+ natural T cells with distinct cytotoxic activities and Th1, Th2, and Th0 cytokine secretion patterns // J. Immunol. 1999. V. 163. P. 2314–2321.
3. Iijima N., Iwasaki A. Tissue instruction for migration and retention of TRM cells // Trends Immunol. 2015. V. 36. P. 556–564. doi: 10.1016/j.it.2015.07.002
4. Farber D., Yudanin N., Restifo N. P. Human memory T cells: generation, compartmentalization and homeostasis // Nat. Rev. Immunol. 2014. V. 14. P. 24–35. doi: 10.1038/nri3567
5. Schenkel J. M., Masopust D. Tissue-resident memory T cells // Immunity. 2014. V. 41. P. 885–897. doi: 10.1016/j.immuni.2014.12.007
6. Thome J. J., Yudanin N., Ohmura Y. et al. Spatial map of human T cell compartmentalization and maintenance over decades of life // Cell. 2014. V. 159. P. 814–828. doi: 10.1016/j.cell.2014.10.026
7. Britanova O. V., Putintseva E. V., Shugay M. et al. Age-related decrease in TCR repertoire diversity measured with deep and normalized sequence profiling // J. Immunol. 2014. V. 192. P. 2689–2698. doi: 10.4049/jimmunol.1302064
8. Park C. O., Kupper T. S. The emerging role of resident memory T cells in protective immunity and inflammatory disease // Nat. Med. 2015. V. 21. P. 688–697. doi: 10.1038/nm.3883
9. Steinert E. M., Schenkel J. M., Fraser K. A. et al. Quantifying memory CD8 T cells reveals regionalization of immunosurveillance // Cell. 2015. V. 161. P. 737–749. doi: 10.1016/j.cell.2015.03.031
10. Frost E. L., Kersh A. E., Evavold B. D., Lukacher A. E. Cutting edge: resident memory CD8 T cells express high-affinity TCRs // J. Immunol. 2015. V. 195. P. 3520–3524. doi: 10.4049/jimmunol.1501521
11. Diverse functions of mucosal resident memory T cells / Eds K. S. Schluns, K. Klonowski // Frontiers in Immunology [Electronic resource]. N.Y., 2015. doi: 10.3389/fimmu.2015.00090
12. Godfrey D. I., Uldrich A. P., McCluskey J. et al. The burgeoning family of unconventional T cells // Nat. Immunol. 2015. V. 16. P. 1114–1123. doi: 10.1038/ni.3298
13. Castellana D., Paus R., Perez-Moreno M. Macrophages contribute to the cyclic activation of adult hair follicle stem cells // PLoS Biol. [Electronic resource]. 2014. V. 12. e1002002. doi: 10.1371/journal.pbio.1002002
14. Rodero M. P., Khosrotehrani K. Skin wound healing modulation by macrophages // Int. J. Clin. Exp. Pathol. 2010. V. 3. № 7. P. 643–653.
К
клеткам иммунной системы относят
лимфоциты,
макрофаги и другие антиген- представляющие
клетки (А-
клетки, от англ. accessory-
вспомогательный), а также так называемую
третью
популяцию клеток (т.е.
клеток, не имеющих основных поверхностных
маркеров Т- и В- лимфоцитов, А- клеток).
По
функциональным свойствам все
иммунокомпетентные клетки разделяют
на эффекторные
и регуляторные.
Взаимодействие клеток в иммунном ответе
осуществляется с помощью гуморальных
медиаторов — цитокинов.
Основные клетки иммунной системы- Т- и
В- лимфоциты.
Лимфоциты.
В
организме лимфоциты постоянно
рециркулируют между зонами скопления
лимфоидной ткани. Расположение лимфоцитов
в лимфоидных органах и их миграция по
кровеносному и лимфатическому руслу
строго упорядочены и связаны с функциями
различных субпопуляций.
Лимфоциты
имеют общую морфологическую характеристику,
однако их функции, поверхностные CD
(от claster
differenciation)
маркеры, индивидуальное (клональное)
происхождение, различны.
По
наличию поверхностных CD
маркеров лимфоциты разделяют на
функционально различные популяции и
субпопуляции, прежде всего на Т-
(тимусзависимые,
прошедшие первичную дифференцировку
в тимусе) лимфоциты и В
— (bursa-
зависимые, прошедшие созревание в сумке
Фабрициуса у птиц или его аналогах у
млекопитающих) лимфоциты.
Т-
лимфоциты.
Локализация.
Обычно
локализуются в так называемых Т- зависимых
зонах периферических лимфоидных органов
(периартикулярно в белой пульпе селезенки
и паракортикальных зонах лимфоузлов).
