Содержание
- Жизнь в гидротермальных источниках
- Типы гидротермальных источников
- Где они находятся?
- Значение
Несмотря на свой негостеприимный вид, гидротермальные источники, также известные как “черные курильщики” и “белые курильщики”, поддерживают жизнь многих организмов, включая экстремофилов. Они представляют собой по сути, подводные гейзеры, созданные тектоническими плитами. Эти огромные плиты в земной коре движутся и создают трещины на дне океана.
Океаническая вода попадает в трещины, нагревается земной магмой, а затем выделяется через гидротермальные отверстия. Вода, выходящая из гидротермальных источников, может достигать невероятных температур до 400°С, даже если вода за пределами этих источников имеет температуру, близкую к замерзанию. Несмотря на то что вода очень горячая, она не кипит, потому что находится под высоким давлением.
Из-за отдаленного глубоководного местоположения, гидротермальные источники были открыты сравнительно недавно. Только в 1977 году во время экспедиции на подводном аппарате Алвин были зафиксированы “черные курильщики”, извергающие горячую воду и минералы в холодные воды Тихого океана, вблизи Галапагосских островов. Еще более удивительно было обнаружить эти негостеприимные области, изобилующие морскими существами.
Жизнь в гидротермальных источниках

Жизнь традиционно воспринимается как энергия солнца, но глубоководные организмы не имеют доступа к солнечному свету, поэтому они зависят от питательных веществ гидротермальных источников. По сравнению с окружающим морским дном, гидротермальные источники имеют плотность организмов в 10 000-100 000 раз больше.
Эти источники способны поддерживать такое огромное количество жизни, потому что их обитатели зависят от хемосинтезирующих бактерий, как источника питания. Гидротермальная вода богата растворенными минералами и поддерживает большую популяцию хемотрофов. Эти бактерии используют соединения серы, особенно сероводород, который высокотоксичный для большинства известных организмов, чтобы получать органический материал в процессе хемосинтеза.
Созданная экосистема отличается от большинства наземных экосистем Земли, которые основаны на солнечной энергии. Однако, хотя часто говорят, что эти сообщества существуют независимо от солнца, некоторые из организмов на самом деле зависят от кислорода, создаваемого фотосинтезирующими организмами, тогда как другие являются анаэробными.
Хемосинтезирующие бактерии превращаются в толстый коврик, который привлекает другие организмы, такие как амфиподы и копеподы. Крупные организмы, такие как улитки, креветки, крабы, трубчатые черви, рыба (особенно угорь, слитожаберный угорь, ошибнеобразные и др.) и осьминоги (особенно Vulcanoctopus hydrothermalis), образуют звенья пищевой цепочки гидротермальных источников.
Типы гидротермальных источников
Как правило, выделяют два типа гидротермальных источников – «черные курильщики» и «белые курильщики».
Самые горячие – «черные курильщики», получили свое название, потому что они извергают темную воду, состоящую в основном из железа и сульфида. Эта комбинация образует моносульфид железа, который придает воде черный цвет.
«Белые курильщики» выпускают более холодную, легкую воду, состоящую из соединений, включая барий, кальций и кремний.
Где они находятся?
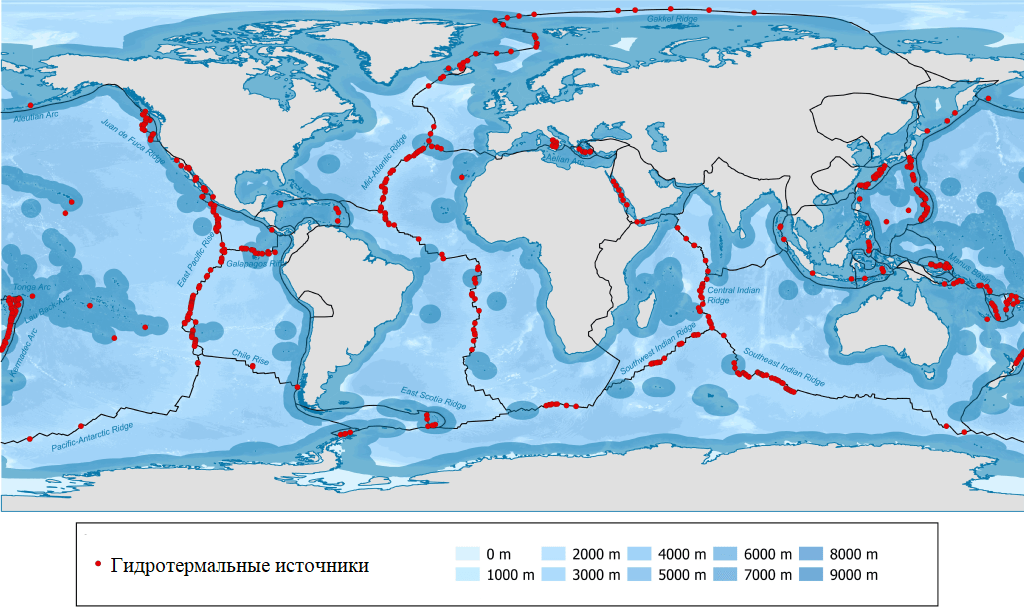
Гидротермальные источники, как правило, распределены вдоль границ тектонических плит Земли, хотя они также могут быть обнаружены внутри плит, в горячих вулканических точках.
Значение
Гидротермальные источники играют важную роль в циркуляции океанических течений и регулируют химический состав океанических вод. Они являются источниками питательных веществ, требуемых для организмов. Микроорганизмы, обнаруженные в гидротермальных источниках, также могут иметь важное значение для разработки лекарственных средств и других продуктов. Добыча полезных ископаемых, обнаруженных в гидротермальных источниках, является новой проблемой, которая может позволить ученым больше узнать об этих негостеприимных районах, но также способна повредить морское дно и окружающие морские экосистемы.

Гугломаг
Спрашивай! Не стесняйся!
Задать вопрос
Не все нашли? Используйте поиск по сайту
Укажите номера предложений, в которых верно передана ГЛАВНАЯ информация, содержащаяся в тексте. Запишите номера этих предложений.
1) Многие районы Мирового океана — от прибрежной полосы до участков, расположенных на больших глубинах, — обладают большими запасами разнообразных полезных ископаемых, однако современные возможности позволяют использовать пока далеко не все эти ископаемые.
2)Современные технологии позволили освоить пока только энергию приливов, хотя часть механической и тепловой энергии Мирового океана образует возобновляемые и другие энергетические ресурсы, которые могут быть использованы человеком.
3) Растворённые в океанской и морской воде органические и неорганические элементы образуют возобновляемые энергетические ресурсы, которые могут быть использованы человеком и способны обеспечить потребности человечества на многие сотни лет.
4) Часть механической и тепловой энергии Мирового океана образует возобновляемые энергетические ресурсы, которые могут быть использованы человеком, однако современные технологии позволили освоить пока только энергию приливов.
5) Часть механической и тепловой энергии Мирового океана (морских течений, волн, прилива) образует океанические и морские возобновляемые энергетические ресурсы, принципиально доступные для производственной деятельности людей.
Установите соответствие между экологическими особенностями и группами организмов: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
А) служат начальным звеном пастбищных цепей питания
Б) определяют пространственную структуру в наземных биогеоценозах
В) могут быть редуцентами
Г) образуют споры для перенесения неблагоприятных условий
Д) обитают в океанах на глубине более километра
Е) формируют первый трофический уровень
ГРУППЫ ОРГАНИЗМОВ
1) гетеротрофные бактерии
2) высшие растения
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| А | Б | В | Г | Д | Е |
1. Глубоководные экосистемы (экватор и южные широты)
Выполнила:
Айдарова В.В.
гр. Эпб-412
2. Содержание
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
Абиссальная зона
Характеристика глубоководных зон океана
Особенности живых организмов абиссальной зоны
Питание в глубоководной зоне
Обитатели абиссальной зоны
Рифтовые зоны
Черные курильщики
Хемоавтотрофные экосистемы рифтовых зон
Особенности хемоавтотрофных экосистем
Геотермальные «поля»
Обитатели хемоавтотрофных экосистем
3. Абиссальная зона
Абиссаль – зона расположена в глубинах ложа Мирового
океана, с глубиной более 2000 метров. Это наиболее
глубоководная зона мирового океана.
4. Характеристика глубоководных зон океана
Глубина более 2000 м;
Плотность биомассы от 0,01 кг/м до 15 кг/м;
Полная темнота;
Огромное давление;
Высокое содержание сероводорода и ядовитых
металлов;
Имеются выходы термальных источников;
Пониженная температура воды;
Недостаток пищевых ресурсов;
5.
В данной экосистеме серные бактерии играют роль
растений.
Холодные воды (от 0 до 5 по Цельсию);
Соленость составляет 35‰;
Дно покрыто известняковыми и кремнистыми илами;
Накопление слоя в глубоководных участках идет очень
медленно, за одно тысячелетие это слой вырастает
всего на один сантиметр;
Из царства растений в абиссальной зоне встречаются
только бактерии и несколько видов плесневых грибов.
6. Особенности живых организмов абиссальной зоны
Животные на больших глубинах бесцветны или слабо
окрашены в дополнительный розовый, красный цвет;
Развиты органы осязания;
У многих животных развились очень тонкие органы
чувств, которые позволяют им обнаружить присутствие
другого животного по легкому изменению давления,
вызванному движущимся телом.
7. Адаптация живых организмов
Органы свечения;
Редукция плавательного пузыря у рыб или заполнение
его полости жировой тканью;
Атрофированы органов зрения;
Отлично развиты осязательные органы, которые,
словно антенны, торчат в разные стороны.
8. Питание в глубоководной зоне
Пищевая цепь:
1. Серобактерии;
2. Погонофоры, внутри тела которых обитают
бактерии, перерабатывающие сероводород
(гигантские черви, крупными двустворчатыми
моллюсками, отдельными видами рыб и т.д.) ;
3. Детритофаги.
Пищей для обитателей (помимо хищников)
глубоководной зоны служат непрерывно
опускающиеся остатки организмов и растений из
пелагической зоны.
9. Обитатели абиссальной зоны
Погонофоры —беспозвоночные животных, обитающих в хитиновых
трубках;
Ксенофиофоры — эти простейшие барофильные бактерии;
Фораминиферы — отряд простейших подкласса корненожек с
цитоплазматическим телом, одетым раковиной;
Двустворчатые и брюхоногие моллюски;
Равноногие раки;
Многощетинковые черви;
Голотурии;
Бокоплавы;
Рыбы;
10. Stomias boa
Stomias boa (Scaly
dragonfish) глубоководная рыба
семейства Stomiidae;
Максимальный
размер достигает
32 см.
11. Ласиогнаты
Ласиогнаты (лат.
Lasiognathus) — род
глубоководных
лучепёрых рыб
семейства
тауматихтовых.
Ласиогнаты
обитают на
глубине 4
километра.
12. Footballfish
Встречается в тропических
и субтропических водах
Атлантического,
Индийскогои, Тихого
океана.
Был обнаружен на
максимальной глубине 4
км, самки вырастают в
длину до 60 см, самцы до 4
см;
13. Скорпеноподобная морская рыба
Paraliparis copei copei
(Blacksnout seasnail) скорпеноподобная
морская рыба
семейства Liparidae;
Обитает во всех
мировых океанах.
14. Mertensia ovum — морское животное типа гребневиков
Имеют яркую
пигментацию.
Гребневики широко
распространены по всему
миру и найдены во всех
частях Мирового океана.
15. Рифтовые зоны (места разломов плит литосферы)
В абиссальной зоне существуют рифтовые зоны в
которых в 70 х гг. ХХ века были открыты
хемоавтотрофные экосистемы, получившие название
глубоководных геотермальных «оазисов».
В рифтовых зонах из расщелин горной породы
выделяются горячие воды, насыщенные
сероводородом, сульфидами железа, цинка, меди и
других тяжелых металлов.
Температура «гейзеров» достигает 300°C.
16. Черные курильщики
17.
18. Хемоавтотрофные экосистемы рифтовых зон
У оснований «черных курильщиков» и формируется «оазис»,
являющиеся хемоавтотрофными экосистемами.
Продуцентами этих экосистем являются серобактерии,
образующие скопления – бактериальные маты.
Вестиментиферы – представители типа погонофор (черви
длиной 1–2,2 м, заключенные в длинные белые трубки из
хитиноподобного вещества).
В этих экосистемах, кроме того, много видов животных
хищников (крабы, моллюски, некоторые глубоководные
рыбы)
19. Особенности хемоавтотрофных экосистем
Очень высокая продуктивность
Экосистемы «оазисов» существуют недолго и
разрушаются после того, как прекратится деятельность
подводных гейзеров.
Кроме «оазисов» существуют еще и геотермальные
«поля».
20. Геотермальные «поля»
Обнаружены вдоль Центрального Атлантического хребта,
простирающегося от Исландии до экватора.
Глубина до 800 м.
Они охватывают непосредственно хребет и окружающие его приподнятые
участки дна, ширина «полей» может достигать 75 км.
Температура вод, поднимающихся из расщелин – от 50 до 300оС.
Жизнь экосистем «полей», в отличие от «оазисов», представлена только
бактериями.
На сегодняшний день исследовано более 40 «полей».
21. Обитатели хемоавтотрофных экосистем
СИДЯЧИЕ
ЧЕРВЕОБРАЗНЫЕ
ОРМЫ –
ПОГОНОФОРЫ (ТИП
POGONOPHORA)
ХАРАКТЕРНЫЙ
ПРЕДСТАВИТЕЛЬ
ПОГОНОФОР,
ОТНОСЯЩИЙСЯ К
КЛАССУ
VESTIMENTIFERA
Распространены во
всех морях.
22. Calyptogena magnifica
Живут на глубинах от 200 до
1500 м.
Длина раковин может
достигать 25 см.
Обитает в Тихом океане,
экваториальные зоны
Атлантического океана.
23. Червь-комменсал батимодиолы Branchipolynoe symmytilida.
Червькомменсал
батимодиолы
Branchipolynoe
symmytilida.
24. Рыба из семейства бельдюговых
Охожая на толстого
угря рыба длиной не
более 30 см.
Обитает в тихом
океане.
25. СПАСИБО ЗА ВНИМАНИЕ!!!

На основе ландшафтно-экологического подхода – выделения руководящих видов и их комплексов, проведено биогеографическое районирование гидротермали и описаны особенности ее вертикального распределения. В гидротермали Мирового океана выделено пять основных географических биоценотических комплексов, восемь биоценотических регионов и три вертикальных биоценотических комплекса, границы которых в основном совпадают с аналогичными границами обычной, не гидротермальной фауны.