Функции.
Т-
лимфоциты распознают процессированный
и представленный на поверхности антиген-
представляющих ( А ) клеток антиген. Они
отвечают за клеточный
иммунитет,
иммунные реакции клеточного типа.
Отдельные субпопуляции помогают В-
лимфоцитам реагировать на Т-
зависимые антигены
выработкой антител.
Происхождение
и созревание.
Родоначальницей
всех клеток крови, в том числе лимфоцитов,
является единая
стволовая клетка костного мозга.
Она генерирует два типа клеток-
предшественников- лимфоидную стволовую
клетку и предшественника клеток красной
крови, от которой происходят и клетки-
предшественники лейкоцитов и макрофагов.
Образование
и созревание иммунокомпетентных клеток
осуществляется в центральных органах
иммунитета (для Т- лимфоцитов- в тимусе).
Клетки- предшественники Т- лимфоцитов
попадают в тимус, где пре- Т- клетки
(тимоциты) созревают, пролиферируют и
проходят дифференцировку на отдельные
субклассы в результате взаимодействия
с эпителиальными и дендритными клетками
стромы и воздействия гормоноподобных
полипептидных факторов, секретируемых
эпителиальными клетками тимуса ( альфа1-
тимозин, тимопоэтин, тимулин и др.).
При
дифференцировке Т- лимфоциты приобретают
определенный
набор мембранных CD—
маркеров.
Т-клетки разделяют на субпопуляции в
соответствии с их функцией и профилем
CD-
маркеров.
Т-
лимфоциты распознают антигены с помощью
двух типов мембранных гликопротеинов-
Т-
клеточных рецепторов
(семейство Ig-
подобных молекул) и CD3,
нековалентно связанных между собой. Их
рецепторы, в отличие от антител и
рецепторов В- лимфоцитов, не распознают
свободно циркулирующие антигены. Они
распознают пептидные фрагменты,
представляемые им А- клетками через
комплекс чужеродных веществ с
соответствующим белком главной системы
гистосовместимости 1 и 2 класса.
Выделяют
три основные группы Т- лимфоцитов-
помощники
(активаторы), эффекторы, регуляторы.
Первая
группа- помощники (активаторы),
в состав которых входят Т-
хелперы1, Т- хелперы2, индукторы Т-
хелперов, индукторы Т- супрессоров.
1.
Т- хелперы1 несут
рецепторы CD4
(как и Т- хелперы2) и CD44,
отвечают за созревание Т-
цитотоксических лимфоцитов (Т- киллеров),
активируют
Т- хелперы2 и цитотоксическую функцию
макрофагов, секретируют ИЛ-2, ИЛ-3 и другие
цитокины.
2.
Т- хелперы2 имеют
общий для хелперов CD4
и специфический CD28
рецепторы, обеспечивают пролиферацию
и дифференцировку В- лимфоцитов в
антителпродуцирующие (плазматические)
клетки, синтез антител, тормозят функцию
Т- хелперов1, секретируют ИЛ-4, ИЛ-5 и ИЛ-6.
3.
Индукторы Т- хелперов
несут CD29,
отвечают за экспрессию антигенов HLA
класса 2 на макрофагах и других А- клетках.
4.
Индукторы Т- супрессоров
несут CD45
специфический рецептор, отвечают за
секрецию ИЛ-1 макрофагами, активацию
дифференцировки предшественников Т-
супрессоров.
Вторая
группа- Т- эффекторы.
В нее входит только одна субпопуляция.
5.
Т-
цитотоксические лимфоциты (Т- киллеры).
Имеют
специфический рецептор CD8,
лизируют клетки- мишени, несущие
чужеродные антигены или измененные
аутоантигены (трансплантант, опухоль,
вирус и др.). ЦТЛ распознают чужеродный
эпитоп вирусного или опухолевого
антигена в комплексе с молекулой класса
1 HLA
в плазматической мембране клетки-
мишени.
Третья
группа- Т-клетки- регуляторы.
Представлена двумя основными
субпопуляциями.
6.
Т-
супрессоры
имеют важное значение в регуляции
иммунитета, обеспечивая подавление
функций Т- хелперов 1 и 2, В- лимфоцитов.
Имеют рецепторы CD11,
CD8.
Группа функционально разнородна. Их
активация происходит в результате
непосредственной стимуляции антигеном
без существенного участия главной
системы гистосовместимости.
7.
Т-
контсупрессоры.
Не имеют CD4,
CD8,
имеют рецептор к особому лейкину.
Способствуют подавлению функций Т-
супрессоров, вырабатывают резистентность
Т- хелперов к эффекту Т- супрессоров.
В-
лимфоциты.