В глубинах океанов, куда уже не проникает дневной свет, а органика, достигающая дна, весьма скудна (и многократно переварена обитателями верхних слоев воды – от животных до бактерий), где и быть ничего не должно, кроме зазеркальной Сахары, — в массе проживают организмы, многим из которых нет аналогов в нашем обычном, солнечном, мире. Эта жизнь не распределена равномерно, а сгруппирована по большей части на гидротермальных полях, в океанических рифтах – границах между раздвигающимися литосферными плитами. Здесь происходит выделение флюидов – морской воды, проникшей под поверхность земной коры на несколько сотен метров, обогатившейся там солями металлов, сероводородом, метаном и другими углеводородами, и в новом качестве возвращающейся в океан, в виде гейзеров с температурой свыше 350°С – черных и белых курильщиков. В этих районах, малопригодных для обычных живых существ, создаются благоприятные условия для развития хемосинтезирующих микроорганизмов – взвешенных в толще воды и образующих бактериальные маты на склонах курильщиков – и животных, этими организмами питающихся или вступивших с ними в симбиотические отношения. Хемоавтотрофные и метанотрофные микроорганизмы, способные синтезировать органическое вещество, используя углекислоту и/или метан в качестве источника углерода, а восстановленные минеральные соединения (H2, H2S, S, S2O32-, SO2, NH3, Fe2+, Mn2+ и др.) – в качестве источника энергии, могут прокормить многие виды животных. Гидротермальные экосистемы почти целиком зависят от геотермальной, а не солнечной энергии.
На уступах курильщиков, подальше от устья, но в зоне, куда из курильщика в изобилии выбрасываются мелкие частицы серы, при температуре 30-40°С проживают 20-и сантиметровые трубчатые помпейские черви (Alvinella pompejana), которые относятся к классу многощетинковых червей – полихет. Помпейские черви получили свое видовое название в память о засыпанных горячим вулканическим пеплом Помпеях, а родовое имя унаследовали от обитаемого подводного аппарата «Алвин», из которого исследователи наблюдали гидротемальную жизнь впервые в 1977 г. Вероятно, помпейские черви питаются как за счет всасывания низкомолекулярных веществ, выделяемых бактериями-симбионтами, живущими на поверхности их тела, так и за счет фильтрации воды.

В Атлантическом океане в этой зоне пасутся стада креветок Rimicaris. Эти креветки выращивают бактерий-симбионтов на жабрах.
Следующим концентрическим рядом вокруг гидротермального источника (вода здесь еще теплая и дрожит «муаром», как воздух над раскаленным асфальтом) располагаются вестиментиферы, родственники погонофор – крупные организмы, длиной от 5-7 см до 2,5 м. Их тело тоже заключено в трубку, прикрепленную ко дну. У взрослых вестиментифер нет ни рта, ни кишечника, зато есть специальный орган – трофосома (остаток кишечника), приютившая симбиотических сероводородокисляющих бактерий. Бактерии-симбионты получают от вестиментифер сероводород и кислород и делятся в свою очередь с хозяином органическими веществами, которые служат единственным источником питания вестиментифер. Исследования показали, что личинки вестиментифер не имеют симбиотических бактерий, зато имеют нормально развитый рот и кишечник. До недавнего времени считалось, что личинки приобретают «своих» бактерий, заглатывая их из окружающей среды, но недавно было показано, что симбиотические бактерии проникают в молодых осевших на дно вестиментифер через наружные покровы тела, т.е. процесс напоминает заражение патогенными микроорганизмами. Видовой состав вестиментифер варьирует в зависимости от региона. Также в разных регионах с ними соседствуют двустворчатые или брюхоногие моллюски, либо многощетинковые черви. Эти животные тоже отлично приспособлены к жизни на специфическом поле гидротерм. Так, например, у двустворчатых моллюсков рода Calyptogena кишечник короткий и ротовые лопасти не функционируют, но клетках их жаберного эпителия обнаружены симбиотические бактерии (см.: Прочтен геном симбиотической бактерии из клеток гидротермального моллюска. «Элементы», 22.02.07). У двустворок рода Bathymodiolus в жабрах также обитают бактерии, однако пищеварительная система этого моллюска развита нормально. Bathymodiolus могут питаться и за счет симбионтов-бактерий, и с помощью обычной для двустворок фильтрации.

В Атлантике зону воздействия теплого флюида делят креветки Rimicaris и двустворчатые моллюски Bathymodiolus.
Именно этой зоной ограничено распространение наиболее специализированных гидротермальных форм. Вне прямого влияния флюида наблюдается концентрация фоновой фауны: десятиногих и усоногих раков, осьминогов, актиний и рыб, которые питаются обитателями гидротерм, но не привязаны к самим гидротермам жестко.
Изучение гидротермальных сукцессий затруднено малой продолжительностью существования самих гидротерм, сравнимой с продолжительностью прохождения сукцессий: в некоторых источниках концентрация Н2S в горячем растворе за 12 лет уменьшается в 86 раз! Предположительно, возникающие гидротермы колонизируются сначала помпейскими червями, которых частично сменяют вестиментиферы. К ним постепенно присоединяются двустворчатые моллюски или гастроподы. Далее увеличивается количество и разнообразие десятиногих ракообразных. Однако в любой момент колебания активности гидротермы способны вернуть сукцессию на любую из уже пройденных стадий.
В общем, на гидротермальных полях наблюдается низкое видовое разнообразие при высокой численности.
Окаменелости, похожие на трубки вестиментифер и гидротермальных двустворок, найдены в залежах руд, образовавшихся около 350 млн лет назад. В гидротермалях обитают архаичные животные (например, усоногие ракообразные рода Neolepas) – малоизменившиеся потомки видов, существовавших более 230-130 миллионов лет назад. Среди нескольких сот видов, известных из гидротермальных оазисов, более 88% водятся только в этих биотопах и выработали многочисленных приспособления, связанные с проживанием в специфических условиях. Все это свидетельствуют о древнем происхождении обитателей гидротерм. Однако незначительное изменение генома некоторых симбиотических бактерий говорит о том, что симбиотическая система находится на раннем этапе становления. На сегодняшний день ученые пришли к выводу о том, что большинство современных гидротермальных видов имеют молодой (кайнозойский) возраст.
Самобытность гидротермальных оазисов – способ добычи пищи и энергии, объемы хемосинтетической продукции, в десятки тысяч раз превышающие таковые на соседних участках морского дна, само видовое своеобразие – породили представление об уникальности, изолированности и энергетической независимости гидротермальных систем от солнечной энергии. Гидротермы – как зарождение жизни на земле. Гидротермы – иная, альтернативная жизнь… По словам Г.М. Виноградова, считалось, будто «мирки гидротерм плотно замкнуты сами на себя… их обитатели когда-то давно пришли из внешнего мира – и закрыли за собой дверь».
Но, во-первых, всем гидротермальным животным, а также аэробным хемосинтезирующим и метанокисляющим бактериям, требуется кислород, представляющий собой продукт фотосинтеза.
Во-вторых, даже самые специализированные организмы становятся «независимыми» только с какого-то момента своего существования: почти все обитатели гидротермальных полей имеют планктонных личинок, живущих в толще воды и потребляющих «обычные» фотосинтетические продукты питания. Даже молодь специализированных креветок Rimicaris получает за счет симбиотических бактерий только 30% углерода.
В-третьих, некоторые весьма подвижные организмы уходят далеко от самих источников, разнося произведенное там органическое вещество на значительные расстояния (правда, КПД использования бактериальной продукции чрезвычайно высок, и основная часть создаваемой органики используется внутри самих гидротерм).
Таким образом, представления об энергетической независимости гидротермальных экосистем, в том числе и в прошлые геологические эпохи, несколько преувеличены.
В статье С. В. Галкина из Института океанологии им. П.П. Ширшова РАН гидротермаль рассматривается в контексте общего существования глубоководной фауны. В работе представлены результаты исследований, проведенных в разных масштабах: от закономерностей распределения организмов вокруг отдельных источников до общей картины биогеографического районирования и вертикального распределения биоценотических комплексов в Мировом океане. При анализе использованы данные визуальных наблюдений из глубоководных обитаемых станций «Пайсис» и «Мир». Абиотические факторы, влияющие на биоценоз, равно как и взаимоотношения между самими видами, в обсуждаемой статье не рассматриваются.