Существует
несколько подтипов В- лимфоцитов.
Основная функция В- клеток- эффекторное
участие в гуморальных иммунных реакциях,
дифференциация в результате антигенной
стимуляции в плазматические клетки,
продуцирующие антитела.
Образование
В- клеток у плода происходит в печени,
в дальнейшем- в костном мозге. Процесс
созревания В- клеток осуществляется в
две стадии- антиген
— независимую и антиген — зависимую.
Антиген
-независимая фаза. В-
лимфоцит в процессе созревания проходит
стадию пре-
В- лимфоцита- активно
пролиферирующей клетки, имеющей
цитоплазменные H-
цепи типа C
мю (т.е. IgM).
Следующая стадия- незрелый
В- лимфоцит
характеризуется появлением мембранного
(рецепторного) IgM
на поверхности. Конечная стадия
антигеннезависимой дифференцировки-
образование зрелого
В- лимфоцита,
который может иметь два мембранных
рецептора с одинаковой антигенной
специфичностью (изотипа) — IgM
и IgD.
Зрелые В- лимфоциты покидают костный
мозг и заселяют селезенку, лимфоузлы и
другие скопления лимфоидной ткани, где
их развитие задерживается до встречи
со “своим” антигеном, т.е. до осуществления
антиген- зависимой дифференцировки.
Антиген-
зависимая дифференцировка
включает активацию, пролиферацию и
дифференцировку В- клеток в плазматические
клетки и В- клетки памяти. Активация
осуществляется различными путями, что
зависит от свойств антигенов и участия
других клеток ( макрофагов, Т- хелперов).
Большинство антигенов, индуцирующих
синтез антител, для индукции иммунного
ответа требуют участия Т- клеток- тимус-
зависимые пнтигены.
Тимус-
независимые антигены (ЛПС,
высокомолекулярные синтетические
полимеры) способны стимулировать синтез
антител без помощи Т- лимфоцитов.
В-
лимфоцит с помощью своих иммуноглобулиновых
рецепторов распознает и связывает
антиген. Одновременно с В- клеткой
антиген по представлению макрофага
распознается Т- хелпером (Т- хелпером
2), который активируется и начинает
синтезировать факторы роста и
дифференцировки. Активированный этими
факторами В- лимфоцит претерпевает ряд
делений и одновременно дифференцируется
в плазматические клетки, продуцирующие
антитела.
Пути
активации В- клеток и кооперации клеток
в иммунном ответе на различные антигены
и с участием популяций имеющих и не
имеющих антиген Lyb5
популяций В- клеток отличаются. Активация
В- лимфоцитов может осуществляться:
—
Т- зависимым антигеном при участии
белков МНС класса 2 Т- хелпера;
—
Т- независимым антигеном, имеющим в
составе митогенные компоненты;
—
поликлональным активатором (ЛПС);
—
анти- мю иммуноглобулинами;
—
Т- независимым антигеном, не имеющим
митогенного компонента.
Кооперация
клеток в иммунном ответе.
В
формировании иммунного ответа включаются
все звенья иммунной системы- системы
макрофагов, Т- и В- лимфоцитов, комплемента,
интерферонов и главная система
гистосовместимости.
В
кратком виде можно выделить следующие
этапы.
1.
Поглощение и процессинг антигена
макрофагом.
2.
Представление процессированного
антигена макрофагом с помощью белка
главной системы гистосовместимости
класса 2 Т- хелперам.
3.
Узнавание антигена Т- хелперами и их
активация.
4.
Узнавание антигена и активация В-
лимфоцитов.
5.
Дифференциация В- лимфоцитов в
плазматические клетки, синтез антител.
6.
Взаимодействие антител с антигеном,
активация систем комплемента и макрофагов,
интерферонов.
7.
Представление при участии белков МНС
класса 1 чужеродных антигенов Т- киллерам,
разрушение инфицированных чужеродными
антигенами клеток Т- киллерами.
8.
Индукция Т- и В- клеток иммунной памяти,
способных специфически распознавать
антиген и участвовать во вторичном
иммунном ответе ( антигенстимулированные
лимфоциты).
Клетки
иммунной памяти.
Поддержание долгоживущих и метаболически
малоактивных клеток памяти, рециркулирующих
в организме, является основой длительного
сохранения приобретенного иммунитета.
Состояние иммунной памяти обусловлено
не только длительностью жизни Т- и В-
клеток памяти, но и их антигенной
стимуляцией. Длительное сохранение
антигенов в организме обеспечивается
дендритными клетками (депо антигенов),
сохраняющими их на своей поверхности.
Соседние файлы в папке 2
- #
- #
- #
- #
- #