Экосистемы гидротермных источников имеют весьма четкие границы. Среди гидротермального населения доминирующие (руководящие) виды ясно выражены. В то же время, гидротермали сформированы близкими условиями обитания и обладают чертами физиономического сходства. Следовательно, их можно сравнивать, опираясь на доминирующие виды и сообщества, этими видами возглавляемые. Микросообществу с конкретным доминирующим видом, по аналогии с геоботаникой, присвоен ранг ассоциации. Ареалы распространения таких ассоциаций, определяемых видами-доминантами, очерчивают, по сути, не области распространения видов, а области распространения возглавляемых ими сообществ, и мы имеем дело не с фаунистическим, а с ландшафтно-экологическим подходом.
На гидротермальных источниках океана можно встретить различные фаунистические ассоциации – в том числе, по причине сукцессионной смены видов. Но в пределах достаточно крупных областей список видов более или менее постоянен. Такими областями являются «гидротермальные районы» – группы источников, идущих вдоль одного рифта, размерами в десятки километров, отстоящие друг от друга на сотни и тысячи километров. Они и приняты в обсуждаемой работе за «минимальный выдел».
В результате анализа 62 доминирующих видов и 27 гидротермальных районов было отмечено восемь биогеографических регионов. Набор ассоциаций в пределах одного региона назван биоценотическим комплексом.
Приведенная картина биоценотического районирования Мирового океана не противоречит в целом предложенным ранее схемам, полученным в результате сопоставления фаунистических ареалов – как всех видов гидротермали, так и строго специфических ее обитателей. По доминированию тех или иных семейств животных было выделено пять крупных биогеографических комплексов: Восточно-Пацифический, Западно-Пацифический, Южно-Японский, Атлантический, Индо-океанский. При вычленении регионов доминанты рассматривались более подробно – до рода – потому регионов получилось восемь.
Сравнение фауны гидротермалей океана показывает, что она возникла в западной Пацифике, откуда через Индийский океан проникла в южную Атлантику и далее расселилась по всему Атлантическому океану.
Вертикальное распределение обитателей гидротермали представлено тремя основными комплексами ассоциаций: сублиторально-верхнебатиальным (82-1000м), батиальным (1500-3000м) и абиссальным (3000-3600м).
На глубинах 82-110 м, в зонах небольших температурных аномалий, описаны лишь поселения вестиментифер и сопутствующей им фоновой фауны. Большинство классических сообществ гидротермали развиваются начиная с глубин 400м, а специализированные формы доминируют глубже 2000м.
Если роль фотосинтеза с глубиной уменьшается, то значение хемосинтетической органики возрастает, и сама структура гидротермальных экосистем по мере возрастания глубины – усложняется. Несмотря на распространенность бактериального фотосинтеза на относительно небольших глубинах (например, в районах холодных метановых высачиваний – сипах, в толще донного осадка ниже поверхностного окисленного слоя, в местах захоронения органики – древесины или трупов крупных животных), там не сформировалось сложных сообществ, чьим основным источником энергии был бы хемосинтез. Для существования «хемосинтетических» экосистем, по всей видимости, важен не только сам поток энергии от хемосинтеза, но и преобладание его над фотосинтезом. Жизнь рядом с гидротермальным источником не очень полезна для здоровья: ради защиты от тяжелых металлов, извергаемых теми же курильщиками, животным приходится снижать скорость питания (фильтрации) или синтезировать весьма «дорогостоящие» металлотионеиновые белковые комплексы для инактивации металлов. В зоне фотосинтеза хемосинтетические экосистемы не конкурентоспособны. К тому же, на больших глубинах возрастает давление, соответственно, растворимость газов и проницаемость мембран, и, вероятно, изменяется равновесие химических реакций, что может повысить «выгоду» хемосинтеза.
Тем не менее, границы вертикальных гидротермальных биоценологических комплексов совпадают с границами сублиторальной, батиальной и абиссальной зон глобальной вертикальной зональности океана, выделенными по распространению обычной не гидротермальной фауны. В эти крупномасштабные экосистемы гидротермальные сообщества входят как весьма своеобразный, но вполне естественный компонент.
Зона эвфотическая — верхняя (в среднем 200 м) зона океана, где освещенность достаточна для фото-синтетической жизнедеятельности растений. Здесь обильно представлен фитопланктон. Наиболее интенсивно процесс фотосинтеза идет на глубинах 25—30 м, где освещенность составляет не менее 1/3 от освещенности поверхности моря. На глубине более 100 м интенсивность освещения снижается до величины 1/100. В районах Мирового океана, где воды особенно прозрачны, фитопланктон может обитать на глубинах до 150—200 м.[ …]
Глубинные воды Мирового океана отличаются большой однородностью, но вместе с тем все типы этих вод имеют свои характерные черты. Формируются глубинные воды главным образом в высоких широтах в результате смешения поверхностных и промежуточных вод в областях циклонических круговоротов, расположенных вблизи материков. К основным очагам образования глубинных вод относятся северо-западные районы Тихого, Атлантического океанов и районы Антарктиды. Они располагаются между промежуточными и придонными водами. Толщина этих вод в среднем 2000—2500 м. Она максимальна (до 3000 м) в экваториальной зоне и в районе субантарктических котловин.[ …]
Глубина D получила название глубины трения. На горизонте, равном удвоенной глубине трения, направления векторов скорости дрейфового течения на этой глубине и на поверхности океана совпадут. Если глубина водоема в рассматриваемом районе больше глубины трения, то такой водоем следует считать бесконечно глубоким. Таким образом, в приэкваториальной зоне Мирового океана глубины, вне зависимости от их реального значения, следует считать малыми и рассматривать дрейфовые течения как течения в неглубоком море.[ …]
С глубиной плотность изменяется в связи с изменением температуры, солености и давления. При понижении температуры и увеличении солености плотность увеличивается. Однако нормальная стратификация плотности нарушается в отдельных районах Мирового океана в связи с региональными, сезонными и другими изменениями температуры и солености. В экваториальной зоне, где поверхностные воды относительно опреснены и имеют температуру 25—28° С, они подстилаются более солеными холодными водами, поэтому плотность резко возрастает до горизонта 200 м, а затем медленно увеличивается к 1500 м, после чего становится почти постоянной. В умеренных широтах, где в предзимнее время происходит охлаждение поверхностных вод, плотность увеличивается, развиваются конвективные токи и более плотная вода опускается, а менее плотная поднимается к поверхности — возникает вертикальное перемешивание слоев.[ …]
В рифтовых зонах Мирового океана было выявлено около 139 глубинных гидротермальных полей (65 из них активных, см. рис. 5.1) [466, 367]. Можно ожидать, что число таких систем будет увеличиваться по мере дальнейших исследований рифтовых зон. Наличие 17 активных гидротермальных систем вдоль отрезка неовулканической зоны длиной 250 км в рифтовой системе Исландии и по крайней мере 14 активных гидротермальных систем вдоль отрезка длиной 900 км в Красном море указывает пространственный диапазон в распределении гидротермальных полей между 15 и 64 км [429].[ …]
Своеобразной зоной Мирового океана, характеризующейся высокой рыбопродуктивностью, является апвеллинг, т.е. подъем вод из глубины в верхние слои океана, как правило, на западных берегах контингентов.[ …]
Поверхностная зона (с нижней границей на глубине в среднем 200 м) характеризуется высокой динамичностью и изменчивостью свойств вод, обусловленной сезонными колебаниями температуры и ветровым волнением. Объем заключенной в ней воды 68,4 млн. км3, что составляет 5,1% объема воды Мирового океана.[ …]
Промежуточная зона (200—2000 м) отличается сменой поверхностной циркуляции с ее широтным переносом вещества и энергии на глубинную, в которой превалирует меридиональный перенос. В высоких широтах к этой зоне приурочен слой более теплой воды, проникшей из низких широт. Объем воды в промежуточной зоне 414,2 млн. км3, или 31,0% Мирового океана.[ …]
Самая верхняя часть океана, куда проникает свет и где создается первичная продукция, называется эвфотической. Ее мощность в открытом океане доходит до 200 м, а в прибрежной части — не более 30 м. По сравнению с километровыми глубинами это зона достаточно тонкая и отделяется компенсационной зоной от значительно большей водной толщи, вплоть до самого дна — афотической зоны.[ …]
В пределах открытого океана выделяют три зоны, основным различием которых является глубина проникновения солнечных лучей (рис. 6.11).[ …]
Помимо экваториальной зоны апвеллинга, подъем глубинных вод возникает там, где сильный постоянный ветер отгоняет поверхностные слои от берега больших водоемов. Учитывая выводы теории Экмана, можно констатировать, что апвеллииг происходит при касательном к берегу направлении ветра (рис. 7.17). Смена направления ветра на противоположное ведет к смене апвеллинга на даунвеллинг или наоборот. На зоны апвеллинга приходится всего 0,1% площади Мирового океана.[ …]
Глубоководные рифтовые зоны океана находятся на глубине около 3000 м и более. Условия жизни в экосистемах глубоководных рифтовых зон очень своеобразны. Это полная темнота, огромное давление, пониженная температура воды, недостаток пищевых ресурсов, высокая концентрация сероводорода и ядовитых металлов, встречаются выходы горячих подземных вод, и т. д. В результате живущие здесь организмы претерпели следующие адаптации: редукция плавательного пузыря у рыб или заполнение его полости жировой тканью, атрофирование органов зрения, развитие органов светосвечения и др. Живые организмы представлены гигантскими червями (погонофорами), крупными двустворчатыми моллюсками, креветками, крабами и отдельными видами рыб. Продуцентами выступают сероводородные бактерии, живущие в симбиозе с моллюсками.[ …]
Материковый склон — это зона перехода от материков к ложу океана, расположенная в пределах 200—2440 м (2500 м). Она характеризуется резкой сменой глубин и значительными уклонами дна. Средние уклоны дна 4—7°, в отдельных районах доходят до 13—14°, как, например, в Бискайском заливе; известны еще большие уклоны дна возле коралловых и вулканических островов.[ …]
При подъеме по разломной зоне с раздвижением до глубин 10 км и менее (от уровня дна океана), что приблизительно соответствует положению границы Мохоровичича в океанической литосфере, ультра-основная мантийная интрузия может попадать в зону циркуляции термальных вод. Здесь при Т= 3 00- 500°С создаются благоприятные условия для процесса серпентинизации ультрабазитов. Наши расчеты (см. рис. 3.17, а), а также наблюдаемые над такими разломиыми зонами повышенные значения теплового потока (в 2-4 раза превышающие нормальные значения q для океанической коры) предполагают наличие температурного интервала серпентинизации на глубинах 3—10 км (эти глубины сильно зависят от положения кровли высокотемпературного интрузивного мантийного материала). Постепенная серпентинизация перидотитов понижает их плотность до значений, меньших плотности окружающих пород океанической коры, и приводит к увеличению их объема на 15-20% [45].[ …]
В дальнейшем будет видно, что глубина трения в средних широтах и при средних скоростях ветра бывает невелика (примерно порядка 100 м). Следовательно, уравнения (52) можно применять в простой форме (47) во всяком море со сколько-нибудь значительной глубиной. Исключение составляет область мирового океана, лежащая по соседству с экватором, где ¡sin ф стремится к нулю, а глубина трения — к бесконечности. Разумеется, пока здесь речь идет об открытом море; что касается прибрежной зоны, то о ней придется много говорить в дальнейшем.[ …]
Батиаль (от греч. — глубокий)— зона, занимающая промежуточное положение между материковой отмелью и ложем океана (от 200—500 до 3000 м), т. е. соответствует глубинам материкового склона. Эта экологическая область характеризуется быстрым нарастанием глубины и гидростатистическим давлением, постепенным понижением температуры (в низких и средних широтах— 5—15°С, в высоких— от 3° до— 1°С), отсутствием фотосинтезирующих растений и др. Донные осадки представлены органогенными илами (из скелетных остатков фораминифер, кокколитофорид и пр.). В этих водах бурно развиваются автотрофные хемосинтезирующие бактерии; характерны многие виды плеченогих, морские перья, иглокожие, десятиногие ракообразные, из придонных рыб обычны долгохвосты, угольная рыба и др. Биомасса — обычно граммы, иногда десятки граммов/м2.[ …]
От описанных выше сейсмоактивных зон срединно-океанических хребтов существенно отличаются те, которые расположены в районах островных дуг и активных континентальных окраин обрамления Тихого океана. Хорошо известно, что характерная особенность таких зон — их проникновение до очень больших глубин. Глубины очагов землетрясений здесь достигают 600 и более километров. При этом, как показали исследования С. А. Федотова, Л. Р. Сайкса и А. Хасегавы, ширина уходящей в глубь зоны сейсмической активности не превышает 50-60 км. Другая важная отличительная особенность этих сейсмоактивных зон — механизмы в очагах землетрясений, вполне определенно свидетельствующие о сжатии литосферы в районе внешнего края островных дуг и активных континентальных окраин.[ …]
Экосистема глубоководных рифтовых зон океана — эта уникальная экосистема была открыта американскими учеными в 1977 г. в рифтовой зоне подводного хребта Тихого океана. Здесь на глубине 2 600 м, в сплошной темноте, при обильном содержании сероводорода и ядовитых металлов, выделяющихся из гидротермальных источников, были обнаружены «оазисы жизни». Живые организмы были представлены гигантскими (до 1—1,5 м длиной), живущими в трубках червями (погонофорами), крупными белыми двустворчатыми моллюсками, креветками, крабами и отдельными экземплярами своеобразных рыб. Биомасса только погонофор достигала 10—15 кг/м2 (на соседних участках дна — всего 0,1—10 г/м2). На рис. 97 показаны особенности данной экосистемы в сравнении с наземными биоценозами. Серобактерии составляют первое звено пищевой цепи этой уникальной экосистемы, далее идут погонофоры, внутри тела которых обитают бактерии, перерабатывающие сероводород в необходимые питательные вещества. В экосистеме рифтовых зон 75 % биомассы составляют организмы, живущие в симбиозе с хемоавтотрофными бактериями. Хищники представлены крабами, брюхоногими моллюсками, отдельными видами рыб (макруридами). Аналогичные «оазисы жизни» были обнаружены в глубоководных рифтовых зонах многих районов Мирового океана. Подробнее можно ознакомиться в книге французского ученого Л. Лобье «Оазисы на дне океана» (Л., 1990).[ …]
На рис. 30 показаны основные экологические зоны Мирового океана, показывающие вертикальную зональность распределения живых организмов. В океане прежде всего выделяют две экологические области: толщу воды — пелагиалъ и дно — ёенталъ. В зависимости от глубины бенталь делится на литоральную (до 200 м), батиальную (до 2500 м), абиссальную (до 6000 м) и улътраабис-сальную (глубже 6000 м) зоны. Пелагиаль также подразделяется на вертикальнее зоны, соответствующие по глубине зонам бентали: эпипелаги-аль, батипелагиаль и абиссопелагиалъ.[ …]
Крутой материковый склон океана заселен представителями батиальной (до 6000 м), абиссальной и ультраабиссальной фауны; в этих зонах, за пределами доступного для фотосинтеза освещения, растения отсутствуют.[ …]
Абиссаль (от греч. — бездонный) — экологическая зона распределения жизни на дне Мирового океана, соответствующая глубинам океанического ложа (2500—6000 м).[ …]
До сих пор речь шла о воздействии на физические параметр: океана и лишь косвенно предполагалось, что таким образом чере эти параметры идет воздействие и на экосистемы. С одной стс роны, подъем богатых биогенными солями глубинных вод може служить фактором повышения биопродуктивности этих в оС щем-то бедных районов. Можн рассчитывать на то, что подъем глубинных вод позволит снизит температуру поверхностных вод хотя бы в каких-то локальны зонах с одновременным увеличением за счет повышения раствс римости кислорода содержания последнего. С другой стороны, со сбросом в среду холодны вод связана гибель теплолюбивых видов с низкой термическо устойчивостью, изменение видового состава организмов, кормово базы и т. п. Кроме того, экосистема будет постоянно подвергатьс воздействию биоцидов, препятствующих обрастанию рабочих эле ментов станции, воздействию различных реагентов, металлов, шг сел и других побочных выбросов.[ …]
Главным фактором, дифференцирующим морскую биоту, является глубина моря (см. рис. 7.4): материковый шельф резко сменяется материковым склоном, плавно переходящим в материковое подножие, которое опускается ниже к ровному ложу океана — абиссальной равнине. Этим морфологическим частям океана примерно соответствуют следующие зоны: неритическая — шельфу (с литоралью — приливно-отливной зоной), батиальная — материковому склону и его подножию; абиссальная — область океанических глубин от 2000 до 5000 м. Абиссальная область разрезается глубокими впадинами и ущельями, глубина которых более 6000 м. Область открытого океана за пределами шельфа называют океанической. Все население океана, так же как и в пресноводных экосистемах, делится п планктон, нектон, бентос. Планктон и нектон, т.е. все, что живет в открытых водах, образует так называемую пелагическую зону.[ …]
| Распределение перепадов температур в приэкваториальных зонах Мирового океана [1] (пунктиром отмечены глубины 1000 м) | ![Распределение перепадов температур в приэкваториальных зонах Мирового океана [1] (пунктиром отмечены глубины 1000 м)](https://ru-ecology.info/static/pngsmall/822124936.png) |
Принято считать, что береговые станции рентабельны, если требуемые глубины с подходящей температурой охлаждающей воды находятся достаточно близко от побережья и длина трубопровода не превышает 1—3 км. Такая ситуация характерна для многих островов тропического пояса, представляющих собой вершины подводных гор и потухших вулканов и не имеющих свойственного материкам протяженного шельфа: их берега достаточно круто спускаются по направлению к океанскому ложу. Если берег достаточно удален от зон требуемых глубин (например, на островах, окруженных коралловыми рифами) или отделен полого понижающимся шельфом, то для сокращения длины трубопроводов энергоблоки станций могут быть вынесены на искусственные острова или стационарные платформы — аналоги используемых при морской добыче нефти и газа. Преимущества наземных и даже островных станций в том, что исчезает необходимость в создании и обслуживании дорогостоящих, подвергающихся воздействию открытого океана сооружений — будь то искусственные острова или стационарные основания. Однако два существенных фактора, ограничивающих береговое базирование, все-таки остаются: органиченность соответствующих островных территорий и необходимость прокладки и защиты трубопроводов.[ …]
Впервые морфологическая характеристика и типизация океанических разломных зон по морфологическим признакам (на примере разломов северо-восточной части Тихого океана) была сделана Г.Менардом и Т.Чейсом [401]. Они определили разломы как ’’длинные и узкие зоны сильно расчлененного рельефа, характеризующиеся присутствием вулканов, линейных хребтов, уступов и обычно отделяющие друг от друга различные топографические провинции с неодинаковыми региональными глубинами”. Выраженность трансформных разломов в рельефе дна океана и аномальных геофизических полях, как правило, довольно резкая и четкая. Это подтвердили и многочисленные детальные исследования, проведенные в последние годы [66, 78, 104, 176, 335]. Высокие приразломные хребты и глубокие впадины, сбросы и трещины характерны для зон трансформных разломов. Аномалии А , АТ, теплового потока и другие свидетельствуют о гетерогенности строения литосферы и сложной динамике разломных зон [335, 142, 47]. Кроме того, разновозрастные блоки литосферы, расположенные по разные стороны от разлома, в соответствии с законом V/ имеют различное строение, выражающееся в различных глубине дна и толщине литосферы, что создает дополнительные региональные аномалии в геофизических полях [38,47].[ …]
Область континентального шельфа, неритическая область, если ее площадь ограничить глубиной до 200 м, составляет около восьми процентов площади океана (29 млн км2) и является самой богатой в фау-нистическом отношении в океане. Прибрежная зона благоприятна по условиям питания, даже в дождевых тропических лесах нет такого разнообразия жизни, как здесь. Очень богат кормом планктон за счет личинок бентосной фауны. Личинки, которые остаются несъеденны-ми, оседают на субстрат и образуют либо эпифауну (прикрепленную), либо инфауну (закапывающуюся).[ …]
У планктона также выражена вертикальная дифференциация при адаптации разных видов к разным глубинам и разной интенсивности освещения. Вертикальные миграции влияют на распределение этих видов, и потому ярусность по вертикали в этом сообществе менее очевидна, чем в лесу. Сообщества освещенных зон на дне океана ниже уровня прилива дифференцированы отчасти интенсивностью света. Виды зеленых водорослей концентрируются на мелководье, виды бурых водорослей распространены на несколько больших глубинах, а еще ниже особенно обильны красные водоросли. Бурые и красные водоросли содержат, помимо хлорофилла и каротиноидов, дополнительные пигменты, что позволяет им использовать свет низкой интенсивности и отличный по спектральному составу от света в мелководьях. Вертикальная дифференциация, таким образом, является общей чертой природных сообществ.[ …]
Абиссальные ландшафты — царство мрака, холодных, малоподвижных вод и очень бедной органической жизни. В олиштроф-ных зонах Океана биомасса бентоса колеблется от 0,05 и менее до 0,1 г/м2, несколько повышаясь в областях богатого поверхностного планктона. Но и здесь, на таких больших глубинах, встречены «оазисы жизни». Почвы» абиссальных ландшафтов образованы илами. Состав их, как и наземных почв, зависит от широты места и высоты (в данном случае глубины). Где-то на глубине 4000—5000 м преобладавшие ранее карбонатные илы сменяются бескарбонатными (красными глинами, радио-ляриевым илом в тропиках и диатомовым в умеренных широтах).[ …]
Здесь х коэффициент термической диффузии пород литосферы, Ф — функция вероятности, (Т + Сг) — температуры мантии под осевой зоной срединного хребта, т.е. при / = 0. В модели пограничного слоя глубина изотерм и подошвы литосферы, а также глубина дна океана И, отсчитываемая от ее значения на оси хребта, увеличиваются пропорционально значению V/.[ …]
В высоких широтах (выше 50°) происходит разрушение сезонного термоклина с конвективным перемешиванием водных масс. В приполярных областях океана имеет место восходящее движение глубинных масс. Поэтому эти широты океана относятся к высокопродуктивным районам. По мере дальнейшего продвижения к полюсам продуктивность начинает падать из-за понижения температуры воды и уменьшения ее освещенности. Для океана характерна не только пространственная изменчивость продуктивности, но и повсеместная сезонная изменчивость. Сезонная изменчивость продуктивности обусловлена в значительной мере реакцией фитопланктона на сезонные изменения условий среды обитания, прежде всего освещенности и температуры. Наибольшая сезонная контрастность наблюдается в умеренной зоне океана.[ …]
Поступление магмы в магматическую камеру происходит, видимо, эпизодически, и является функцией высвобождения большого количества расплавленного вещества с глубин более 30 — 40 км в верхней мантии. Концентрация расплавленного вещества в центральной части сегмента приводит к увеличению объема (разбуханию) магматической камеры и миграции расплава вдоль оси к краям сегмента. С приближением к трансформному разлому глубина кровли, как правило, опускается вплоть до полного исчезновения соответствующего горизонта близ трансформного разлома. Это в значительной степени обусловлено охлаждающим влиянием более старого литосферного блока, граничащего с осевой зоной по трансформному разлому (эффект трансформного разлома). Соответственно, наблюдается и постепенное погружение уровня дна океана (см. рис.3.2).[ …]
В антарктическом районе южного полушария дно океанов покрыто ледниковыми и айсберговыми отложениями и диатомовыми илами, встречающимися также на севере Тихого океана. Дно Индийского океана выстлано илом с большим содержанием углекислого кальция; глубоководные впадины — красной глиной. Наиболее разнообразны отложения дна Тихого океана, где на севере господствуют диатомовые илы, северная половина покрыта в области глубин свыше 4000 м красной глиной; в приэкваториальной зоне восточной части океана распространены илы с кремнистым остатком (радиоляриевые), в южной половине на глубинах до 4000 м встречаются известково-карбонатные илы,. красная глина, на юге — диатомовые и ледниковые отложения. В районах вулканических островов и коралловых рифов встречается вулканический и коралловый песок и ил (рис. 7).[ …]
Смена континентальной земной коры на океаническую происходит не постепенно, а скачкообразно, сопровождаясь образованием морфоструктур особого рода, свойственных переходным, точнее— контактным, зонам. Иногда их называют периферическими областями океанов. Главнейшими морфоструктурами их являются островные дуги с действующими вулканами, резко переходящие в сторону океана в глубоководные желоба. Именно здесь, в узких, глубочайших (до 11 км) впадинах Мирового океана, проходит структурная граница континентальной и океанической коры, совпадающая с глубинными разломами, известными у геологов под названием зоны Заварицкого — Бенъофа. Разломы, падающие под материк, идут на глубину до 700 км.[ …]
Второй специальный эксперимент по изучению синоптической изменчивости океанских течений («Полигон—70») был проведен советскими океанологами во главе с Институтом океанологии АН СССР в феврале—сентябре 1970 г. в северной пассатной зоне Атлантики, где в течение шести месяцев были осуществлены непрерывные измерения течений на 10 глубинах от 25 до 1500 м на 17 заякоренных буйковых станциях, образовывавших крест размерами 200X200 км с центром в точке 16°ЗГ 14, 33°30 Ш, и был также выполнен ряд гидрологических съемок.[ …]
Так была внесена поправка в представление о не-возобновимости минеральных богатств. Полезные ископаемые за исключением торфа и некоторых других естественных образований невозобновимы в отработанных месторождениях на досягаемой для человека глубине недр континентов. Это и понятно — невозвратно исчезли те физико-химические и иные условия в зоне месторождения, которые в далеком прошлом геологической истории создали ценные для человека минеральные образования. Другое дело добыча со дна существующего океана гранулированных руд. Мы можем их взять, а в создавшей эти руды естественной действующей лаборатории, какой является океан, не прекратятся процессы рудообразования.[ …]
Если гравитационные аномалии в свободном воздухе на континентах и океанах не имеют принципиальных различий, то в редукции Буге эта разница проявляется весьма заметно. Введение поправки за влияние промежуточного слоя в океане приводит к получению высоких положительных значений аномалий Буге, тем больших, чем больше глубина океана. Данный факт обусловлен теоретическим нарушением при введении поправки Буге («засыпке» океана) природной изостазии океанической литосферы. Так, в гребневых зонах СОХ аномалия Буге составляет около 200мГал, для абиссальных океанических котловин — в среднем от 200 до 350 мГал. Несомненно, что в аномалиях Буге отражаются генеральные черты рельефа дна океана в той степени, в какой они изостатически скомпенсированы, поскольку основной вклад в аномалии Буге вносит именно теоретическая поправка.[ …]
Основными процессами, определяющими профиль окраины, возникшей у тылового края континента (пассивной окраины), являются почти перманентные погружения, особенно значительные в дистальной, приокеанической ее половине. Лишь частично они компенсируются накоплением осадков. Во времени окраина разрастается как вследствие вовлечения в прогибание все более удаленных от океана континентальных блоков, так и в результате формирования мощной осадочной линзы на материковом подножии. Разрастание происходит главным образом за счет соседних участков океанского ложа и является следствием непрекращаю-щейся эрозии прилегающих к окраине районов континента, а также глубинных его областей. Это находит отражение не только в ненеиленизации суши, но и в смягчении, выравнивании рельефа в подводных участках зоны перехода. Происходит своего рода агградация: выравнивание поверхности переходных зон в областях с пассивным тектоническим режимом. Вообще говоря, эта тенденция характерна для любой окраины, однако в тектонически активных зонах она не реализуется вследствие орогенеза, складкообразования, роста вулканических построек.[ …]
В соответствии с особенностями морской воды температура ее даже на поверхности лишена резких контрастов, свойственных приземным слоям воздуха, и колеблется в пределах от —2°С (температура замерзания) до 29°С в открытом Океане (до 35,6°С в Персидском заливе). Но это справедливо в отношении температуры воды на поверхности, обусловленной поступлением солнечной радиации. В рифтовых зонах Океана на больших глубинах открыты мощные гидротермы с температурой воды, находящейся под большим давлением, до 250—300°С. И это не эпизодические излияния перегретых глубинных вод, а длительно (даже по геологическим масштабам) или же постоянно существующие на дне Океана озерки сверхгорячей воды, о чем говорит их экологически неповторимая бактериальная фауна, использующая для своего питания соединения серы. В этом случае амплитуда абсолютного максимума и минимума температуры воды Океана составит 300°С, что в два раза превышает амплитуду предельно высоких и низких температур воздуха у земной поверхности.[ …]
Рассеивание вещества биострома распространяется на значительную часть толщи географической оболочки, а в атмосфере даже выходит за ее пределы. Жизнеспособные организмы обнаружены на высоте более 80 км. В атмосфере нет автономной жизни, но воздушная тропосфера — транспортер, переносчик на огромное расстояние семян и спор растений, микроорганизмов, среда, в которой проводят значительную часть жизни многие насекомые и птицы. Рассеивание водно-поверхностного биострома распространяется на всю толщу океанических вод вплоть до донной пленки жизни. Дело в том, что глубже эвфотической зоны сообщества практически лишены собственных продуцентов, энергетически они полностью зависимы от сообществ верхней зоны фотосинтеза и на этом основании не могут рассматриваться полноценными биоценозами в понимании Ю. Одума (М. Е. Виноградов, 1977). С нарастанием глубины биомасса и численность планктона быстро уменьшаются. В батипелагиали в самых продуктивных районах океана биомасса не превышает 20—30 мг/м3 — это в сотни раз меньше, чем в соответствующих районах на поверхности океана. Глубже 3000 м, в абиссопелагиали, биомасса и численность планктона исключительно низкие.[ …]
В океане выделяют
несколько областей с различными
экологическими условиями: толщу воды
— пелагиаль,
среду обитания
пелагических животных, и дно океана
бенталь,
заселенную
бентосом, т.е. организмами, обитающими
на грунте или в толще грунта. Бенталь
подразделяют на супралитораль,
литораль, сублитораль, батиаль и
абиссаль (рис.
77).
Супралитораль —
побережье океана, расположенное выше
уровня воды в
самый высокий прилив, но еще подверженное
эпизодическому воздействию океанических
вод при нагонах и штормах.
Здесь обитают как наземные, так и морские
организмы.
Литораль —
прибрежная зона морского дна, осушающаяся
во время отлива.
Располагается между уровнями воды в
самый низкий
отлив и в самый высокий прилив, простираясь
до глубины 40—50
м. Обитатели литорали дважды в сутки
покрываются водой и
освобождаются от нее; они живут при
условии резких изменений
температуры и солености воды, действия
прибоя, при прямом
солнечном свете. Некоторые животные,
прикрепляющиеся к
скалам, могут поселяться так высоко,
что покрываются водой лишь
два раза в месяц, при максимальном
сизигийном приливе. Значительное
число видов вне этой полосы существовать
не могут. В
приполярных странах в полосе литорали
жизнь крайне обеднена,
так как значительную часть года она
покрыта льдами.
Сублитораль —
прибрежная
часть океана, непосредственно примыкающая
к литорали и располагающаяся на глубинах
до 200
м, реже глубже, простирающаяся от уровня
воды при самом низком
отливе до нижней границы распространения
донной растительности.
С сублиторалью связаны богатые промыслы
беспозвоночных.
Выделяются мелководья
с мягким и твердым грунтом. На мягком
грунте особенно обильны свойственные
только мелководьям водоросли
и цветковые растения. Из водорослей на
более мелких участках преобладают
зеленые и бурые, в том числе ламинарии
и фукусы, а на
больших глубинах — красные водоросли
(багрянки).
На мягком грунте мелководий обитают
черви, крабы, офиу-ры, морские звезды,
многие моллюски. Некоторые обитатели
способны
зарываться на глубину до 50 см, предпочитая
песчаный
грунт илистому; поскольку последний
из-за обилия гниющих
органических остатков характеризуется
недостатком или отсутствием
кислорода, он заселен слабее песчаного.
Мелководья со
скалистым грунтом отличаются твердостью
субстрата, что препятствует проникновению
в него многих животных. Здесь
встречаются сверлящие организмы, активно
внедряющиеся
в скалы и камни. Обитающие на поверхности
субстрата
усоногие раки и моллюски имеют крепкие
раковины и прочно прикрепляются
к грунту, что не позволяет волнам оторвать
их. В пещерах и
трещинах, характерных для скальных
грунтов, поселяются головоногие
моллюски, а также некоторые другие
беспозвоночные и рыбы (например,
мурены).
Для сублиторали
характерна прерывистость распространения
вдоль континентов;
она прерывается предустьевыми
пространствами
с их сильно опресненной, а иногда даже
пресной водой.
Хотя сублитораль,
если ограничить ее площадь материковой
отмелью до
глубины 200 м, составляет всего 8% площади
океана (29 млн км2),
она значительно богаче в фаунистическом
и флористическом
отношении по сравнению с остальными
областями океана.
Батиаль располагается
на материковых склонах на глубинах
200—2000 м между
неритовой зоной и. абиссалью. Эта область
характеризуется
слабой освещенностью в верхних частях
и полным отсутствием видимых лучей
спектра в нижних. Для нее характерны
незначительные колебания температуры
и солености. Здесь отлагаются
преимущественно илы.
Абиссаль —
область наибольших морских глубин,
превышающих
2000 м, соответствующая ложу Мирового
океана. Характеризуется
чрезвычайно медленным передвижением
воды от полюсов к
экватору, постоянно низкими температурами
(~ 0°), практически
почти полным отсутствием света. Здесь
не встречаются зеленые
растения, накапливаются преимущественно
органогенные (в
том числе радиоляриевые и глобигериновые)
илы.
Толща воды —
пелагиаль —
подразделяется наэпипелагиаль
-освещенные
слои воды до глубины 200 м, — и глубоководную
пелагиаль.
Для пелагиали
характерны представители четырех групп
организмов:
нектона,
планктона, плейстона и нейстона. Нектон
— совокупность
активно плавающих организмов, основные
представители
этой группы — пелагические рыбы. Виды
планктона —
организмы,
переносимые морскими течениями и
волнами, так как
сами они могут передвигаться весьма
медленно. Плейстон
и нейстон
— это небольшое
число видов растений и животных, обитающих
на поверхности воды.
Основу всей жизни
пелагиали составляет фитопланктон.
Малая скорость движения планктона
ни в коей мере не может противостоять
движению воды. У видов планктона (как и
у представителей
нектона) имеются приспособления,
препятствующие погружению
в толщу воды. Из них можно назвать
следующие: уменьшение
относительной плотности за счет выработки
веществ, имеющих
малую плотность (в первую очередь жиров),
а также насыщения
тканей и полостей тела водой (так, у
некоторых медуз
вода составляет 99% от массы тела).
Некоторые виды имеют специальные
воздушные камеры, уменьшающие их
плотность.
К фитопланктону
относятся почти исключительно
микроскопические
одноклеточные организмы, имеющие
хлорофилл и способные
к ассимиляции. Они и являются источником
первичной продукции. Значительно меньшую
роль в этом отношении играют
зеленые растения, прикрепленные ко дну,
— различные водоросли
и цветковые растения, а также хемотрофы.
Биомасса фитопланктона составляет в
Мировом океане 1,5
млрд т, а его продукция — 559 млрдт,
биомасса и продукция фитобентоса
соответственно 0,2 и 0,2 млрд т. В этом
состоит фундаментальное отличие морских
экосистем от наземных, где продукция
составляет лишь небольшую часть общей
биомассы. В целом
водоросли в Мировом океане (включая
фитопланктон) создают
органическое вещество (сухая масса) в
количестве примерно
74% от мировой продукции. «Урожайность»
водорослей составляет
1,3-2,0 т сухого вещества на 1 га поверхности
водной толщи в год (рис. 78).
Зоопланктон
представлен в океане микроскопическими
и более крупными организмами: медузами
и сифонофорами из кишечнополостных,
сальпами и аппендикуляриями из хордовых.
Неосвещенная область
океана, охватывающая батиаль и абиссаль,
обладает некоторыми общими особенностями,
связанными с пессимальными условиями
существования.
Абиссаль занимает
~ 8% морского дна и основную толщу морской
воды, значительно превосходя по площади
шельфовую область, а
по объему — освещенную пелагиаль. При
этом, несомненно, абиссальный
бентос является производным от бентоса
освещенной
области океана, а население абиссальной
пелагиали произошло от обитателей
освещенной пелагиали.
Наряду с отсутствием
света на отборе глубоководных форм
сказываются
высокое давление и низкие температуры,
характерные
для глубин и создающие здесь пессимальные
условия существования.
Об уменьшении числа видов с глубиной
свидетельствуют
хотя бы данные океанологической
экспедиции на корабле «Челленджер».
Ею были обнаружены на разных глубинах
следующие
количества видов крабов: до 36 м — 190;
36—180 м — 75; 180-360 м — 28; 360-900 м — 21; 900-1800
м — 3; 1800— 3600 м — 2.
Та же закономерность прослежена и для
числа видов простейших — фораминифер,
которое равно на глубинах до 100
м — 138, а на глубинах 5000-6000 м — всего 19.
.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #